
刈割对入侵植物牛膝菊生长和繁殖特征的影响
2022-12-16 11:35初丽爽李海燕杨允菲
植物资源与环境学报 2022年6期
初丽爽, 李海燕,①, 杨允菲, 郭 健
(1. 东北师范大学: a. 草地科学研究所, b. 植被生态科学教育部重点实验室, c. 吉林松嫩草地生态系统国家野外科学观测研究站, 吉林 长春 130024; 2. 徐州工程学院环境工程学院, 江苏 徐州 221018)
表型可塑性是衡量植物适应环境能力的重要指标[1],包括形态和繁殖可塑性等,面对不同的环境因子,植物通过适当调整生长和繁殖特征,最大限度地适应环境,降低环境干扰带来的负面影响,实现种群延续[2-4]。入侵植物进入新栖息地后往往能快速生长、繁殖和扩散,这离不开其自身较高的可塑性和生态适应性[5]。与本土植物相比,入侵植物表现出强大的竞争力和繁殖力,可通过调整自身的资源分配适应入侵地环境[5,6],并通过向周围释放化学物质产生化感作用,抑制本土植物生长发育[7,8];此外,入侵植物还会改变土壤养分平衡,影响土壤微生物多样性,使土壤环境更有利于自身生长,严重威胁生态系统及生物多样性[9,10]。豚草(AmbrosiaartemisiifoliaLinn.)[11]、加拿大一枝黄花(SolidagocanadensisLinn.)[12]和微甘菊(MikaniamicranthaH. B. K)[13]等菊科(Asteraceae)植物已成为极具危害性的入侵植物。因此,开展入侵植物生长和繁殖特征研究,对控制入侵植物和保护生物多样性具有重要意义[14]。
刈割是草地利用管理和有害植物控制的常用方式之一[15]。草地生产中,适当刈割可充分发挥植物均衡性生长和补偿性生长作用[16]。刈割对草地植物的影响因强度、时期和频次等不同而异[17]。研究发现,与不刈割相比,适宜的刈割不仅可以增加草地植物地上生物量和营养价值[18,19],还可以抑制杂草生长[20]。对多年生入侵植物互花米草(SpartinaalternifloraLois.)进行刈割可阻止其地上部分生长及生物量累积,降低其光合能力,抑制植株生长[21,22]。孕穗期低位刈割能明显减少一年生入侵植物少花蒺藜草(CenchrusspinifexCav.)的结实数量[23]。因此,有效的刈割能够明显抑制入侵植物尤其是一年生入侵植物的生长、繁殖和扩散。
牛膝菊(GalinsogaparvifloraCav.)为牛膝菊属(GalinsogaRuiz et Pav.)一年生草本植物,其适应环境广泛,具有种子量大、生长迅速和易扩散等特点,现已扩展至中国33个省份(地区),严重影响农田作物、蔬菜和果树生长[24]。在中国东北地区,牛膝菊的入侵主要危害草坪、绿地、疏林、果园和蔬菜地等[25]。现有研究主要通过化学防除、生物防治和植物化感作用等方式抑制牛膝菊生长[26-28],刈割对牛膝菊生长和繁殖特征的影响以及其生长的补偿性反应和防控扩散的有效性尚不明确,仍有待进一步研究。
鉴于此,本研究基于植物种群表型可塑性调节的核心理论地位,通过3个强度和3个频次的刈割处理,研究刈割对牛膝菊生长和繁殖特征的影响,并比较不同刈割处理下牛膝菊干质量补偿指数的差异,进而探究刈割对牛膝菊防治的效果及可行性,以期揭示刈割下牛膝菊的表型可塑性调节策略,为入侵植物的有效防控提供科学依据。
1 材料和方法
1.1 材料
实验于2018年5月至8月在东北师范大学试验基地的温室大棚(东经125°19′、北纬43°52′)内进行。供试牛膝菊种子于2017年秋季采集自东北师范大学校园内,保存于阴凉干燥处。实验采用盆栽方式进行,土壤为长春市周边农田的壤土,总氮和总磷含量分别为0.09和0.06 mg·g-1,pH 6.6。于2018年5月末,在每个花盆(内径25 cm、高20 cm)中播种10粒种子,共播种60盆,定期浇水、除草。6月中旬定苗,选择大小基本一致的幼苗,每盆留4株幼苗。
