
38年生马尾松种源生长及材性联合分析
2022-11-29 13:51胡兴峰孙晓波陈厚平殷安政季孔庶
南京林业大学学报(自然科学版) 2022年3期
胡兴峰,吴 帆,孙晓波,陈厚平,殷安政,季孔庶*
(1.南京林业大学林木遗传与生物技术省部共建教育部重点实验室,南方现代林业协同创新中心,江苏 南京 210037;2.安徽省六安市裕安区国有林木良种场,安徽 六安 237010)
马尾松(Pinusmassoniana)是我国南方主要针叶树种之一。马尾松木材管胞较长、柔韧性强、管胞间易交错、结合强度大,近30年来,被确定为我国重要的纸浆用材树种进行定向培育,在我国造纸原料林建设中具有特殊地位[1]。相关研究表明,马尾松管胞形态、木材基本密度和微纤丝角等直接决定其木材物理性质,影响木材加工利用,尤其是在制浆造纸方面,木材管胞形态特征与制浆造纸质量密切相关,影响生产纸张的匀度、抗张强度、耐折度、耐破度、撕裂度、形稳性和不透明度等指标[2-6]。马尾松地理分布范围广,我国相继开展了产区区划[7]、立地分类与评价[8]、建筑材林培育[9]、种源试验[10-11]、种子园建立[12-13]以及分子水平上的相关研究[14-15],并取得了丰硕的研究成果。马尾松木材微纤丝角[5, 16]、木材密度[17]、管胞长和宽度[18]等材质性状在种源间、种源内的家系间、不同林分、单株间、同一个体不同组织间差异显著或极显著。马尾松树高、地径和冠幅等生长性状在地理上存在明显的纬向变异规律,呈现出从南向北逐渐减小的变异趋势,东西向的变异规律不显著[19]。径向水平上微纤丝角由髓心至树皮随年轮增加而明显减小,之后趋于稳定[16]。相关分析表明,有些研究结果支撑生长性状与基本密度呈显著正相关[16],这种情况下,可以通过生长性状的遗传改良使木材基本密度获得遗传增益。但是在多数研究结果中,马尾松的生长性状与基本密度之间呈不相关或者负相关[20],说明通过高生长量单株的选择会对基本密度产生负效应,同时也给通过选择速生且材质优良的马尾松优株来进行遗传改良带来了挑战。
生长与材性联合分析研究是林木遗传改良的重要研究手段之一,可筛选出综合性状优良的种源、家系或单株[21-23]。众所周知,性状表达伴随着林木发育进程而发生变化,而目前涉及的马尾松生长与材性的相关性研究中大多都是以幼龄林或中龄林马尾松为试验材料[24],先前由于试验材料的限制未见成熟材的研究结果,致使选择育种结论与生产实际无法准确关联。因此,本研究以55个地理种源组成的38年生马尾松试验林为对象,通过生长和材性测定,了解马尾松生长和材性的地理、木材径向的变异规律,研究各性状之间的相关性,旨在结合生长和材性,筛选出综合性状优良的种源,为纸浆材原料林营建提供良种,同时为相关研究提供借鉴。
1 材料与方法
1.1 试验地及马尾松种源概况
试验林位于安徽省六安市裕安区林木良种基地(116°12′E,31°40′N),北亚热带温暖湿润的季风气候,四季分明,年降雨量1 239.8 mm,无霜期近225 d,年均气温15 ℃,海拔80~110 m,坡度5°~15°,土壤大多为下蜀系黄土母质上发育而成的黏盘黄棕壤,土层厚70~150 cm,pH 5.5~6.5。
