
银杏胚乳不同发育时期生理代谢变化与其胚性感受态的相关性研究
2022-11-29 11:51马娟娟吴琴霞王瑞敏袁斌龄胡宇辰曹福亮
南京林业大学学报(自然科学版) 2022年4期
马娟娟, 吴琴霞, 陈 颖, 王瑞敏, 袁斌龄, 胡宇辰,曹福亮
(南京林业大学生物与环境学院,南方现代林业协同创新中心,江苏 南京 210037)
银杏(Ginkgobiloba)因集药用、食用、观赏、材用于一体而备受关注[1]。目前银杏育苗方法仍为种子繁殖,其遗传性状易改变,使银杏优良品种的性状难以维持,而良种是银杏工业原料林高产优质的基础[2]。银杏的嫁接、扦插、组织培养是获得良种无性系的最佳方法,但嫁接和扦插繁殖系数低、周期长, 远不能满足市场的需求[3]。因此在短期内获得大量的银杏良种苗木,组织培养特别是体细胞胚发生(somatic embryogenesis,SE)培养技术是快速、省力、规模化繁育林木良种的最有效无性繁殖技术。体胚发生是植物外植体通过一种体外发育程序,将体细胞通过重新编程的方式沿着与有性合子胚发育相似的胚胎发生途径形成体细胞胚的过程[4-7]。
体细胞胚的发生过程可分为3个相对独立的阶段:诱导阶段(胚性细胞的获得)、胚性细胞增殖阶段、体胚发育阶段[8]。其中诱导阶段是植物细胞感受外界的信号刺激(胁迫或外源植物生长调节剂)通过脱分化而获得胚性,形成胚性细胞的过程。植物细胞或组织脱分化或去分化的能力称为植物胚性感受态,它的高低直接决定了能否成功诱导出体细胞胚及难易程度[4]。胚性感受态受外植体的分化状态及外植体类型的影响较大,分化程度越低的细胞去分化能力越强,就越易形成胚性愈伤组织;外植体材料的基因型、发育阶段以及生理状态等决定了其胚性感受态的强弱[9-10]。研究表明,未成熟胚(合子胚的原胚)的胚性感受态明显强于其他阶段[11]。
目前全世界已有多种高等植物通过离体培养获得体细胞胚,落叶松(Larixgmelinii)、火炬松(Pinustaeda)、杂交马褂木(Liriodendronchinense×tulipifera)等林木成熟的体细胞胚胎发生技术体系已经建立,实现了林木新品种的规模化繁育。但作为我国特有树种的银杏目前还没有完全建立繁殖快、规模大、频率高的体细胞再生体系,而诱导具有高度胚性发生能力的愈伤组织是建立银杏体细胞胚培养体系的第一步[11]。另外银杏种子属胚发育未完成型,成熟时(9月)胚还没有完全形成,有的还没有胚,而银杏的胚乳从授粉后就开始生长,比胚发育周期早而持续时间长,是研究银杏胚性愈伤诱导很好的材料。本研究以不同发育阶段胚乳为材料,研究其各时期的生理生化状态及其变化规律,同时以不同发育时期的胚乳为外植体, 进行胚性愈伤组织的诱导,研究银杏胚乳发育状态与其胚性感受态之间的相关性, 筛选出具有较高胚性感受态的银杏胚乳发育阶段,为银杏体细胞胚胎发生体系技术的建立及应用提供依据。
1 材料与方法
1.1 供试材料
供试果实采自南京林业大学校园内(118°49′E,32°04′N)同一地点、生长良好的3棵20年生‘大佛指’银杏树。采种时期分别为6月10日、6月30日、7月20日、8月10日、8月17日、8月24日、8月31日、9月15日8个时间段(激素测定增加7月10日采种样品)。采种后一部分果实放入-70 ℃超低温冰箱中保存,并取胚乳(银杏种子属生理后熟型,在8月下旬种胚才开始发育, 8月下旬—9月的样品中去除胚)用于各类指标测定,另一部分果实取胚乳立即用于愈伤组织的诱导培养。
1.2 胚乳不同发育时期生理指标测定
