
视觉发育关键期单眼形觉剥夺对大鼠视功能和视皮层PKCζ蛋白表达的影响△
2022-12-28 13:11路致远毕爱玲毕宏生王兴荣
眼科新进展 2022年12期
李 谦 路致远 毕爱玲 李 洋 毕宏生 王兴荣
弱视是儿童视觉发育期常见的一种眼病,其发病机制较为复杂,与视皮层发育及突触可塑性密切相关。弱视患者会出现双眼视觉异常和视力下降等不同程度视功能损害的表现,其中立体视功能缺失最为常见[1]。立体视觉是指双眼感知周围物体三维空间立体结构的能力[2]。弱视导致的儿童视功能损害已成为严重的社会学问题,因此,深入研究弱视的发病机制对于该病的预防和治疗具有重要意义。
蛋白激酶C(PKC)属于一种磷脂依赖性丝氨酸/苏氨酸激酶,是最早被发现的激酶,主要包括典型或传统PKC、新型PKC和非典型PKC。其中PKMζ是非典型PKC的一种亚型,由PKCζ异构体的独立催化域组成,通过PKCζ基因的内部启动子开启表达[3],PKMζ和PKCζ二者具有相同的结构域和较高的氨基酸同源性,一般认为二者功能相似,在学习巩固、记忆维持及突触可塑性等方面发挥着重要作用[4]。单眼形觉剥夺可通过影响视皮层神经元发育导致弱视的产生,作为一种特异性蛋白激酶,PKCζ是否在视皮层突触可塑性中发挥重要作用,仍需要进一步探讨。本研究分析视觉发育关键期单眼形觉剥夺对大鼠视功能和视皮层PKCζ蛋白表达的影响,为弱视发病机制研究提供新的方向。
1 材料与方法
1.1 实验动物及分组选取出生后13 d SPF级Long Evans大鼠28只,常规检查双眼情况,排除眼部器质性疾病。标准环境下饲养,控制12 h/12 h的昼夜光照规律,温度和湿度适宜,食物水源充足,自由饮食。利用随机数字表法将大鼠随机分为两组,正常对照组、弱视模型组,每组14只,以右眼作为实验眼。本研究中动物处理遵循《实验动物管理条例》(2017修订版)的规定。
1.2 主要试剂及仪器兔抗PKC ζ(美国Santa Cruz Biotechnology公司),兔抗Actin、 N-亚甲基双丙烯酰胺、SDS(美国Sigma公司),蛋白预染Marker(美国Thermo Fisher Scientific公司),Immobilon Western化学发光底物(美国 Millipore 公司);视觉电生理仪(英国 Optoprobe公司),脑切片模具(深圳市瑞沃德生命科技有限公司),荧光显微镜(日本Olympus株式会社),FUSION FX7多功能成像系统(法国Vilber Lourmat公司)等。
1.3 方法
1.3.1 大鼠弱视模型的建立参照参考文献[5-6]中的方法建立多维度弱视模型。在大鼠出生后13 d,腹腔注射50 g·L-1的水合氯醛溶液进行麻醉,剪去距离上下睑缘0.8~1.2 mm处的皮肤和组织,用6-0眼科缝合线对右侧眼睑进行2~4针的间断缝合,建立单眼形觉剥夺模型。若术后出现脱线、漏光、眼内感染等情况,予以弃用,最终纳入实验大鼠共28只,每组14只。两组大鼠均在相同条件下饲养至出生后21 d时剪开眼睑,第51天行闪光视觉诱发电位(F-VEP)检查,记录P2波振幅和潜伏期,验证弱视模型的建立。