1981年春营造马尾松地理种源试验林,造林密度为10 000株/hm2,共64个种源(表1),采用完全随机区组试验设计,9株小区(3行,每行3株),6次重复。该试验林38年生,期间未间伐,保存率为36.52%,因后期树木个体间生长竞争及病虫害等原因致9个种源存活株数不能满足统计学分析要求,故本研究仅以满足统计学分析要求的55个种源为试验材料。

表1 供试马尾松种源及编号
1.2 研究方法
1.2.1 材料获取
2019年2月,在对马尾松胸径(diameter at breast height, DBH)和树高(tree height,H)进行每木检尺的基础上,将55个种源马尾松按每小区每种源挑选3株生长健壮且无病虫害的优良单株(最大株),采用生长锥(锥径φ=5.5 mm)在树木胸高处由南向北方向钻取一根通过髓心的完整木芯。
1.2.2 基本密度测定
将木芯自髓心向外,每5个年轮切一段(1~5、6~10、11~15、16~20、21~25、26~30、31~35、35~38取样,编号依次为Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ、Ⅶ、Ⅷ)(图1),每年轮段的木材基本密度(basic density, BD,公式中记为dB)采用Smith[25]提出的饱和含水量法(maximum moisture content method)测定。
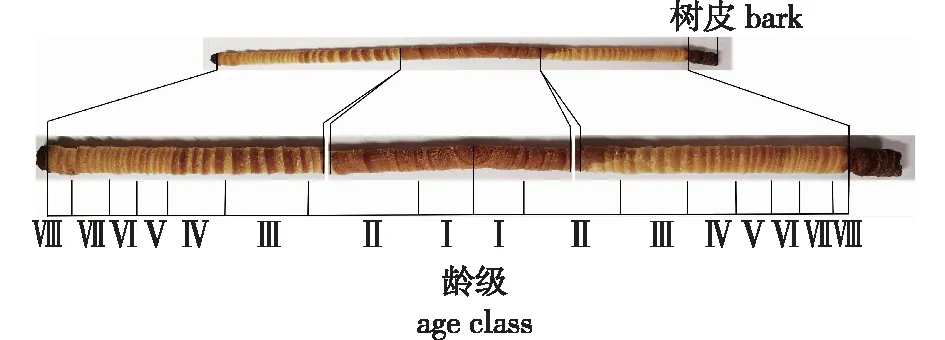
图1 马尾松完整木芯图
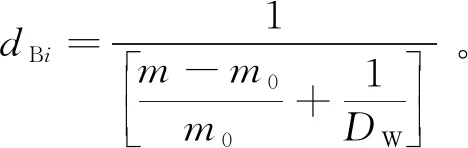
式中:dBi为i年轮段木芯的基本密度;m为i年轮段木芯充分吸水后的质量,g;m0为i年轮段木芯烘干后的质量,g;DW为木材物质比重(取1.53 g/cm3)。
1.2.3 管胞形态测定
将髓心至树皮第1、6、11、16、21、26、31、36年轮木芯切成0.05 cm小段,采用富兰克林离析法解离木材,制片,用3%(质量分数)番红水溶液染色1 min,将制好的样片放置在数码显微图像电脑分析系统上对木材管胞形态进行测定,用软件DigiLabⅡ-C拍照取图,Motic Images Plus 2.0测量管胞长度(tracheid length, TL)和宽度(tracheid width, TW)。管胞长度在1倍物镜下测定,每个样品30次重复;管胞宽度在40倍物镜下测定,每个样品50次重复。
1.2.4 微纤丝角测定
将风干木芯从髓心至韧皮部(去掉树皮)依次取第1、6、11、16、21、26、31、36年轮的早材部分切取厚度约为0.1 cm的试样,采用X-射线衍射法测定,测定仪器及参数为:Utima Ⅳ组合型多功能水平X-射线衍射仪,X光管为铜靶,管电压40 kV,管电流为30 mA,测试范围90°~270°,扫描速度为72°/min,扫描步距为0.36°。扫描得到试样的衍射强度曲线图,利用Origin软件拟合高斯函数,采用0.6 T法计算出微纤丝角(microfibril angle, MFA)。
