
水稻C2H2型锌指蛋白研究进展
2023-01-05 07:22吕朝鹏杨红春
江西农业 2022年22期
吕朝鹏,杨红春
(武汉大学生命科学学院杂交水稻国家重点实验室,湖北 武汉 430072)
锌指蛋白是一类重要的转录因子,具有结合Zn2+的手指状结构,能够调控植物生长发育、参与胁迫应答[1,2]。根据锌指蛋白氨基酸序列中半胱氨酸和组氨酸残基排列顺序和数量的差异,可分为C2H2、C2HC、C3H、C2HC5、C3HC4、C4、C4HC3、C6、C8九种亚型[3]。植物中存在大量的C2H2型锌指蛋白,水稻中至少发现了189个该类蛋白[4]。本文总结了近年来水稻中C2H2型锌指蛋白在水稻生长发育、非生物胁迫中的研究进展,以期对水稻中该类蛋白的研究提供参考。
1 C2H2型锌指蛋白分类与结构特征
首个C2H2型锌指蛋白TFIIIA(transcription factor IIIA)发现于非洲爪蟾的卵母细胞中,能够结合并起始5SrRNA基因的转录[5]。植物中首个被报道的C2H2型锌指蛋白是矮牵牛花瓣中优势表达的EPF1[6],随后在大豆[7]、小麦[8]、拟南芥[9]、水稻[10]中均发现了C2H2型锌指蛋白。
C2H2型锌指蛋白含有保守的CX2-4CX3FX5LX2HX3-5H序列(C代表半胱氨酸、X代表任意氨基酸,H代表组氨酸),这段保守的序列形成1个双链反向平行的β折叠和1个α螺旋,并通过结合锌离子构成稳定的锌指结构[11]。根据α螺旋区域是否存在保守的QALGGH序列,C2H2型锌指蛋白可分为包含该序列的Q型和不含该序列C型[11]。当Q型锌指蛋白中该保守序列突变时,蛋白对DNA的结合能力减弱甚至丧失[12]。此外,根据锌指数量和位置的不同又可将其分为三C2H2型、分散成对排列的C2H2型及多个毗邻的C2H2型[13],如图1所示。
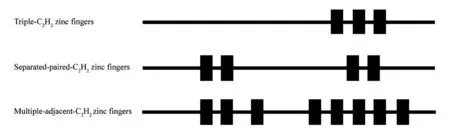
图1 植物C2H2型锌指蛋白结构图
绝大多数C2H2型锌指蛋白除了锌指结构域外,还含有核定位信号(nuclear localization signal,NLS),可能和蛋白互作相关的L-BOX结构域,以及和转录活性相关的EAR结构域(ethylene-responsive element-binding factor(ERF)-associated amphiphilic repression (EAR) domain)[14]。尽管C2H2型锌指蛋白OsDRZ1(Oryza sativa droughtresponsive zinc finger protein 1)和ZFP179都拥有EAR结构域,然而OsDRZ1具有转录抑制活性,ZFP179具有转录激活作用[15,16],说明C2H2型锌指蛋白对转录的抑制或者激活不仅仅取决于EAR结构域。
2 C2H2型锌指蛋白在水稻生长发育中的功能
C2H2型锌指蛋白调控了水稻株高、小穗发育以及开花时间等。近年来水稻中报道的该类蛋白在生长发育中的功能如表1所示。

表1 水稻C2H2型锌指蛋白在水稻生长、发育中的功能
2.1 C2H2型锌指蛋白ZFP207、OsDRZ1调控水稻株高
株高是影响水稻生物量、抗倒伏性、机械收获效果的重要因素。赤霉素(Gibberellin, GA)是影响株高的主要激素,正向调控植物株高[17]。C2H2型锌指蛋白ZFP207、OsDRZ1通过GA调控水稻株高。ZFP207在水稻节间高水平表达,ZFP207过表达植株表现出矮化、根长变短、花粉活性降低等类似GA缺乏的发育缺陷,外源施加GA3能回复过表达ZFP207引起的半矮杆表型,表明ZFP207对水稻的生长抑制和GA信号途径相关。ZFP207结合绿色革命基因SD1/OsGA20ox2(GA20 oxidase)染色质,并抑制其表达。SD1/OsGA20ox2编码参与GA合成的GA20氧化酶,通过调控GA的含量调控株高[18]。ZFP207通过抑制SD1/OsGA20ox2的表达阻碍具有生物活性GA的合成,从而负向调控株高[19,20]。过表达OsDRZ1可以促进水稻生长,RNA-Seq分析表明,OsDRZ1过表达植株中GA受体蛋白基因GID1L2表达上调,该基因的上调可能增强GA信号,促进水稻生长[16]。
2.2 C2H2型锌指蛋白SIP1调控水稻花期