1.2 方法
1.2.1 实验设置 参考文献[29]中的方法设置刈割强度和刈割频次,其中,刈割强度分为剪除株高的1/4(轻度刈割)、1/2(中度刈割)和3/4(重度刈割),刈割频次分为1、2和3次,未刈割处理为对照组(CK),共10个处理组,每个处理组6盆(即6个重复)。在牛膝菊株高达到15 cm时(6月23日)进行第1次刈割,第2和第3次刈割分别在第1次刈割后的15 d(7月8日)和30 d(7月23日)进行。
1.2.2 测定指标 8月初开始采集各处理组每株牛膝菊的成熟花序,8月末对牛膝菊进行整株取样测定,记录不同处理组每株牛膝菊的一级分枝数和二级分枝数,采用卷尺(精度0.1 cm)测量一级分枝长(单株所有一级分枝基部至顶部距离的总和)和二级分枝长(单株所有二级分枝基部至顶部距离的总和)。测量后将植株分为根、茎、叶、花序和种子,统计每株牛膝菊的种子数,之后将各组织置于65 ℃烘箱中烘干至恒质量,使用千分之一天平称量每株牛膝菊的根、茎、叶、花序和种子的干质量及百粒质量。根据公式“总干质量=根干质量+茎干质量+叶干质量+花序干质量”计算总干质量;根据公式“某器官干质量分配=(该器官干质量/总干质量)×100%”计算各器官干质量分配;根据公式“生殖分配=(花序干质量/总干质量)×100%”计算生殖分配;根据公式“干质量补偿指数(CI)=刈割处理组的总干质量/对照组的总干质量”计算干质量补偿指数,其中,CI>1属于超补偿生长,CI=1属于等补偿生长,CI<1属于欠补偿生长。
1.3 数据处理和分析
使用EXCEL 2016和SPSS 23.0软件对数据进行统计分析,采用双因素方差分析对刈割频次和刈割强度的主效应及交互效应进行检验,采用单因素方差分析和LSD法进行方差分析和多重比较(α=0.05)。对干质量补偿指数与“1”进行独立样本t检验[30]。使用Origin 2021软件作图。
2 结果和分析
2.1 刈割处理对牛膝菊生长特征的影响
双因素方差分析结果(表1)显示:刈割强度和刈割频次对牛膝菊一级分枝数、二级分枝数、一级分枝长和二级分枝长有极显著(P<0.01)或显著(P<0.05)影响,二者的交互作用对牛膝菊一级分枝数和二级分枝数有极显著影响。
由图1可见:总体上看,与对照(未刈割)相比,刈割处理会减少牛膝菊分枝数,促进其分枝伸长。在相同的刈割频次下,随着刈割强度的增加,牛膝菊一级分枝数和二级分枝数总体呈降低的趋势,一级分枝长和二级分枝长总体呈升高的趋势。随着刈割频次的增加,轻度刈割(剪除株高的1/4)的牛膝菊二级分枝数和一级分枝长呈降低的趋势,二级分枝长呈先增加后降低的趋势,刈割2和3次的牛膝菊二级分枝数和一级分枝长总体显著低于刈割1次,二级分枝长显著高于刈割1次;中度刈割(剪除株高的1/2)的牛膝菊一级分枝长和二级分枝长呈先增加后降低的趋势,刈割2次的牛膝菊一级分枝长和二级分枝长显著高于刈割1次;重度刈割(剪除株高的3/4)的牛膝菊一级分枝数和一级分枝长无显著变化,刈割2和3次的牛膝菊二级分枝数显著低于刈割1次,二级分枝长显著高于刈割1次。此外,与其他刈割频次比,同一强度刈割2次的牛膝菊二级分枝数最小,二级分枝长最大。
2.2 刈割处理对牛膝菊干质量及繁殖特征的影响