1.3 数据处理
性状均值、变异系数、相关分析、优良种源评价和综合选择采用Excel 2010和IBM SPSS 20.0软件进行,方差分析利用SAS/STAT软件中的GLM程序。
1)马尾松单株立木材积按1978年国家林业部颁布的标准材积计算公式:
V马尾松=0.000 062 341 803×D1.855 149 7×H0.956 824 92。
式中:V马尾松为各种源平均材积,m3;D为各种源平均胸径,cm;H为各种源平均树高,m。
2)单株材性值据木芯各龄级对应性状与龄级年轮宽度的加权平均数计算而得。
3)方差分析采用的线性随机模型(yijk):
yijk=μ+αi+βj+εijk。
式中:μ为总体均值;αi为种源效应;βi为区组主效应;εijk为随机误差。
4)遗传参数估计。计算3种遗传参数为:
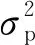
5)利用Origin 2017软件进行性状地理变异趋势作图。性状与产地经纬度间的相关系数计算公式为:

6)性状间相关系数计算公式:

7)表型协方差和遗传协方差模型如下:


2 结果与分析
2.1 马尾松各性状种源变异分析
对38年生的55个地理种源马尾松树高、胸径、材积、管胞长度、管胞宽度、微纤丝角和基本密度7个性状进行统计分析,其平均值、标准差和表型变异系数见表2。由表2可见,马尾松各性状不同种源平均变异系数为3.02%~58.61%,其中材性的变异系数最小,包括管胞长、宽度和木材基本密度等表型变异系数均小于10%。
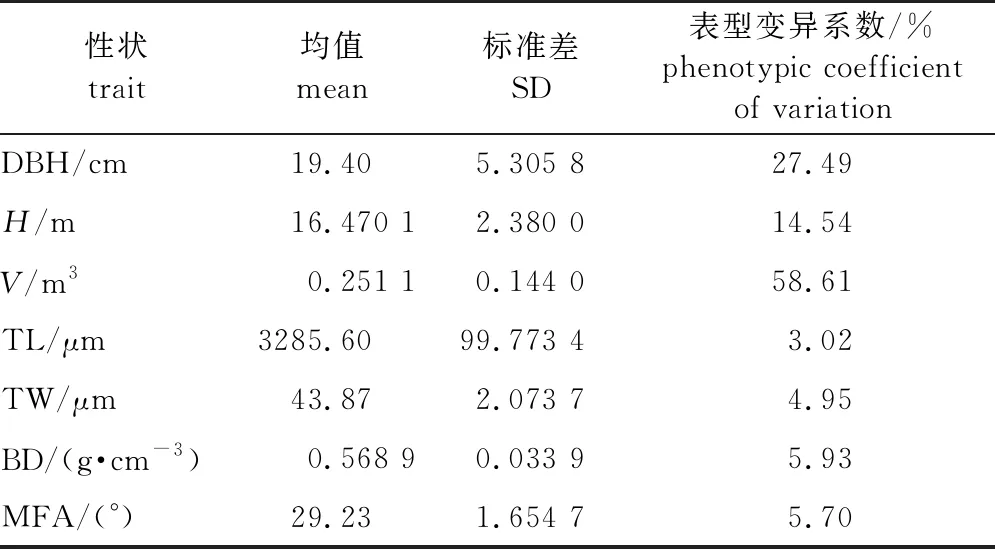
表2 马尾松各性状均值、标准差和表型变异系数
方差分析(表3)表明,马尾松不同种源的胸径、树高、材积、管胞长度、管胞宽度、微纤丝角和基本密度均差异极显著(P<0.01),除管胞宽度和基本密度外,其他性状在区组水平间差异极显著(P<0.01),说明马尾松种源管胞宽度和基本密度相对稳定,在区组间的差异较小。
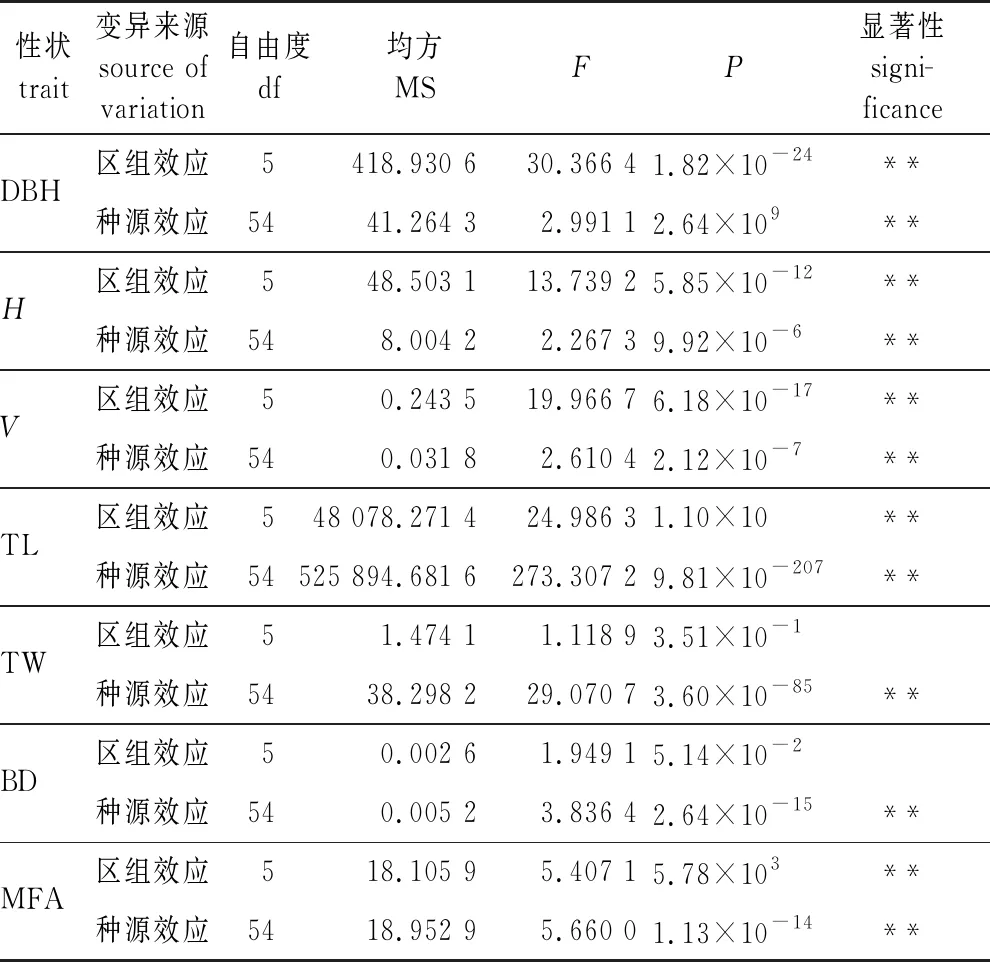
表3 马尾松各性状种源间方差分析
2.2 马尾松各性状遗传参数估计
由马尾松生长性状与材性的遗传参数估计(表4)可知,马尾松各性状表型变异系数和遗传变异系数的分布范围分别为3.02%~58.61%和5.56%~28.16%。胸径、树高、材积、管胞长度、管胞宽度、基本密度和微纤丝角均受中度以上遗传控制,其种源遗传力分别为0.665 7、0.558 9、0.616 9、0.996 3、0.965 6、0.739 3和0.823 3,且材性种源遗传力高于生长性状的种源遗传力。
2.3 马尾松种源性状随产地经纬度的变异趋势
