
人为干扰对乌溪江流域山蜡梅种群结构与动态的影响
2022-12-16 11:35龚笑飞余水生张川英袁位高朱锦茹姚良锦李婷婷唐敏俊
植物资源与环境学报 2022年6期
龚笑飞, 余水生, 张川英, 袁位高, 朱锦茹, 姚良锦, 李婷婷,①, 唐敏俊
(1. 浙江省遂昌县生态林业发展中心, 浙江 遂昌 323300; 2. 浙江省林业科学研究院, 浙江 杭州 310023)
山蜡梅(ChimonanthusnitensOliv.)隶属于蜡梅科(Calycanthaceae)蜡梅属(ChimonanthusLindl.),为常绿灌木,常生于山地疏林中或石灰岩山地,主要分布于皖、浙、苏、赣、闽、湘、鄂、云、贵等省[1,2]。山蜡梅含有多种活性成分(如黄酮类、生物碱和香豆素等)[3],其根和叶都具有极高的药用价值,可镇咳祛痰、消毒抑菌、降压、增强免疫,是中国宝贵的中药资源[4]。在农林活动活跃区,山蜡梅野生资源常因竹林和其他树种的抚育而被大量砍伐,加上过度采挖,其野生资源大量减少,自然更新日益困难[5]。
植物的种群结构能够反映其在时间和空间上的变化规律,从而体现种群动态变化趋势及群落演替趋势[6-8]。植物种群的年龄结构能够反映其生存现状、与环境的适合度以及受干扰状态,预测种群的未来发展趋势,对指导种群的人工复壮、更新,恢复种群稳定性具有重要意义[9-11]。湖南[12]和江西[13]等地已经先后展开了对山蜡梅野生资源的调查工作,但关于山蜡梅的种群结构和动态特征及外界干扰对山蜡梅种群结构的影响尚不清楚,亟待研究。
浙江省遂昌县乌溪江沿岸分布着大片以山蜡梅为主、伴生柳叶蜡梅(C.salicifoliusHu)的植物群落。笔者初步调查发现,虽然山蜡梅为这些群落的优势种,但在人为干扰下山蜡梅的种群数量大幅下降,种群结构的动态平衡遭到严重破坏。为了保护这一山蜡梅野生资源,笔者对该区域天然次生林和人为干扰林中山蜡梅的个体数量和植株大小进行了调查,并采用多种种群生态学方法分析了该区域山蜡梅野生种群的结构和数量变化动态,预测了种群的未来发展趋势,以期揭示乌溪江流域山蜡梅野生种群的生存现状和受干扰程度,评估该山蜡梅种群与自然环境的适合度,为有效保护和合理管理山蜡梅野生资源、保证山蜡梅野生资源的可持续利用提供理论依据。
1 研究区自然概况和研究方法
1.1 研究区自然概况
浙江省遂昌县的乌溪江流域(东经118°55′36″~119°00′32″、北纬28°21′21″~28°37′13″)全长约56 km,海拔258~318 m,属亚热带季风气候,多年平均气温16.8 ℃,最高气温40.6 ℃,最低气温-9.9 ℃,全年雨水充足。该区域林分类型多样,土壤为红壤、黄壤和岩性土,呈酸性,有机质丰富,速效钾和水解氮含量中等到丰富,速效磷较为缺乏。群落内乔木层植物有杉木〔Cunninghamialanceolata(Lamb.) Hook.〕、苦槠〔Castanopsissclerophylla(Lindl.) Schott.