1.3.2 F-VEP检查F-VEP检查过程严格参照国际临床视觉电生理学会发布的临床视觉电生理规范化标准,采用全自动视觉电生理系统来完成。具体步骤:将大鼠置于黑暗环境中进行暗适应,暗适应12 h后给予10 g·L-1戊巴比妥钠溶液进行腹腔注射麻醉。将大鼠放置在电生理仪的固定平台上,安装F-VEP检查专用针电极。电极正极位置在动物大脑枕叶的距状裂视中枢,负极位置在动物同侧颊部,接地线位置为动物尾部皮下。用闪烁光作为刺激光,刺激频率1 Hz,通频带宽0.5~85.0 Hz,分析时间250 ms,叠加60次,连续测量至少3次,记录P2波振幅和潜伏期。
1.3.3 视觉悬崖实验检测大鼠立体视功能根据参考文献[7]设计视觉悬崖任务检测大鼠立体视功能。应用摄像头进行视频记录,每只动物观察记录5 min。在下一只动物开始之前,应先清除上一只动物留下的粪便和尿液,然后用体积分数75%酒精擦洗有机玻璃箱底部和周围,避免对下一只动物测试产生影响。在实验结束后应用动物行为追踪软件Tox Trac对大鼠行为视频进行分析,根据分析结果及相关公式计算每只大鼠的辨别指数(discrimination index,DI)[7], DI=(ts-td)/ttot,其中ts和td分别为大鼠探索平台浅侧和深侧所花费的时间,ttot为测试过程的总时间。
1.3.4 HE染色观察大鼠视皮层形态结构F-VEP检测结束后快速断头取脑并放在脑切片模具中,采用前囟标记法,根据大鼠脑立体定位图谱[8]切取左侧视皮层脑组织,将取下的视皮层组织放入40 g·L-1多聚甲醛中进行固定,经过脱水、固定、包埋、切片,再进行脱蜡水化,在脱蜡前置于60 ℃烘烤60 min,将组织切片分别置于二甲苯I和II中浸泡5 min,无水乙醇浸泡5 min,采用体积分数95%、85%、75%乙醇分别浸泡2 min,最后用蒸馏水浸泡2 min。用苏木素染色3 min,水洗5 min;伊红染色30 s,自来水冲洗。脱水透明后,采用中性树胶封片,在光学显微镜下观察并拍照。
1.3.5 Western blot检测大鼠视皮层中PKCζ蛋白的表达本实验选取左侧视皮层(剥夺眼对侧)进行研究,利用Western blot检测PKCζ蛋白在视皮层的表达情况。视皮层取材方法同上,采用组织裂解法提取视皮层蛋白,取上清液用BCA法测定蛋白水平。将蛋白置于100 ℃水浴加热使蛋白变性。进行SDS聚丙烯酰胺凝胶恒压电泳,电泳至嗅酚蓝到胶底部即可停止。恒流转膜后,用50 g·L-1脱脂奶粉溶液封闭孵育。按比例加入PKCζ抗体(1200)、Actin 抗体(11000)置于4 ℃层析冷柜摇床上孵育过夜。室温下进行二抗避光孵育,ECL发光反应后,利用FUSION FX7多功能成像系统拍摄显影照片,用凝胶成像系统Bio-1D 软件对所得条带进行数据处理和分析。