2.2.1 对牛膝菊干质量及种子数的影响 双因素方差分析结果(表2)显示:刈割强度对牛膝菊茎干质量、花序干质量、总干质量、种子干质量、百粒质量和种子数有极显著(P<0.01)或显著(P<0.05)影响,刈割频次对牛膝菊根干质量、茎干质量、叶干质量、花序干质量、总干质量、种子干质量、百粒质量和种子数有极显著影响,二者的交互作用对牛膝菊根干质量、总干质量、种子干质量、百粒质量和种子数有极显著或显著影响。

表1 刈割强度和刈割频次对牛膝菊生长特征影响的双因素方差分析1)



由图2可见:总体上看,与对照(未刈割)相比,刈割处理会减少牛膝菊茎干质量和叶干质量,刈割3次会明显减少牛膝菊花序干质量,增加根干质量。在相同的刈割频次下,随着刈割强度的增加,牛膝菊茎和叶干质量总体呈降低的趋势。刈割1和2次的牛膝菊根干质量和叶干质量在不同刈割强度间无显著差异。在相同的刈割强度下,随着刈割频次的增加,牛膝菊根干质量呈升高的趋势,茎干质量、叶干质量和花序干质量总体呈降低的趋势,轻度刈割(剪除株高的1/4)3次的牛膝菊根干质量最大,3种刈割强度的牛膝菊叶干质量在不同刈割频次间无显著差异,刈割3次的牛膝菊花序干质量最小。此外,轻度刈割下,刈割3次的牛膝菊总干质量显著高于其他刈割频次,而中度刈割(剪除株高的1/2)和重度刈割(剪除株高的3/4)下,牛膝菊的总干质量在不同刈割频次间无显著差异。

表2 刈割强度和刈割频次对牛膝菊干质量及种子数影响的双因素方差分析1)







: 未刈割No mowing; : 轻度刈割(剪除株高的1/4) Light mowing (cutting off 1/4 of plant height); : 中度刈割(剪除株高的1/2) Moderate mowing (cutting off 1/2 of plant height); : 重度刈割(剪除株高的3/4) Severe mowing (cutting off 3/4 of plant height). 不同小写字母表示同一刈割频次下不同刈割强度之间差异显著(P<0.05)Different uppercases indicate the significant (P<0.05) difference among different mowing intensities under the same mowing frequency; 不同大写字母表示同一刈割强度下不同刈割频次之间差异显著(P<0.05)Different lowercases indicate the significant (P<0.05) difference among different mowing frequencies under the same mowing intensity.
由图2还可见:不同的刈割处理对牛膝菊繁殖特征的影响表现出较大差异。刈割1次的牛膝菊种子干质量和种子数在不同刈割强度间无显著差异;轻度刈割2次的牛膝菊种子干质量和种子数最少,轻度刈割3次的牛膝菊种子干质量和种子数最大且显著高于其他处理组。此外,与对照相比,同一频次中度和重度刈割的牛膝菊百粒质量显著降低。
2.2.2 对牛膝菊各组织干质量分配的影响 双因素方差分析结果(表3)显示:刈割强度对牛膝菊根干质量分配、茎干质量分配和生殖分配有极显著或显著影响,刈割频次对牛膝菊根干质量分配、茎干质量分配、叶干质量分配和生殖分配有极显著影响,二者的交互作用对牛膝菊生殖分配有极显著影响。
由图3可见:总体上看,与对照相比,重度刈割(剪除株高的3/4)或刈割3次会显著减少茎干质量分配,增加根干质量分配。刈割1和3次的牛膝菊根干质量分配、茎干质量分配、叶干质量分配和生殖分配在不同刈割强度间无显著差异。随着刈割强度的增加,刈割2次的牛膝菊根干质量分配和生殖分配明显升高,茎干质量分配总体显著降低;与对照相比,不同强度刈割3次的牛膝菊根干质量分配显著升高,茎干质量分配、叶干质量分配和生殖分配总体显著降低。在相同的刈割强度下,刈割3次的牛膝菊根干质量分配显著高于其他刈割频次,茎干质量分配、叶干质量分配和生殖分配总体显著低于其他刈割频次。