〕、石栎〔Lithocarpusglaber(Thunb.) Nakai〕、青冈〔Cyclobalanopsisglauca(Thunb.) Oerst.〕、黄瑞木〔Adinandramillettii(Hook. et Arn.) Benth. et Hook. f. ex Hance〕和水杉(MetasequoiaglyptostroboidesHu et Cheng)等;灌木层植物有山蜡梅、杜茎山〔Maesajaponica(Thunb.) Moritzi. ex Zoll.〕、檵木〔Loropetalumchinense(R. Br.) Oliv〕、峨眉鼠刺(IteaoblongaHand.-Mazz.)、香花崖豆藤(MillettiadielsianaHarms)、白花苦灯笼〔Tarennamollissima(Hook. et Arn.) Rob.〕和水团花〔Adinapilulifera(Lam.) Franch. ex Drake〕等;草本层植物有狗脊蕨〔Woodwardiajaponica(Linn. f.) Sm.〕、美丽复叶耳蕨〔Arachniodesspeciosa(D. Don) Ching〕、里白〔Diplopterygiumglaucum(Thunb. ex Houtt.) Nakai〕、芒萁〔Dicranopterispedate(Houtt.) Nakaike〕、山类芦(NeyraudiamontanaKeng)和淡竹叶(LophatherumgracileBrongn.)等。
1.2 研究方法
1.2.1 样地设置及植物调查 于2021年6月至10月在浙江省遂昌县境内由北向南踏查乌溪江两岸的山蜡梅分布情况。在乌溪江流域不同区段,选择山蜡梅分布较为集中的区域,选择6个地点设置样地,依次编号P1至P6。根据实际地形划分样地,样地面积均为400 m2,每个样地平均分成4个样方。其中,P1、P2和P3样地位于乌溪江两岸的陡峭林地中,为未经人为干扰的天然次生常绿阔叶林(简称天然林),林分密度为1 800 hm-2;P4、P5和P6样地位于人为间伐抚育后的常绿阔叶林内(简称干扰林),林分密度为1 530 hm-2。各样地的基本概况见表1。测量每个样方内所有山蜡梅的基径、株高、冠幅等指标,并对样地内所有山蜡梅进行单株拍照和定位。

表1 供试乌溪江流域各样地的基本概况
1.2.2 种群结构分析 基于种群的龄级结构与径级结构对相同环境具有反应一致性的规律[14],以径级代替龄级划分年龄结构。根据山蜡梅的生长特性,结合实地调查结果,参照灌木、小乔木的径级划分方法[15-17],依据基径(BD)分为8个径级,其中,BD≤1.5 cm为Ⅰ级,1.5 cm
灌木的株高在数量性状中较为稳定,且测量误差小[18],因此,分析植物种群的高度结构不仅可以显示其垂直空间资源竞争情况,也可以作为种群结构稳定性的辅助判断依据。依据株高(H)分为6个等级,从1.0 m起,每增加1.0 m为1个等级,即1.0 m