由马尾松各性状与经纬度的相关分析(表5)可知,除木材基本密度与经度呈不显著正相关外,树高、胸径、材积、管胞长度、管胞宽度和微纤丝角与经度均呈不显著负相关。基本密度和微纤丝角与纬度呈不显著正相关,其他性状与之呈负相关,并且树高、胸径和材积与纬度呈极显著负相关,相关系数分别为:-0.427、-0.561和-0.539。说明马尾松的生长性状从南向北逐渐减小,其速生种源分布在南部种源区。由于马尾松不同种源的水热条件、海拔等不同,各性状在地理上表现出一定的变异趋势(图2),胸径、树高和材积总体上从东南向西北逐渐减小,树高在东经111° ~114°处较高,管胞长度、管胞宽度、基本密度和微纤丝角从南向北先增大后减小,其中管胞长度、管胞宽度和基本密度均在北纬27°~30°处最大。

表5 马尾松各性状与经纬度间的相关性分析
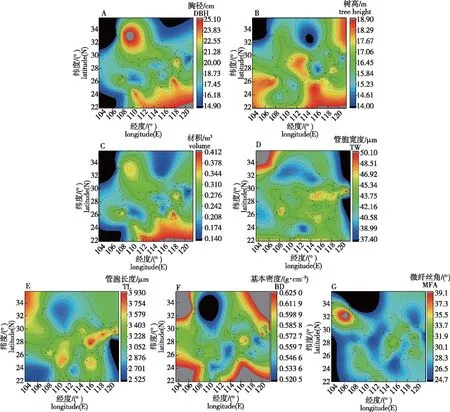
图2 马尾松各性状地理变异趋势图
2.4 不同种源马尾松材性径向变化规律
马尾松各材性在径向不同龄级间差异极显著(P<0.001)(表6)。马尾松各材性径向不同龄级间存在一定的变化规律(图3),微纤丝角(MFA)从髓心至树皮总体随着龄级增大而减小,变化范围为20.8°~35.4°,并且第Ⅳ龄级后各龄级间变化小于第Ⅳ龄级前各龄级间的变化;管胞宽度(TW)从髓心至树皮总体随着龄级的逐渐增大而增大,变化范围为40.7~47.6 μm,从第Ⅳ龄级开始变化幅度逐渐减小,至第Ⅶ龄级后有所上升;管胞长度(TL)从髓心至树皮总体随着龄级的逐渐增大而增大,变化范围为2 365.1~4 481.7μm,变化幅度随着龄级的增大而减小;基本密度(BD)从髓心至树皮总体随着龄级的逐渐增大而增大,变化范围为0.499 9~0.747 9 g/cm3。

表6 马尾松各材性龄级间方差分析
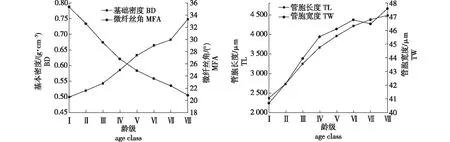
图3 马尾松各性状不同龄级遗传变异分析图
2.5 不同种源马尾松各性状相关性分析
马尾松生长与材性相关分析(表7)可知,树高与胸径,材积与树高、胸径的表型相关系数分别为0.635、0.719和0.963(P<0.01)。木材基本密度与胸径、材积的表型相关系数分别为-0.231、-0.197(P<0.01),说明生长性状与材性间呈显著负相关,且生长性状间的相关性高于生长与材性之间的相关性。管胞长与宽度间呈显著正相关(P<0.01),胸径、树高与管胞长度,树高与木材基本密度,以及材积与微纤丝角均呈不显著负相关。胸径、树高与管胞宽度,微纤丝角与胸径、树高、管胞宽度和基本密度间呈不显著正相关。

表7 马尾松各性状间的相关性分析
通过遗传相关性分析发现,管胞长度与管胞宽度,胸径与树高,材积与胸径、树高的遗传相关系数分别为0.524、0.632、0.735和0.967(P<0.01)。胸径与微纤丝角,基本密度与树高、胸径、材积的遗传相关系数分别为-0.333、-0.581、-0.417和-0.516(P<0.01),说明生长性状与材性间呈极显著负相关,且生长性状间相关性高于生长与材性。管胞长度、宽度与胸径、树高和材积,基本密度与管胞长度、微纤丝角遗传上呈不显著正相关;基本密度与管胞宽度,微纤丝角与树高、材积、管胞宽度和管胞长度遗传上均呈不显著负相关。