开花时间是一项重要的农艺性状,展示植物对季节和地域的适应情况,影响作物产量。水稻Hd3a(Heading date 3a)、RFT1(RICE FLOWERING LOCUS T 1)分别在短日照、长日照条件下促进开花[21,22]。Ehd1(Early heading date 1)促进Hd3a、RFT1的表达,加速开花[23]。C2H2型锌指蛋白SIP1(SDG723/OsTrx1/OsSET33 Interaction Protein 1) 参与Ehd1-RFT1通路调控开花[1]。在长日照条件下,SIP1突变体中Ehd1、RFT1表达下调,开花延迟。研究表明SIP1能够和组蛋白H3K4甲基转移酶OsTrX1(Oryza sativa Trithorax1)互作,招募OsTrX1结合到Ehd1染色质上,促进基因激活修饰H3K4me3在Ehd1染色质上的沉积,增加Ehd1的表达促进开花。
2.3 C2H2型锌指蛋白LRG1、NSG1、LHS1调控水稻小穗发育
水稻的每个小穗由1朵小花和1对护颖、1对副护颖组成,最终发育成1粒种子,其中小花由6个雄蕊、1个雌蕊、1个外稃、1个内稃、2个浆片共同组成。C2H2型锌指蛋白LRG1(LACKING RUDIMENTARY GLUME 1)、NSG1(NONSTOP GLUMES1)、SL1(STAMENLESS 1)通过调控花器官决定基因的表达,控制小穗发育。LRG1在小穗发育期间在花序中表达增加,突变体中,护颖变长、产生额外的外稃样器官,并且和野生型外稃具有相似的维管束数量;同时花器官决定基因OsMADS1、OsMADS14、OsMADS15、DL(DROOPING LEAF)也在变长的护颖和外稃样器官中异位表达,暗示lrg1中额外产生了具有部分外稃特征的器官。这表明,C2H2型锌指蛋白LRG1抑制护颖到外稃的同源转化,维持小穗的正常发育[24]。此外,NSG1突变体中除了含有正常数量和形状的外稃外,部分护颖、浆片转变为外稃样器官,花器官决定基因LHS1(LEAFY HULL STERILE1)、DL在护颖中异位表达,并且DL也在浆片中异位表达,暗示C2H2型锌指蛋白NSG1抑制护颖、浆片到外稃的同源转化,维持小穗的正常发育。LHS1启动子区存在DBS(DST-binding sequence)元件,NSG1结合该基序,并通过和OsTPRs(TOPLESS-RELATED genes)互作招募组蛋白去乙酰化酶HDACs(histone deacetylases),从而降低LHS1染色质上的乙酰化水平,抑制其表达,调控小穗发育[25]。此外,单锌指C2H2型蛋白突变体SL1中浆片和雄蕊转变为内稃、外稃样器官,且SPW1(SUPERWOMAN1)/OsMADS16在雄蕊原基和浆片原基中的表达消失,而SPW1/OsMADS16是决定雄蕊和浆片形成的关键基因[26],因此,SL1可能通过促进SPW1/OsMADS16转录决定浆片和雄蕊的发育[27]。
2.4 C2H2型锌指蛋白DST、LRG1调控水稻穗及籽粒发育
水稻产量主要由籽粒数、千粒重决定,籽粒数由水稻穗分枝数决定,籽粒大小是影响千粒重的重要因素。水稻C2H2型锌指蛋白DST(DROUGHT AND SALT TOLERANCE)能够通过细胞分裂素(cytokinin, CK)途径调控穗分枝数进而决定籽粒数,影响作物产量[16]。DST半显性等位突变DSTreg1中,CK在茎尖分生组织处积累导致穗分枝增加[28],籽粒数增加[29]。DST能够激活CK氧化酶基因Gn1a/OsCKX2(Grain number 1a/Cytokinin oxidase 2)的表达从而降解具有生物活性的CK。而DSTreg1转录激活功能丧失,导致OsCKX2表达下降,促进CK在SAM中积累,穗分枝数目增加,最终增加籽粒数量。并且将DSTreg1引入到水稻中,水稻的小穗数目增加,籽粒数也增加,暗示了其在农业生产中的潜力。
C2H2型锌指蛋白LRG1不仅调控花发育,也通过调控细胞周期、细胞扩展基因的表达,参与颖壳大小的调控。LRG1突变体中颖壳纵切面的内外层表皮细胞变短,细胞数目减少,导致籽粒变短;横截面的细胞增大,造成颖壳变宽[24]。同时LRG1突变体细胞增殖速度变慢,细胞增殖和扩展相关基因BG1(Big Grain 1)、BG2、GS3(Grain Size 3)表达增加;促进籽粒增大的基因GS2、GS5表达减少;细胞周期相关基因CDKA1(Cyclin-dependent kinase A-1)、CDKA2降低;细胞扩展相关基因EXPA1(expansin-A 1)表达明显减少,但EXPA25表达增加。这些结果表明,C2H2型锌指蛋白LRG1通过正向或者负向调控细胞增殖和扩展相关的基因,控制颖壳的细胞增殖和扩展,从而影响穗粒大小。