采用WPS Office EXCEL 2019软件分别绘制山蜡梅种群的天然林(P1至P3)、干扰林(P4至P6)以及整体(P1至P6)的径级和高度级结构图。基于所有山蜡梅植株株高和基径数据,对株高和基径进行Pearson相关性分析,参考张婕等[19]的方法绘制山蜡梅基径和株高散点图,采用SPSS 19.0软件构建回归模型。
1.2.3 种群结构动态量化分析 参考陈晓德[20]的方法对不同样地山蜡梅种群进行动态量化分析。用Vn表示该种群从n到n+1径级的山蜡梅个体数量变化动态,即相邻径级间的动态指数;用Vpi表示不考虑未来外部环境干扰时,种群的数量变化动态指数,即种群动态指数;用Vpi′表示在受到外部干扰时种群的数量变化动态指数,即随机干扰动态指数。Vn、Vpi和Vpi′值的正值、零和负值分别表示2个径级间或整个种群的山蜡梅个体数量的增长、稳定和衰减的动态趋势。用Pmax表示遭受外界干扰时,种群结构对外界干扰的敏感性指数,即随机干扰风险概率。
1.2.4 静态生命表编制及生存分析 分别编制山蜡梅种群的天然林、干扰林和整体的静态生命表,并绘制存活曲线、死亡率曲线和消失率曲线。各参数的计算参考相关文献[6,15,21]中的方法。由于编制特定时间的生命表调查的个体为多个世代的重叠,造成分析结果产生一些负值数据,本研究中的负值为人为干扰的结果。为了分析人为干扰作用的程度,本研究未对实验数据进行匀滑处理,而直接采用实测数据。
参照杨凤祥等[22]的方法,利用生存率函数S(i)、积累死亡率函数F(i)、死亡密度函数f(ti)和危险率函数λ(ti)分析山蜡梅的生存规律,各函数的计算公式参照文献[23-25]。采用指数函数和幂函数检验山蜡梅种群存活曲线的类型[26]。
1.2.5 种群个体数量的时间序列预测 采用时间序列分析中的一次移动平均法[25,27,28]预测山蜡梅种群在天然林和干扰林以及整个种群在未来2、4、6个径级的发展趋势,分析乌溪江流域山蜡梅的种群动态。
2 结果和分析
2.1 山蜡梅种群的结构特征
2.1.1 径级结构特征 供试6个样地山蜡梅种群的径级结构统计结果显示:在调查的6个样地中共发现365株山蜡梅,其中天然林内有234株,干扰林内有131株,以径级为横坐标、个体数量占比为纵坐标绘制径级结构图,结果见图1。

结果表明:天然林内的山蜡梅个体数量在Ⅰ级〔基径(BD)≤1.5 cm〕和Ⅱ级(1.5 cm 2.1.2 株高与基径的相关性 基于株高和基径构建的供试山蜡梅种群的回归模型,结果见图2。经计算,山蜡梅株高与基径的相关系数为0.646,且相关性达到极显著(P<0.01)水平,拟合的一元线性回归方程为y=0.839x+2.069(F=258.673,P=2.684×10-44)。说明山蜡梅的株高与基径呈极显著正相关,不同高度级的个体数量分布情况亦可以反映山蜡梅种群的年龄结构状况。 图2 基于株高和基径的乌溪江流域山蜡梅种群的回归模型 2.1.3 高度级结构特征 供试6个样地山蜡梅种群的高度级结构统计结果(表2)显示:天然林中,H3级〔3.0 m<株高(H)≤4.0 m〕山蜡梅的个体数量最多,共82株,占天然林样地内山蜡梅个体总数的35.0%;H5级(5.0 m 表2 乌溪江流域山蜡梅种群的高度级结构 对乌溪江流域山蜡梅种群的动态量化分析结果见表3。