1.4 统计学方法将实验所得的数据采用SPSS 22.0统计软件进行分析,计量数据用均数±标准差来表示。采用独立样本t检验进行组间比较。检验水准:α=0.05。
2 结果
2.1 两组大鼠F-VEP检查结果F-VEP检查结果显示:与正常对照组相比,弱视模型组大鼠右眼(剥夺眼)的P2波潜伏期明显延长,P2波振幅明显降低,差异均有统计学意义(均为P<0.05) 。说明大鼠弱视造模成功(图1和表1)。
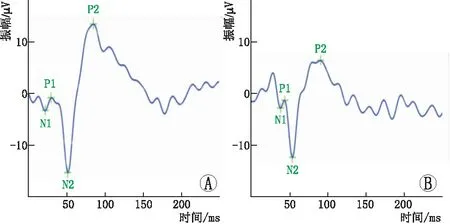
图1 两组大鼠右眼F-VEP检查结果 A:正常对照组;B:弱视模型组。
2.2 两组大鼠视觉悬崖实验检测结果两组大鼠视觉悬崖实验检测结果显示:正常对照组大鼠探索行为轨迹集中在右侧(非悬崖侧);而弱视模型组大鼠探索行为轨迹紊乱,探索区域大多分布在左侧 (悬崖侧)。弱视模型组大鼠DI明显低于正常对照组 (P<0.05) (图2和图3)。
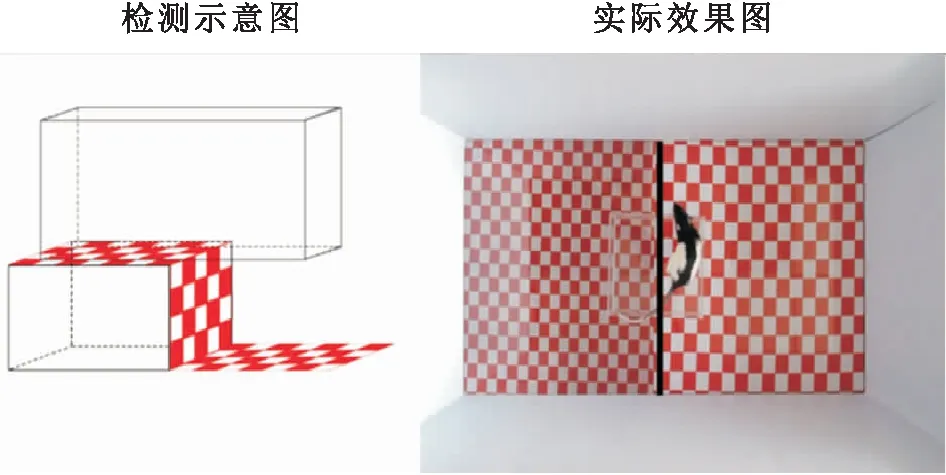
图2 视觉悬崖实验检测示意图及实际效果图 实际效果图:黑色竖线左侧为悬崖侧,右侧为非悬崖侧。

图3 视觉悬崖实验测试行为轨迹图及统计图 与正常对照组相比,*P<0.05。
2.3 两组大鼠视皮层组织HE染色结果HE染色结果显示:正常对照组大鼠视皮层神经元数量丰富,结构完整,细胞包膜清晰,胞核大而圆,核仁明显,未出现明显神经元变性和坏死;弱视模型组大鼠视皮层神经元数量明显减少,结构不完整,细胞包膜不清,胞核变小,出现不同程度的胞体固缩 (图4)。

图4 两组大鼠视皮层组织HE染色结果
2.4 两组大鼠视皮层组织中PKCζ的表达情况Western blot检测结果显示:弱视模型组大鼠左侧视皮层PKCζ蛋白相对表达量明显低于正常对照组(P<0.01)(图5)。
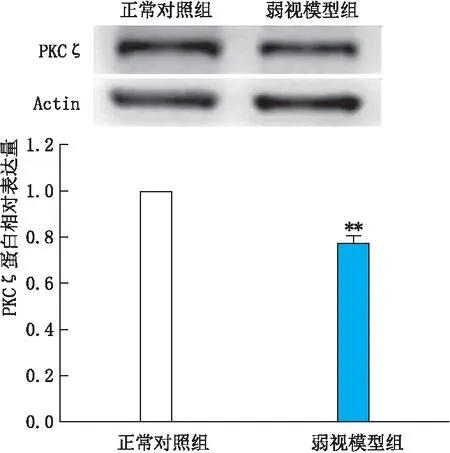
图5 两组大鼠视皮层组织中PKCζ的蛋白表达情况 与正常对照组相比,**P<0.01。
3 讨论
20世纪60年代,有研究人员对6~14个月龄的婴儿进行了一项深度视觉的研究——视觉悬崖实验,该实验后来成为发展心理学的经典研究方法[9]。随着实验技术的进步,视觉悬崖实验逐渐用于测试动物深度视觉能力[10-11]。Russell等[12]通过测量老鼠在平台之间跳跃的能力,对大鼠先天视功能进行相关研究,首次证明了老鼠天生具有深度视觉感知能力。视觉悬崖实验基本原理是利用大鼠在立体视觉正常的情况下具有自发回避危险(视觉悬崖侧)的倾向性来检测研究对象的立体视功能,它已成为评估大鼠立体视功能的一种简单方法[13]。