3 C2H2型锌指蛋白在水稻胁迫中的功能
盐、干旱、冷等非生物胁迫能够通过渗透胁迫、氧化胁迫和离子毒害等次级作用损伤细胞,抑制植物生长。植物通过合成脯氨酸、可溶性糖、甘氨酸、甜菜碱等渗透调节物质维持细胞渗透平衡,并且利用超氧化物歧化酶(Super Oxide Dimutase,SOD)、过氧化氢酶(Catalase,CAT)、过氧化物酶(peroxidase,POD)、谷胱甘肽过氧化物酶(glutathione peroxidase,GPX)清除过氧化氢(H2O2)、超氧阴离子(O2-)等活性氧(reactive oxygen species,ROS),减少ROS造成的细胞损伤[30]。近年来有关水稻中C2H2型锌指蛋白在胁迫中的功能如表2所示。

表2 水稻C2H2型锌指蛋白在非生物胁迫中的功能
3.1 C2H2型锌指蛋白ZFP179、ZFP213、DST、ZFP252等参与水稻对盐胁迫的应答
盐胁迫是植物面临的主要非生物胁迫之一,高盐抑制植物生长导致粮食减产,严重制约农业生产,威胁粮食安全。过表达C2H2型锌指蛋白基因ZFP179、ZFP213、ZFP252能够通过维持渗透平衡和ROS稳态,提高水稻幼苗在盐胁迫下的存活率[31-34]。盐处理ZFP179过表达植株幼苗时,脯氨酸合成限速酶基因OsP5CS(Pyrroline-5-carboxylate synthetase)、脯氨酸转运蛋白基因OsProT(Proline transporter 1-like)、胁迫相关基因OsDREB2A(Dehydration-Responsive Element-Binding Protein 2A)、OsLea3(Late embryogenesis abundant protein, group 3) 表达水平提高,可溶性糖和脯氨酸含量增加[15,35],暗示ZFP179可能通过促进脯氨酸、可溶性糖合成、转运相关基因的表达,促进细胞内脯氨酸、可溶性糖的积累,维持渗透平衡,提高水稻在盐胁迫下的存活率。盐处理OsZFP213过表达植株,幼苗中OsSOD、OsAPX、OsCAT表达增加,H2O2含量减少,存活率提高[32]。而过表达AtSOD、OsAPX2能够提高植物对盐胁迫的耐受性[36,37],表明C2H2型锌指蛋白OsZFP213可能通过提高ROS的清除能力,减少H2O2的积累从而提高水稻在盐胁迫下的存活率。此外,盐处理DST突变体,茎中Na+的积累减少,幼苗对盐的耐受性增加。该突变体可能通过促使气孔关闭,降低呼吸速率从而减少Na+从根到茎的转运,减少盐离子在水稻中的积累,提高其在盐胁迫下的存活率[38]。
3.2 C2H2型锌指蛋白 ZFP252、ZFP245、OsDRZ1、DST等参与水稻对干旱胁迫的应答
全球人口的增长造成农业用水需求不断增加,然而全球可用的淡水资源日益匮乏,因此干旱对全球粮食安全影响越来越大。植物受到干旱胁迫时,种子萌发、开花受精等过程受到影响,导致作物减产。过表达C2H2型锌指蛋白基因ZFP252、ZFP245、OsDRZ1等能够通过维持渗透平衡、ROS稳态,提高水稻幼苗在干旱胁迫下的存活率。和盐处理ZFP179过表达植株类似,干旱处理ZFP252过表达植株,OsP5CS、OsProT、OsDREB1A、OsLea3表达水平提高,脯氨酸和可溶性糖含量增加,幼苗存活率提高[31]。这暗示C2H2型锌指蛋白ZFP252可能通过促进渗透物质的积累增强植物的抗旱性。干旱处理ZFP245、OsDRZ1过表达的植株,相比于野生型,幼苗中H2O2、O2-积累更少,POD、SOD活性增加,存活率提高[10,16]。RNA-Seq分析表明,与野生型相比,干旱处理下OsDRZ1过表达植物的抗氧化活性相关基因表达水平更高[16],暗示C2H2型锌指蛋白OsDRZ1、ZFP245可能通过维持ROS稳态增强植物的抗旱性。此外,C2H2型锌指蛋白OsDRZ1、DST也能通过调控气孔活动增强植物的抗旱性。干旱处理OsDRZ1过表达植株,气孔关闭程度增加,幼苗相对水含量更高,相对水分丢失速率更低,耐旱性增强[16]。与此不同的是,DST突变体气孔密度减少、关闭程度增加,水稻的抗旱性增强。DST能够通过直接结合到Prx24(Peroxidase 24 precursor)启动子区域的DBS元件,促进Prx24表达,分解控制水稻气孔关闭的H2O2[39]。干旱条件下,DST表达水平降低造成Prx24表达下降和H2O2积累,从而促进气孔关闭,增强水稻的抗旱性[38]。