由表3可见:天然林中,山蜡梅各径级的动态指数均大于0,种群动态指数(Vpi)为27.71%,随机干扰动态指数(Vpi′)为1.15%,随机干扰风险概率(Pmax)为0.042,说明天然林中山蜡梅各径级的个体数量呈增长趋势,种群较为稳定,但易受外界干扰影响。干扰林中,V1至V4的动态指数均小于0,而V5至V7的动态指数均大于0,Vpi值为7.00%,Vpi′值为0.11%,Pmax值为0.016,说明干扰林中山蜡梅幼龄个体缺失造成种群内小径级和中径级个体数量呈现衰减趋势,种群增长趋势减弱,趋于稳定,同样易受外界干扰。对山蜡梅种群整体而言,除V4外,其余Vn值均大于0,说明该山蜡梅种群个体在由Ⅳ级〔2.5 cm<基径(BD)≤3.0 cm〕向Ⅴ级(3.0 cm 表3 乌溪江流域山蜡梅种群的动态指数1) 由编制的乌溪江流域山蜡梅种群静态生命表(表4)可见,天然林中,山蜡梅种群各径级的个体数(ax)和标准化个体存活数(lx)均随径级增加逐渐减少;Ⅰ级〔基径(BD)≤1.5 cm〕的生命期望(ex)最高,达到3.493,总体来看,随着径级增加,各径级的ex值呈逐渐下降的趋势。Ⅱ级(1.5 cm 干扰林中,Ⅴ级(3.0 cm 表4 乌溪江流域山蜡梅种群的静态生命表1) 总体来看,随着径级增加,整个种群的ex值逐级降低;从Ⅲ级到Ⅳ级的qx值为负值且最低,Ⅳ级的Kx值也为负值且最低;从Ⅵ级到Ⅶ级的qx值最高,Ⅵ级的Kx值也最高。 天然林山蜡梅种群存活曲线指数函数和幂函数的拟合方程分别为Nx=8.166e-0.088x(R2=0.911,F=61.424,P=0.000)和Nx=7.883x-0.272(R2=0.718,F=12.275,P=0.008);种群整体存活曲线指数函数和幂函数的拟合方程分别为Nx=7.745e-0.046x(R2=0.822,F=27.732,P=0.002)和Nx=7.564x-0.140(R2=0.614,F=9.546,P=0.021)。2种函数拟合结果的P值均小于0.05,达到显著水平,且指数函数的R2和F值均大于幂函数。结合上述方程及存活曲线(图3-A),天然林和种群整体的存活曲线更趋于Deevey-Ⅱ型,且各径级存活个体数量相差较大,因此,天然林和种群整体的存活曲线趋于Deevey-Ⅱ型中的B1亚型;干扰林的存活曲线趋于Deevey-Ⅰ 型,预示干扰林中的山蜡梅种群个体在达到平均生存年龄时将集中死亡。 由图3-B和图3-C所示,天然林和整体的山蜡梅种群的死亡率曲线和消失率曲线的变化趋势一致,均呈双峰型,2个峰值分别出现在Ⅱ级〔1.5 cm<基径(BD)≤2.0 cm〕和Ⅵ级(3.5 cm 由乌溪江流域山蜡梅种群的生存分析结果(表5)可见,天然林中,山蜡梅种群的生存率函数和累积死亡率函数分别呈持续降低和持续升高的趋势,二者此消彼长。