本实验中我们采用红白棋盘格图案和透明玻璃板营造一种“视觉悬崖”虚拟效果,通过计算DI来评估大鼠的视功能。双眼视功能正常才能区分悬崖侧和非悬崖侧,DI可以从某种程度上反映出双眼视功能情况。结果表明,正常对照组大鼠在探索过程中比较偏好非悬崖侧,回避悬崖侧,说明正常大鼠双眼视觉较好,具有良好的深度视觉感知能力。而弱视模型组大鼠DI较低,而且探索过程中比较偏好悬崖侧,对悬崖深度的感知能力明显下降,表明其双眼视功能较差,深度视觉受到损害。这与前人的研究结果一致,即在视觉发育关键期进行形觉剥夺后,可造成弱视大鼠的视功能严重受损[14]。一方面,单眼形觉剥夺减少了大鼠双眼接收到的光刺激,导致大鼠视皮层神经元兴奋性降低,双眼匹配过程部分中断,引发视皮层功能改变,造成视觉感知能力丧失。另一方面,在视觉发育关键期,视觉环路的形成过程存在着以兴奋性神经递质谷氨酸为代表的兴奋性神经元和以抑制性神经递质γ-氨基丁酸为代表的抑制性神经元,兴奋与抑制神经元的平衡对关键期可塑性的调节发挥着重要的“开关”作用[15-16]。Pettigrew[17]研究视觉经验对小猫视皮层神经元发育的特异性影响发现,视觉经验在小猫视皮层方向选择中发挥着重要作用,证明了视皮层视差选择功能的发展需要视觉经验的刺激,并且视皮层神经元的视差选择功能的改善可能是由皮质内抑制机制的逐渐出现引起的。由此推断单眼形觉剥夺可能引起了视皮层兴奋-抑制神经元平衡系统紊乱,导致视皮层神经元的视差选择功能受损,引起了双眼处理信息能力的缺失,影响了双眼视神经兴奋性的传导,从而造成了立体视觉发育异常。
PKCζ是PKC家族成员之一,具有一定的神经特异性,在大部分皮质、边缘系统和丘脑中广泛表达[18]。有人利用免疫组织化学的方法对猫不同发育阶段的视皮层PKC蛋白的表达情况进行研究发现,视皮层PKC 的表达变化与神经可塑性的发展具有一致性,人们认为PKC可能参与了视觉发育神经可塑性调控过程[19]。非典型PKC (PKCζ和PKMζ)是PKC的一种亚型,在大脑早期发育中发挥作用,与长时程增强诱导过程中突触增强和突触标记有关[20-21]。Castillo等[22]应用免疫组织化学方法对黑暗环境饲养的大鼠视皮层进行研究发现,大鼠视皮层内PKCζ和PKMζ表达减少,由此可推测,单眼形觉剥夺可能也会造成大鼠视皮层PKCζ和PKMζ减少。本实验通过建立多维度弱视大鼠模型,应用Western blot检测大鼠视皮层PKC蛋白表达变化发现:弱视模型组大鼠左侧视皮层PKCζ蛋白相对表达量明显低于正常对照组。PKC蛋白表达水平的下降可能与单眼形觉剥夺后视皮层神经元的发育受损或延迟成熟有关,单眼形觉剥夺后引起视皮层突触神经元传递效率持续性下降或突触传递强度减弱,造成了视皮层突触可塑性的显著变化。
猜你喜欢
中华实验眼科杂志(2022年6期)2022-11-15
小天使·三年级语数英综合(2020年2期)2020-01-13
首都体育学院学报(2019年5期)2019-10-18
今日农业(2019年12期)2019-08-15
读友·少年文学(清雅版)(2019年11期)2019-04-20
中医眼耳鼻喉杂志(2019年3期)2019-04-13
中医眼耳鼻喉杂志(2019年3期)2019-04-13
当代陕西(2018年24期)2019-01-21
中国当代医药(2017年17期)2017-07-25
支部建设(2016年18期)2016-11-28