将上述8个时间段采摘的银杏果实去除外种皮后,取出胚乳进行生理生化分析。含水率、可溶性蛋白含量(考马斯亮蓝法)、过氧化物酶(POD)活性(愈创木酚比色法)、过氧化氢酶(CAT)活性(紫外吸收法)、超氧化物歧化酶(SOD)活性(NBT光化学还原法)的测定均参照文献[12]方法,苯丙氨酸解氨酶(PAL)与总黄酮含量的测定参照邓桂春等[13]的方法,4-香豆酸辅酶A连接酶(4CL)提取及活性测定参照何慕涵等[14]的方法,肉桂醇脱氢酶(CAD)活性的测定参照袁斌玲等[15]方法。
植物激素含量的测定:将9个时间段(增加了7月10日时间段)的银杏果实去除外种皮后,取出胚乳并去除内种皮(皮衣),将种仁研磨成浆状,采用酶联免疫法(ELISA)测定吲哚乙酸(IAA)、赤霉素(GA)、玉米素核苷(ZR)、异戊烯基腺苷(iPA)、脱落酸(ABA)5种激素的含量[16](由中国农业大学实验室协助测定)。
1.3 不同时期银杏胚乳愈伤组织的诱导
1.3.1 外植体消毒接种
6月10日采集的果实中还没形成骨质中种皮,因而对其整个果实进行消毒,之后采集的果实都已形成骨质中种皮,将果实中的外种皮去除后获得银杏种子(6月10日后果实都带有骨质中种皮,也称为白果,去除骨质中种皮和内种皮后称为种仁),对种子进行消毒。先用体积分数70%酒精对种子浸泡60 s,再用质量分数0.3%的高锰酸钾溶液浸泡20 min,期间不断搅拌,将消毒后的种子用无菌水冲洗3~4次,最后用无菌滤纸吸干表面水分。接种时用高压灭菌过的核桃夹夹开种子骨质中种皮,取出种仁,将去除内种皮后的种仁(6月10日—8月10日全部为胚乳,8月17日—9月15日部分含有胚)横放在已灭菌的滤纸上,用不锈钢刀切取种仁外层(胚乳)1~3 mm薄片,平放在培养基上,每瓶放置4片胚乳。
1.3.2 愈伤组织的诱导
①培养基的筛选:对同一时期(6月10日)的胚乳进行基本培养基和调节剂的筛选。选用MS、DCR[17]为基本培养基,附加1.0、1.5 mg/L的萘乙酸(NAA)、2,4-二氯苯氧乙酸(2,4-D),以及0.5、1.0 mg/L 6-苄氨基嘌呤(6-BA),进行正交设计,共8个处理,每个处理6瓶,重复3次。②为研究银杏胚乳诱导率与银杏胚乳发育阶段的相关性,利用筛选出的培养基(MS+1.5 mg/L NAA+1.0 mg/L BA),对6月10日—9月15日8个时间段的胚乳进行愈伤组织诱导及胚性感受态研究。每个时期接种8瓶,3次重复。此后每30 d继代1次。
1.3.3 胚性愈伤组织的诱导
待上述培养过程中诱导的外植体继代2次后,用无菌刀片从带胚乳切片上的愈伤组织轻轻切下每块大约0.5 cm3的愈伤组织,接种到新鲜培养基上进行胚性愈伤组织的诱导。
1.3.4 愈伤组织生长情况的测定
接种15 d后统计不同时期胚乳的污染率,污染率=污染的胚乳片数/接种的胚乳片数×100%。
在各个时期胚乳接种后2个月统计相对应的愈伤组织诱导率。
愈伤组织诱导率=诱导出愈伤组织的胚乳片数/接种的胚乳片数×100%。
1.3.5 培养条件
将接种过的培养瓶于室温下暗培养15 d后再进行15 d光培养,分别在接种后2个月统计各个时期胚乳作为外植体的愈伤组织诱导率,以及胚性愈伤组织的统计。上述培养基中添加蔗糖30 g/L、琼脂6.0 g/L,pH调至5.8左右,培养室温度(25±2)℃,光照周期为12 h/d,光照度为2 000 lx。
1.4 数据处理
试验设置3次重复,采用Excel 2016对数据进行处理,SPSS 21.0软件进行统计分析,方差分析采用Duncan新复极差法(P<0.05为差异显著)。