2.6 马尾松多性状综合指数选择
在马尾松的改良中,期望得到胸径、树高和材积大,管胞长度长,管胞宽度小,基本密度大,微纤丝角小的优良纸浆材种源,为此利用等权法获得胸径(式中记为DDBH)、树高(H)、材积(V)、管胞长度(lT)、管胞宽度(WT)、基本密度(dB)和微纤丝角(AMFA)7个性状的经济权重分别为:0.171 6、0.378 6、6.201 7、0.003 2、-0.298 3、22.360 7、-0.339 7。由以上7个性状构建综合指数选择方程为:I=0.114 2DDBH0.211 6H+ 3.825 9V+0.003 2lT-0.288 1WT+16.531 3dB-0.279 7AMFA。将各种源各性状均值代入选择指数方程,对马尾松所有种源生长性状和材性进行综合评价,获得平均选择指数I=6.041,其标准差(σ)=0.855,再以种源选择指数值大于I+2σ=7.751为优良种源,共选出了5个符合改良条件的优良种源(表8),入选率为9.09%,其胸径、树高、材积、管胞长度获得平均遗传增益分别为:3.69%、5.72%、15.12%、7.05%。

表8 马尾松5个优良种源各性状均值及选择值指数
3 讨 论
3.1 马尾松不同种源遗传变异
笔者利用已达成熟期的马尾松种源试验林,发现38年生的马尾松胸径、树高、材积、管胞长和宽、基本密度和早材微纤丝角均在种源间差异极显著(P<0.01),进一步验证了马尾松在种源水平上各性状差异显著的结论[12-13],这种差异是由不同种源马尾松长期适应不同立地条件所致,而马尾松的天然群体分布广泛,致使马尾松生长和材性形成种源上的差异。
本研究表明马尾松树高、胸径、材积、管胞长度、管胞宽度和微纤丝角与经度均呈不显著负相关。基本密度和微纤丝角与纬度呈不显著正相关,其他性状与之呈负相关,并且树高、胸径和材积与纬度的负相关极显著,在青钱柳中也有类似的地理变异规律,但是在局部也有不同的变异趋势[27]。本研究表明胸径、树高和材积总体上从东南向西北逐渐减小,树高在东经111°~114°处较高,管胞长度、管胞宽度、基本密度和早材微纤丝角从南向北先增大后减小,其中管胞长度、管胞宽度和基本密度均在北纬27°~30°处最大。这可能是由于不同种源马尾松生长和材性还受到地形地貌、水热条件、人为活动等多种因素的影响,有待进一步研究。
3.2 马尾松不同龄级材性变化规律
木材径向变化是指由于树干径向上在木材构造、木材组成与木材性质方面均有差异的现象。本研究结果表明,马尾松基本密度、管胞长度、宽度及微纤丝角在径向不同龄级间差异极显著,并且存在一定的变化规律。微纤丝角从髓心至树皮随着龄级逐渐增大而减小,管胞长度、管胞宽度和基本密度均由髓心至树皮随着龄级的逐渐增大而增大。此结论与刘青华[28]、谭健晖等[18]、骆秀琴等[29]、毛桃[16]等分别对24年生、22年生、32年生、11年生马尾松木材基本密度、管胞长度、管胞宽度及微纤丝角的研究结果基本一致,也与其他树种的研究结果[30]相似。进一步说明成熟材马尾松木材的基本密度、管胞长度、管胞宽度及微纤丝角在径向上存在一定的变化规律性,总体随着龄级逐渐增大而增大或减小。