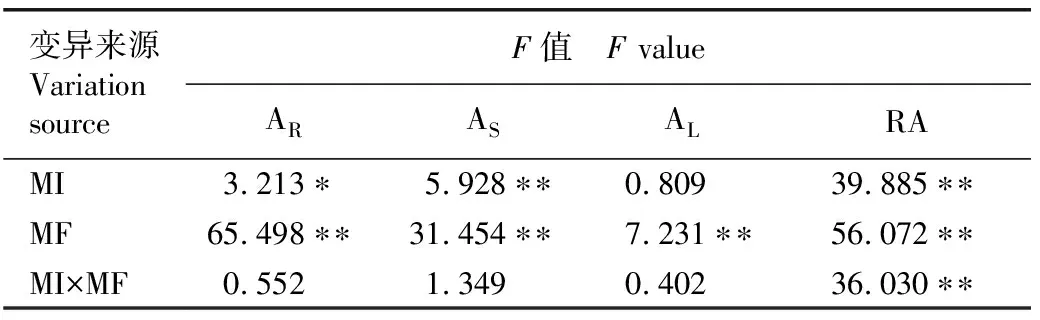
表3 刈割强度和刈割频次对牛膝菊干质量分配影响的双因素方差分析1)


2.3 不同刈割处理下牛膝菊干质量补偿指数的比较
不同刈割处理下牛膝菊干质量补偿指数见表4。结果显示:不同刈割处理下牛膝菊的补偿生长能力不同。3种刈割频次的牛膝菊干质量补偿指数总体随着刈割强度的增加而降低。与轻度刈割(剪除株高的1/4)和中度刈割(剪除株高的1/2)相比,重度刈割(剪除株高的3/4)2和3次会显著(P<0.05)降低牛膝菊的干质量补偿指数。轻度刈割3次的牛膝菊干质量补偿指数显著高于刈割1和2次,中度刈割和重度刈割的牛膝菊干质量补偿指数在不同刈割频次间无显著差异,中度刈割2次的牛膝菊干质量补偿指数为1.00,表现为等补偿生长;轻度刈割和中度刈割3次的牛膝菊干质量补偿指数分别为1.18和1.01,表现为超补偿生长,但差异不显著;其他处理组的牛膝菊干质量补偿指数均小于1,表现为欠补偿生长,其中,轻度刈割2次和重度刈割1、2和3次均会使牛膝菊出现显著的欠补偿生长。

表4 不同刈割处理下牛膝菊干质量补偿指数
3 讨 论
3.1 牛膝菊的生长与形态可塑性
入侵植物在生长特征方面对刈割等干扰具有一定的耐受性,针对不同的刈割强度和刈割频次,植物会通过再生对损伤产生不同的响应[9]。本研究中,刈割处理会影响牛膝菊的生长特征,总体上看,与对照(未刈割)相比,刈割处理会减少牛膝菊分枝数,促进其分枝伸长。与吕林有等[23]对一年生少花蒺藜草的刈割研究结果一致,即在植物生长早期进行低位刈割后,受损植株能迅速进行伸长生长。但也有研究发现,一年生入侵植物黄顶菊〔Flaveriabidentis(Linn.) O. Kuntze〕刈割后会增加其分枝数[31]。这可能是由于与黄顶菊表现为茎直立不同,牛膝菊接近地表的枝条会进行土壤的自然压条繁殖使种群可以快速增长[24]。上述研究表明:植物自身的形态特征对于刈割干扰的响应具有决定性作用。
有研究指出,放牧或刈割处理会影响草地生态系统的干质量,且可能影响植物的生物量分配[31]。本研究中,整体上看,高强度〔重度刈割(剪除株高的3/4)〕或高频次(刈割3次)的刈割处理会明显减少牛膝菊茎干质量和叶干质量以及茎干质量分配和叶干质量分配,这可能是由于植物叶片的大量损失,导致其光合能力急剧下降,影响了植株地上部分的生长[28]。对多年生入侵植物五爪金龙〔Ipomoeacairica(Linn.) Sweet〕进行刈割处理,发现当去叶强度大于50%时,地上生物量才会显著降低[32]。此外本研究发现,刈割1和2次基本不会影响根部生长,但刈割3次会使牛膝菊根干质量及根干质量分配总体显著增加,说明通过多次刈割去除植物地上部分,会严重影响牛膝菊的存活,导致植物将更多资源转移至根部以维持自身生存;另一方面,刈割处理使牛膝菊分枝伸长产生压条繁殖,从而增加了须根的干质量。
3.2 牛膝菊的繁殖可塑性
入侵植物具有强大的繁殖能力,部分入侵植物在有性生殖和无性繁殖上的投入均大于原产地种群[33],具有很强的竞争优势和较高的逆境适应能力[34]。本研究中,刈割频次和刈割强度对牛膝菊种子数和种子干质量等有性生殖特征存在显著影响。与对照(未刈割)相比,刈割1和2次的牛膝菊花序干质量无显著变化,但生殖分配总体显著提高;刈割3次的牛膝菊花序干质量出现降低趋势,且生殖分配和种子百粒质量显著降低,说明一年生的牛膝菊可以通过增加对有性生殖的投入而快速调整自身的有性生殖策略以适应外界的不良干扰,增加刈割频次可能会有效控制牛膝菊的有性生殖。