2 结果与分析
2.1 不同时期银杏果实及种仁生长状态的变化
在6月10日采摘的银杏果实中未发现坚硬状态的中种皮(图1),整个种仁呈翠绿色状态(图1),6月30日的果实中已形成坚硬状态的中种皮。在6月10日—8月31日,外种皮的变化较缓慢,先是颜色上慢慢从深绿往黄色转变,外种皮软硬程度变化也较小,直到8月31日外种皮开始稍有变软,此后到9月15日外种皮的变化较迅速,从黄绿色快速转变为黄棕色,同时外种皮皱缩并带有白粉,且已产生浓重的气味,说明此时果实形态已完全成熟。去除外种皮和骨质中种皮、内种皮的种仁从6月10日的绿色到7月20日逐渐变为黄绿色,且种仁大小不断增加,硬度也更大。8月31日及9月15日的银杏种仁外部形态已经成熟,颜色变成黄色,硬度达到最大且种仁大小基本长到最大(图2)。
2.2 不同时期银杏胚乳含水率和可溶性蛋白含量的变化
种子内胚乳含水率的变化如图3所示,含水率总体呈先下降后保持稳定的状态,在6月10日时,胚乳含水率处于最高值,到6月30日开始下降,8月10日后一直到8月17日胚乳含水率处于稳定状态,8月10日种仁含水率与7月20日之前存在显著差异(P<0.05)。
可溶性蛋白含量整体呈先上升后下降的状态,6月10日胚乳可溶性蛋白的含量处于最低值,之后持续增长,其中7月20日至8月31日可溶性蛋白含量逐渐增加,到8月31日达到最大值;之后可溶性蛋白含量开始下降,9月15日的显著低于8月31日的(图3)。
2.3 不同时期银杏胚乳抗氧化酶活性的变化
不同时期银杏胚乳抗氧化酶活性变化见图4。
超氧化物歧化酶(SOD)活性在前期胚乳中随着采种日期的延后先上升后下降,后期SOD活性处于稳定状态。6月30日SOD活性显著高于6月10日(P<0.05),并达到最大值;随后SOD活性持续下降,在8月17日下降到最低值,比6月30日最大值低68.6%。此后直到9月15日,胚乳中SOD活性变化不大。过氧化物酶(POD)活性的变化在前期处于低活性状态,在6月30日—7月20日有个低水平时期,8月10日显著升高后,该水平一直维持到8月31日,期间变化不大,但到9月15日时POD活性又显著升高,比8月31日活性提高116.9%。胚乳过氧化氢酶(CAT)活性整体处于先下降后增加的趋势,6月30日CAT活性显著低于6月10日的活性,并下降至最低值,之后持续上升,到8月31日达到最大值,之后9月15日POD活性下降,显著低于8月31日。
2.4 不同时期银杏胚乳黄酮合成关键酶活性及总黄酮含量的变化
不同时期银杏胚乳黄酮合成关键酶活性及总黄酮含量的变化见图5。不同发育时期的胚乳中PAL活性在6月10日至8月31日期间处于持续上升状态,在8月31日达到最大值,比最初期6月10日提高了2.16倍。8月31日之后活性又开始下降,9月15日PAL活性显著低于8月31日活性。胚乳的4CL活性和CAD活性变化相似,在整个测定阶段都趋于稳定状态,变化不显著。胚乳总黄酮含量随时间的变化先下降后上升,在7月20日达到最低点后,在8月10日—9月15日缓慢上升,在9月15日达到最大值,比最低点7月20日提高了59.2%。
2.5 不同时期银杏胚乳中5种激素含量的变化
胚乳IAA含量随时间变化如图6所示,6月10日IAA处于较低水平,然后上升,6月30日—8月10日维持6月30日的水平,变化不大。8月17日有所降低,然后8月24日又回升,一直维持到9月15日。GA的水平出现波动式变化,在6月10日—7月10日缓慢上升,然后急剧上升到7月20日的高点,随后又开始下降,在8月10日—8月17日处于较低水平,8月24日出现第2个高点后又逐渐下降并维持到9月15日。