3.3 马尾松各性状相关性
林木性状的相关性反映了树木各性状之间的关系,其目的是通过易测的性状指标预测较难测的性状指标,达到间接选择的目标。本研究表明,胸径、树高与材积表型之间具显著相关性,木材基本密度与胸径、材积表型上负相关;管胞长度与管胞宽度,胸径与树高,材积与胸径、树高遗传上正相关。胸径与微纤丝角,基本密度与树高、胸径、材积均为遗传上显著负相关,且生长性状间的相关性高于生长与材性之间的相关性,这与郑仁华等[31]、徐立安等[32]分别对9年生、12年生马尾松性状相关性研究结论一致,说明通过胸径、树高可以对材积进行正向选择,而对胸径、树高进行选择时,基本密度则会受到一定限制。而与范林元等[33]、毛桃[16]对6年生、11年生马尾松相关性研究结论不一致。由此可见,以马尾松幼、中龄林为生长性状与材性相关性研究材料可能会造成因材料年龄偏小而出现的现实误差。
3.4 马尾松优良种源联合选择
联合选择是多个性状综合考虑的一种选择方法,在林木遗传育种中是较为理想的一种选择方法,且投入人力、物力相对较小,选择效率高[34]。本研究通过胸径、树高、材积、管胞长度、管胞宽度、木材基本密度及微纤丝角7个性状构建指数方程,从55个种源中筛选出了5、8、13、40和58号综合性状优良的种源,其各性状分别获得遗传增益为3.69%、5.72%、15.12%、7.05%、-1.12%、0.17%和-6.46%,徐立安等[32]选择出的优良种源分布在广西、广东、江西、湖南、福建、贵州境内的部分种源,与本研究所选择出的优良种源区基本相似,材积生长、木材基本密度和木材微纤丝角等性状的选择效果明显,说明通过成熟林马尾松种源选择可获得一定的遗传增益,能为纸浆材马尾松原料林营建提供速生优质的相对可靠种源,这些种源可在与试验点立地条件类似的地区推广造林。
致谢:安徽省林木种苗总站原站长张晓渡高级工程师,安徽省六安市裕安区林木良种场原副场长胡世荣、技术员王志宽等,中国林业科学研究院亚热带林业研究所秦国峰研究员等对该地理种源试验林营建所做出了贡献。
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
新农业(2021年12期)2021-11-29
绿色科技(2021年21期)2021-11-26
南方农业·中旬(2021年5期)2021-10-25
食品安全导刊(2021年21期)2021-08-30
绿色科技(2020年1期)2020-11-29
农民致富之友(2017年4期)2017-04-10
绿色科技(2016年23期)2017-03-15
绿色科技(2016年17期)2016-12-28
卷宗(2012年5期)2012-10-21