3.3 牛膝菊生长和繁殖特征间的权衡
生物不会使生活史的每一组分都达到最大,必须在生活史的不同组分间进行权衡,如生长和繁殖间的权衡[35]。有研究表明:在寄生胁迫条件下,入侵植物空心莲子草〔Alternantheraphiloxeroides(Mart.) Griseb.〕会减少对自身营养生长的投入,将更多的资源投向克隆繁殖,增强自身防御能力,以利于后代的生存和繁衍[36]。本研究结果表明:与对照(未刈割)相比,刈割1和2次的牛膝菊茎干质量总体显著减少,叶干质量明显减少,根干质量无显著变化,生殖分配显著增加;而刈割3次的牛膝菊根干质量总体上显著增加,花序干质量明显减少,生殖分配显著减少,说明在面对刈割干扰时,牛膝菊种群存在生长和繁殖间的权衡。牛膝菊产生不定根,用于吸收土壤中的营养和水分,茎和叶是植物生长所必需的营养器官,花序是生殖器官。当低频次刈割(刈割1和2次)造成牛膝菊营养器官的损失时,植株在保证自身存活和生长的同时,会优先分配资源到繁殖器官来保证种群的延续,从而抵抗刈割带来的不良影响,但高频次刈割(刈割3次)会造成营养器官多次损失,使牛膝菊不得不减弱有性生殖以保证自身的生存。
生活史理论认为,在一个有限的资源环境中,植物不能同时产生大且多的种子,即种子的大小与数量之间存在权衡关系[37],同时种子大小与数量会受植物的个体大小和环境因子的双重影响[38]。本研究结果发现,轻度刈割3次的牛膝菊种子干质量和种子数最大且显著高于其他处理组,而百粒质量显著低于对照,重度刈割3次的牛膝菊种子干质量和种子数明显低于对照。说明牛膝菊在面对外界频繁干扰时会首先产生小而多的种子,以增加自身的适合度,实现种群的扩散和延续,但当干扰超过一定强度时,牛膝菊的有性生殖指标会受到抑制。
3.4 牛膝菊的补偿生长与入侵防除
补偿生长是植物对伤害的一种积极反应,通常关注的是植物在生物量生产方面的补偿[39,40]。随着刈割强度的增加,重度刈割的牛膝菊表现为显著的欠补偿生长。综合有性生殖特征的变化发现,与对照(未刈割)相比,重度刈割3次会显著降低牛膝菊的总干质量、花序干质量、百粒质量以及生殖分配,可有效控制牛膝菊的生长和繁殖,减少其干质量。因此,在对牛膝菊的实际防控中,持续的重度刈割可以作为一种有效控制牛膝菊生长和蔓延的方式加以利用。
4 结 论
刈割处理能够减少牛膝菊的分枝数,但会促进各级枝条的伸长。在不同刈割条件下,牛膝菊会不断调整对有性生殖的投入以延续种群。3种刈割强度下,刈割1和2次会使牛膝菊将资源优先分配到繁殖器官;刈割3次会使牛膝菊生殖分配显著降低,且随着刈割强度的增加,其补偿指数降低。多次重度刈割是降低牛膝菊生长,抑制其有性生殖,进而实现对牛膝菊防控的重要方式。
猜你喜欢
世界科学技术-中医药现代化(2022年9期)2023-01-17
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
中国农业科学(2022年13期)2022-07-26
农业机械学报(2020年3期)2020-04-01
江苏农业科学(2020年21期)2020-03-08
种子(2019年5期)2019-07-02
中成药(2018年6期)2018-07-11
中学生阅读(初中版)(2016年11期)2017-01-13
天然产物研究与开发(2016年11期)2016-06-15
考试周刊(2016年11期)2016-03-17