ZR也出现波动式变化,6月10日—7月10日呈上升趋势,在7月10日达到最高点,7月20日后开始下降,并维持此水平至8月24日,然后下降到8月31日并维持到9月15日。iPA的变化是在7月20日之前都处于高水平状态,然后下降到8月10日最低点,在8月17日有所上升,但低于7月20日之前的水平,8月17日—9月15日维持稳定的水平。ABA的含量6月10日开始上升至7月20日达到最高点,8月10日稍有下降,该水平一直维持到8月24日,8月31日—9月15日又出现显著下降,8月31日时的ABA含量比最高点7月10日下降41.5%。
从IAA与ABA质量比(记为IAA/ABA)来看,6月30日、8月31日 和9月15日3个时期都处于高水平,与其他时期有显著的差异,6月30日为最高点,其他时期6月10日、7月10日、8月10日—8月24日3个时期处于中等水平,7月20日处于最低的水平,但与7月10日、8月10、8月17日、8月24日都没有显著差异(图6)。
(ZR+iPA)与ABA的质量比[记为(ZR+iPA)/ABA]与IAA/ABA有所不同,6月10日—8月10日呈逐渐下降趋势,到8月10日达到最低点,然后8月17日逐渐回升至8月31日,此时的(ZR+iPA)/ABA与7月10日的水平无显著差异,9月15日稍有下降与8月24日持平(图6)。
2.6 不同时期银杏胚乳组织培养污染率与愈伤组织诱导率的变化
选择6月10日取材的银杏胚乳用于培养基的筛选。从表1可以看出,基本培养基类型、调节剂组合影响胚乳愈伤组织的诱导率。从8个处理中可以看出,银杏胚乳在MS+1.5 mg/L 2,4-D+1.0 mg/L BA的配方中愈伤组织诱导率最高,但其诱导出的愈伤组织较容易褐化。而MS+1.5 mg/L NAA+1.0 mg/L BA既具有较高水平的愈伤组织诱导率,又有较低的褐化率,选用此配方较好(表1)。

表1 不同培养基对银杏胚乳愈伤组织诱导率的影响
对上述8个时间段各自的胚乳同时在MS+1.5 mg/L NAA+1.0 mg/L BA培养基上进行愈伤组织诱导,发现不同时期外植体污染率存在较大差异。出现较高污染率的时间段主要集中在8月,其4个时间段的污染率为6.25%~11.46%,差别不大。而6、7和9月外植体污染率水平较低(表2)。
光照培养下,胚乳切片接种后,其表面先慢慢失水,表皮颜色逐渐加深变成褐色,接种后30 d左右褐色的胚乳表皮切口边缘处逐渐有愈伤组织诱导出来。继代后60 d时统计,不同发育时间段的胚乳愈伤组织的诱导率出现较大差异。在6月10日时,愈伤组织诱导率处于一个较高的水平,但此后出现较大幅度的下降,在6月30日、7月20日诱导率处于较低的水平,7月20日的愈伤组织诱导率最低,仅为6月10日的43.57%。但从8月10日开始,愈伤组织诱导率开始迅速上升,8月24日—9月15日愈伤组织诱导率均在88%以上(表2)。
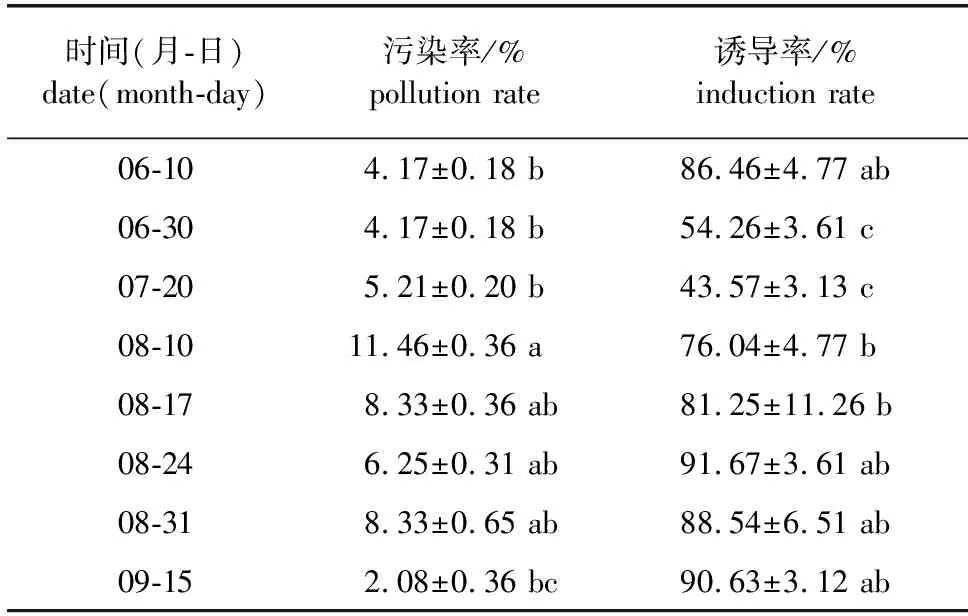
表2 8个时间段银杏胚乳外植体组织培养污染率与愈伤组织诱导率的变化
2.7 胚性愈伤组织的诱导及生长