其中,生存率函数在Ⅱ级〔1.5 cm<基径(BD)≤2.0 cm〕急剧下降,且随着径级的增长,累积死亡率函数逐渐升高,Ⅵ级(3.5 cm 表5 乌溪江流域山蜡梅种群的生存分析1) 干扰林中,山蜡梅种群的生存率函数和累积死亡率函数变化趋势恰好相反,前者先升高后降低,后者则先降低后升高,且二者均在Ⅳ级(2.5 cm 整体来看,乌溪江流域山蜡梅种群的生存率函数和累积死亡率函数的变化趋势与天然林基本相同,且其生存率函数较同径级天然林升高,累积死亡率和危险率较同径级天然林下降,Ⅰ级(BD≤1.5 cm)至Ⅳ级的死亡密度函数低于同径级天然林,而Ⅴ级至Ⅷ级(BD>4.5 cm)的死亡密度函数高于同径级天然林。 对乌溪江流域山蜡梅种群经过2、4和6个径级时间后各径级的个体数量进行预测,结果见图4。结果显示:经过2、4、6个径级时间后,天然林中山蜡梅种群各径级的个体数量均逐渐增多,平均增幅分别为36.5%、87.4%和65.9%。干扰林中,受山蜡梅种群的Ⅰ级〔基径(BD)≤1.5 cm〕和Ⅱ级(1.5 cm 植物种群的年龄结构是其生物学特性和环境因子共同作用的结果[29,30],能够反映植物种群的动态变化和未来演化趋势[31,32]。调查和分析结果表明:山蜡梅为乌溪江流域天然林中的优势种,其种群结构为增长型;干扰林中山蜡梅的种群结构由增长型转变为稳定型。McEvoy等[33]认为,人为干扰破坏会导致植物种群密度下降,实生苗缺乏。李辛雷等[34]发现,在野外条件下,杜鹃红山茶(CamelliaazaleaWei)具有较高的结实率和产苗潜力,但无序砍伐和采挖等人为干扰不仅使其适宜生境破碎化,而且使其种群中小径级幼苗严重缺乏,导致种群呈衰退趋势。从供试山蜡梅的整体种群结构看,该山蜡梅种群仍属于增长型,说明目前的人为干扰水平对乌溪江流域山蜡梅种群的影响尚未超过其耐受阈值,但是若不积极采取相应保护措施,随着人为干扰程度的加剧,乌溪江流域山蜡梅种群的稳定性势必受到不利影响。 种群结构的动态量化分析能够更加准确地评价种群的结构特征[29]。本研究中,天然林中山蜡梅种群的所有径级个体数量均呈增长趋势,种群较为稳定。干扰林中山蜡梅种群V1至V4的动态指数均为负值,说明干扰林中山蜡梅小径级和中径级个体数量的大量减少,使种群呈现衰退特征。相关研究结果[35-38]表明:高脆弱性是很多濒危植物种群的共同特点,人为干扰和环境竞争致使濒危植物的种子萌发和幼苗生长受到阻碍,威胁其种群稳定性。本研究结果显示:天然林、干扰林和整个种群均易受外界干扰。因此,迫切需要对乌溪江流域的山蜡梅种群及时开展封育保护措施,为其实生幼苗生长创造有利条件,以维持其种群个体数量的稳定性和遗传多样性。 生存函数能够更直观、具体地反映种群增长与衰亡的动态过程[19]。生存函数分析结果显示:天然林中山蜡梅种群呈现前期锐减,中期相对稳定,后期达到生理枯竭年龄后快速衰减的特点;干扰林中,山蜡梅种群生存率和累积死亡率在Ⅳ级〔2.5 cm<基径(BD)≤3.0 cm〕存在明显拐点,表现为幼龄期递增、中龄期递减、老龄期衰退的趋势。 