将初诱导60 d后的愈伤组织从原胚乳切片上取下,转接到MS+1.5 mg/L NAA+1.0 mg/L 6-BA +0.25 g/L MES+0.2g/L谷氨酸的胚性愈伤组织诱导培养基中,在继代第4次后各个时期胚乳所培养的愈伤组织差别明显。6月10日的胚乳培养4周后逐渐从切口处出现较多的白色或者淡绿色结构较为松散的非胚性愈伤组织,继续培养后易褐化死亡。6月30日的胚乳接种情况和7月20日基本相同,愈伤组织诱导率较低,在培养过程中也出现褐化现象(图7A—7D)。8月17日—9月15日的胚乳愈伤组织诱导率较高(图7E—7H),且继代培养4次后,逐渐形成黄白色、结构较为紧密、表面呈颗粒状的胚性愈伤组织,在体视显微镜下可观察到愈伤组织中许多晶莹透亮的圆球体。此阶段是胚性愈伤组织发生率最高的时期。
3 讨 论
3.1 银杏胚乳不同发育时期与胚性感受态的关系
与被子植物不同, 裸子植物体细胞胚诱导对外植体的要求更加严格,诱导体细胞胚胎发生的最合适外植体可能因物种而异[18]。以胚性感受态较高的幼胚为外植体进行体胚诱导,幼胚胚性感受态需在特定条件下才能被激活的现象与植物幼胚体内的生理状态密切相关[19]。胚性感受态的高低直接决定了植物体细胞胚诱导的难易程度[20]。因此探究外植体胚性感受态的高低变得尤为重要。本研究发现银杏胚乳愈伤组织诱导率分为3个阶段:第1个阶段为6月10日—6月30日,银杏早期未成熟状胚乳,易诱导出愈伤组织,胚乳的愈伤诱导率高;第2阶段为胚乳发育的中期,6月30日—8月10 日,此时胚乳愈伤组织诱导率比6月10日、8月17日后都低,并伴有一定程度的褐化;第3阶段为胚乳发育后期,8月17日—9月15日,愈伤组织诱导率提高,愈伤组织多黄白色,表面具球形小颗粒的胚性愈伤组织团。早期的胚乳愈伤组织诱导率高,其原因可能是此时有较高的水分含量(6月10日左右),种子正处于旺盛的生长阶段,细胞生理代谢旺盛,但发现此时愈伤组织的形成是整个胚乳切片的愈伤化而成,愈伤组织结构松散,为非胚性愈伤组织,不易成团,继续培养后愈伤组织已褐化死亡。中期的胚乳(6月30日—8月10 日)愈伤组织诱导率比早期和后期的都低,后期愈伤组织也有一定程度的褐化,但也能诱导出颗粒状的胚性愈伤组织。其可能的原因是此时正是种子外种皮形成的时期,其含水量开始下降,而蛋白质开始上升。到8月中旬后,银杏种子生长趋于缓慢,胚乳趋于成熟,其含水量下降并保持一个稳定的状态,有利于胚性愈伤组织的形成。另外8月10日后胚乳可溶性蛋白含量处于一个上升的过程,吕雪芹等[21]通过对不同时期银杏胚乳可溶性蛋白含量的测定也得出相同的结果。胚乳中积累一定含量的可溶性蛋白能提高胚乳的胚性感受态,有利于诱导胚性愈伤组织的形成。因此,胚乳不同发育时期胚性感受态强弱和胚性愈伤组织诱导能力的排列顺序为:第3阶段>第2阶段>第1阶段。
3.2 胚乳发育过程中3种抗氧化酶与胚性感受态的关系
外植体抗氧化酶活性的变化与外植体细胞的分化有直接的关系[22]。本研究中胚乳发育时期的抗氧化酶SOD、POD、CAT酶活性与胚乳愈伤组织诱导有较大相关性。早期(6月10日—6月30日)胚乳的SOD酶具有较高活性,而POD 和CAT 的活性较低,SOD活性在6月30日达到最大值,然后到7月20日下降至最低,而7月20日之后POD和CAT活性逐渐升高,在后期阶段(8月10日—9月15日)胚乳POD和CAT处于高活性水平。这表明胚乳在生长前期需要大量的能量,从而产生较高的线粒体活性以及过氧化物[23]。前期高水平的SOD酶活性可能是由于此时随着外界温度和光照的增加,银杏果实快速生长,但也会由于温度和光照增加产生较多的超氧自由基,SOD 酶活性较高有利于将其清除。在后期胚乳含水量降低,环境胁迫加强,通过POD和CAT活性逐渐升高以清除前期产生的H2O2,另外一定的环境胁迫可提高胚乳愈伤组织的诱导,且多为颗粒状的胚性愈伤组织,说明后期高活性的POD和CAT有利于外植体胚性感受态的升高。
3.3 胚乳发育过程中黄酮变化与胚性感受态关系
黄酮是银杏植株内一种重要的多酚类次生代谢物,对增强植物抗逆性有较大的作用,有很强的生物活性[24-25]。本研究中胚乳发育的过程中,与黄酮代谢相关的酶4CL(4-香豆酸辅酶A连接酶)和CAD(肉桂醇脱氢酶)活性变化较小,不同阶段间差距不大。因此,胚乳中黄酮的积累主要是PAL(苯丙氨酸解氨酶)在起主要作用,高PAL活性有利于黄酮的积累。而中期(6月30日—7月20日)的胚乳愈伤组织诱导率低可能是因为总黄酮含量处于较低水平。后期(8月31日—9月15日)的总黄酮含量达到高峰值,此时的胚乳愈伤诱导率较高,且愈伤组织生长状态较好,已形成胚性愈伤组织,由此可见,高的黄酮含量也是银杏胚乳具有较高胚性感受态的一个因素。
3.4 胚乳发育过程中5种激素含量变化及其比值与胚性感受态的关系
植物的内源激素是影响组织胚性感受态的最重要因素之一[26]。许多学者研究表明,在植物离体细胞发育过程中,生长素起重要作用,比如启动细胞分裂和诱导胚性潜力,从而促进外植体早期发育[27]。本研究中,6月10日的胚乳IAA含量最低,其余各发育时期基本维持在较高水平。从各时期胚乳诱导愈伤组织的情况来看,高水平的IAA对胚性愈伤组织的诱导有重要意义。GA含量在前至中期一直较高,8月31日之后处于较低水平,此时胚乳最易诱导胚性愈伤,推测高含量GA3可能不利于胚性细胞的产生, 这与多数植物体胚的研究结果[28]类似。ZR和iPA含量的变化趋势较为相似,中期(6月30日—7月20日)的含量较高,这个阶段内的胚乳愈伤组织率较低;而8月之后这两种激素水平都较低,愈伤诱导情况较好,说明低浓度的ZR和iPA更有利于胚性愈伤组织的诱导。ABA的含量在6—9月呈先上升后下降的趋势,从各时期胚乳诱导愈伤组织的情况推测,胚性愈伤组织的发生需要较低水平的ABA。
郭玉琼等[29]认为植物内源激素对植物生长发育的调控作用不仅与各种内源激素的含量有关,而且与各类激素在植物体内的相互平衡有密切关系。本研究发现愈伤组织诱导率及胚乳的胚性感受态更受IAA/ABA与(ZR+iPA)/ABA的调控。IAA/ABA在早期处于高水平,而中期(6月30日—7月20日)处于低水平,后期(8月10日—9月15日)又处于高水平,与胚乳愈伤组织诱导率成正相关。而(ZR+iPA)/ABA在早期至中期处于高水平,后期(8月10日之后)处于低水平,有利于胚乳愈伤组织的诱导。
综上所述,银杏胚乳的发育状态与其愈伤组织诱导和胚性愈伤组织形成具有较强的相关性,较高的POD、CAT活性、黄酮含量、IAA/ABA,及较低(ZR+iPA)/ABA是银杏胚乳诱导胚性愈伤组织必需的条件。8月17日—9月15日是银杏诱导胚性愈伤组织的最佳时期,特别7月20日—9月15日是胚乳较高的胚性感受态时期。银杏胚乳发育过程及其与胚性愈伤组织的关联性机制还需进一步研究。
猜你喜欢
粮油与饲料科技(2022年2期)2022-11-24
中国兽医学报(2022年5期)2022-11-16
山东林业科技(2022年3期)2022-07-14
烟台果树(2021年3期)2021-11-29
三农资讯半月报(2021年2期)2021-03-22
辽宁农业科学(2021年1期)2021-03-17
学生导报·东方少年(2019年3期)2019-05-14
生物学教学(2018年5期)2018-11-29
小学阅读指南·高年级版(2016年9期)2016-10-31
农家顾问(2014年4期)2014-06-26