静态生命表能够反映植物种群与生境及其他物种间的适应状态,体现种群的动态趋势,存活曲线、死亡率曲线和消失率曲线则能够揭示植物种群的动态特征,是种群内部各个特征的综合表现[25]。天然林中,山蜡梅种群的死亡率曲线和消失率曲线具有相近的变化趋势,均在Ⅱ级(1.5 cm 研究发现,种子萌发并发育成幼苗阶段受阻是引起植物濒危的主要原因[41]。山蜡梅种子数量较多且萌生性强,其生存林分的郁闭度为0.50~0.75,适宜山蜡梅幼苗实生苗或萌生苗的生长,由此推断山蜡梅种群数量减少与其自身的繁殖能力无关,主要是受外界干扰的影响。人为干扰后,乌溪江流域山蜡梅种群中的大径级个体增幅明显高于小径级个体,这可能是因为小径级个体的定植与生存动态对环境的响应更为敏感,而大径级个体受干扰后相对较难恢复,且山蜡梅的生命周期相对较短,伴随越来越多个体进入老龄阶段,该种群将面临衰退的风险。维持种群稳定的关键是为种子萌发、幼苗定居和生长创造适宜条件,这也是种群成功更新的关键[42],因此,应在山蜡梅种子萌发和幼苗生长期采取必要的人工抚育措施,提高幼苗成活率,这对于维持山蜡梅野生种群的稳定性具有重要作用。 合适的干扰强度和频度可以改善森林生物多样性,调整林分结构,提升森林的整体功能。为促进目的树种抚育经营和野生山蜡梅种群保护工作协调发展,建议采取以下措施对乌溪江流域山蜡梅种群进行保护和管理:1)对山蜡梅种群实施原地保护,并适度疏伐与山蜡梅存在竞争的植物以及山蜡梅的丛生枝或病弱个体,为山蜡梅拓展生存空间,提高其幼苗和幼树的生存期望;2)采取人工播种或补植实生苗到干扰林中的方式补充干扰林中的山蜡梅幼苗和幼树,以维持种群的续存和稳定;3)以乌溪江流域山蜡梅野生种群为资源搜集对象,通过遗传多样性研究,选取核心种质,建立山蜡梅近地种质资源保存库,同时,采取人工培育苗木和促进结实等方法,人为扩大乌溪江流域山蜡梅种群的分布范围。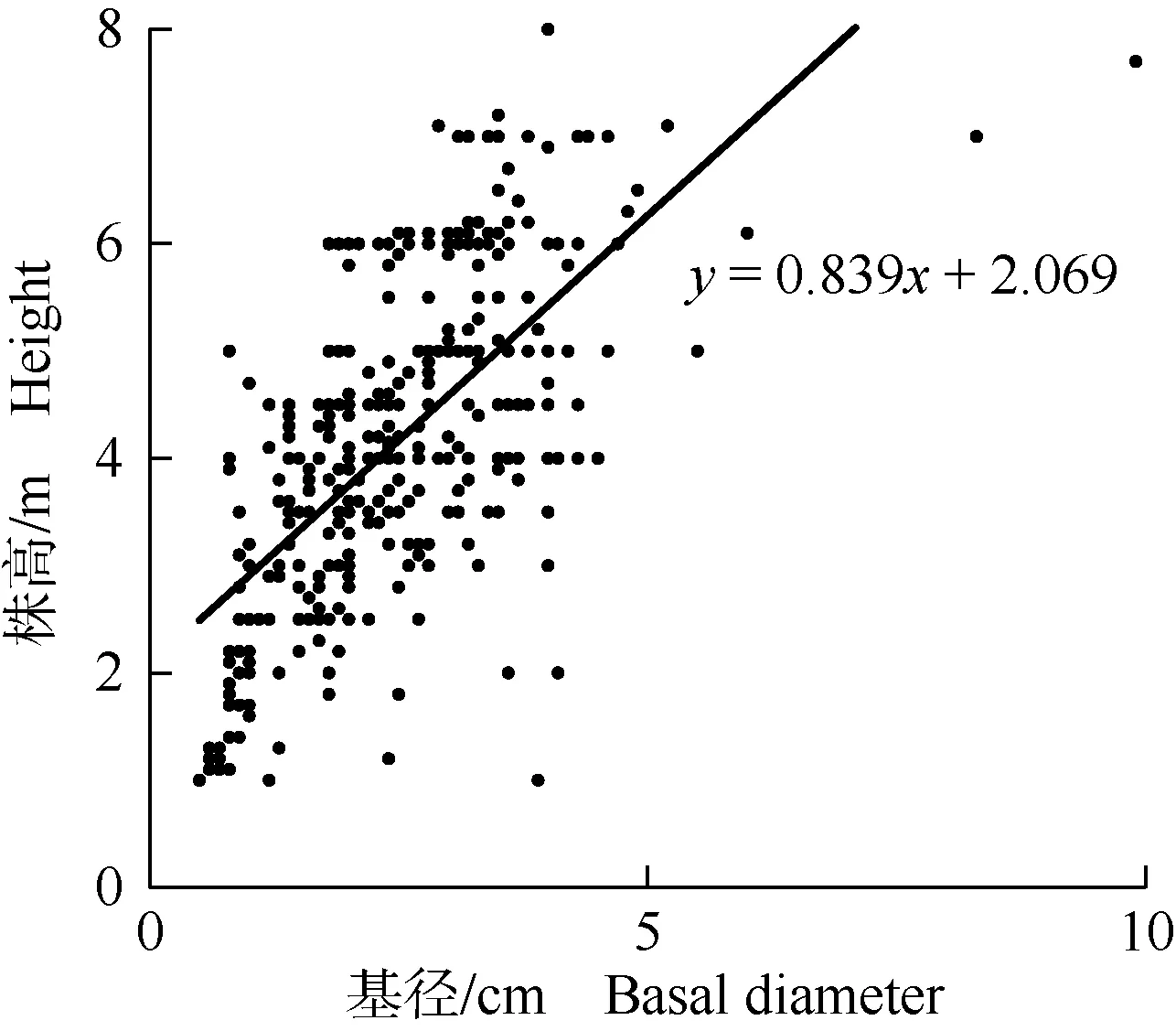

2.2 山蜡梅种群的动态量化分析

2.3 山蜡梅种群的静态生命表分析

2.4 山蜡梅种群的存活曲线、死亡率曲线和消失率曲线分析
2.5 山蜡梅种群的生存分析


2.6 山蜡梅种群的时间序列预测

3 讨 论
3.1 人为干扰对乌溪江流域山蜡梅种群结构的影响
3.2 人为干扰对乌溪江流域山蜡梅种群数量动态变化和稳定性的影响
3.3 乌溪江流域山蜡梅种群的保护管理策略
猜你喜欢
广西林业科学(2022年4期)2022-11-07林业调查规划(2022年3期)2022-06-13科学大众(2021年6期)2021-07-20军工文化(2021年2期)2021-03-30四川林业科技(2019年5期)2019-07-07安徽农学通报(2019年1期)2019-02-14湖北林业科技(2019年6期)2019-01-06启蒙(3-7岁)(2019年1期)2019-01-03湖北林业科技(2017年2期)2017-05-25西北植物学报(2015年9期)2015-07-05