3.3 C2H2型锌指蛋白ZFP245参与水稻对低温胁迫的应答
在营养生长阶段低温胁迫造成水稻分蘖减少,在生殖生长阶段导致水稻雄性不育,在灌浆期降低水稻灌浆率。C2H2型锌指蛋白ZFP245是水稻中鉴定出的首个同胁迫应答相关的蛋白,低温能诱导其表达。低温处理ZFP245过表达植株,OsP5CS表达增加,自由脯氨酸含量增加,植物存活率明显上升[40];同时SOD、POD含量增加,水稻的抗氧化能力提高[10],这些结果暗示,C2H2型锌指蛋白ZFP245可能通过调控渗透物质的积累和维持ROS稳态增强植物的耐冷性。此外,大豆中鉴定出的C2H2型锌指蛋白SCOF(soybean cold-inducible zinc finger protein)通过和bZIP转录因子SGBF-1互作,促进COR(cold response protein)表达[7],而水稻中的C2H2型锌指蛋白是否能够以及如何调控低温基因应答有待进一步的研究。
3.4 C2H2型锌指蛋白OsART1参与水稻对铝胁迫的应答
铝以Al3+离子的形式溶解在酸性土壤中,通过抑制植物根系生长,影响植物对水分、营养物质的吸收,对植物产生毒害,限制作物产量[41,42]。相比于其他主要农作物,水稻对铝的耐受能力更强。这种耐受同C2H2型锌指蛋白OsART1(Al resistance transcription factor 1)相关[43]。OsART1在根中组成型表达,表达水平不受铝的影响,OsART1能够直接结合GGNVS顺式作用元件[44],激活铝耐受相关下游基因OsNRAT1(Nramp aluminum transporter 1)、OsALS1(ALUMINUM SENSITIVE 1)的表达,增加水稻对铝的耐受能力[43]。OsNRAT1(Nramp aluminum transporter 1)是特异性Al3+转运蛋白,定位于除表皮细胞外的所有根尖细胞的质膜上。敲除OsNRAT1,铝与细胞壁结合增加,水稻对铝的吸收减少,敏感性增强[45]。OsALS1定位于液泡膜上,负责将铝隔离到液泡中,减轻铝对水稻的毒害[46]。这些结果表明,C2H2型锌指蛋白OsART1能够通过调控铝下游基因OsNRAT1、OsART1的表达,吸收环境中的Al3+并转运到液泡中,增强水稻对铝胁迫的耐受性。
4 C2H2型锌指蛋白在水稻ABA信号转导途径中的功能
4.1 C2H2型锌指蛋白 ZFP179、ZFP36、ZFP213等通过ABA依赖途径参与水稻对胁迫的响应
脱落酸(abscisic acid,ABA)在种子成熟、萌发、气孔运动等发育和胁迫应答调控中扮演了重要角色[49]。盐、干旱胁迫能够促进ABA积累。ZFP179、ZFP182启动子上存在ABA响应元件ABRE(ABA-responsive element)[15,34],ABA能够诱导ZFP179、ZFP182表达,这暗示C2H2型锌指蛋白ZFP179、ZFP182可能通过ABA依赖途径发挥作用。ABA处理ZFP179过表达植株,渗透压相关基因OsP5CS、OsLea3、OsProT表达增加,暗示C2H2型锌指蛋白ZFP179可能通过ABA依赖途径维持胁迫条件下的渗透平衡[15],提高水稻在逆境下的存活率;ABA处理ZFP182过表达植株,OsMAPK1、OsMAPK5表达上调,SOD、APX活性增强[47];ABA处理ZFP36过表达植株,OsMAPK表达增加,SOD、APX活性增强,水稻对H2O2的敏感性减弱[48]。MAPK是ABA依赖途径中与抗氧化相关的信使分子[50]。这些结果暗示,C2H2型锌指蛋白ZFP182、ZFP36可能通过参与ABA诱导的抗氧化防御反应提高植物的存活率。
4.2 C2H2型锌指蛋白DST、OsDRZ1、ZFP179通过非ABA依赖途径参与水稻对胁迫的响应
ABA能够调控H2O2积累控制气孔关闭,增强植物对干旱的耐受性[51]。DST能够通过调控H2O2含量提高水稻对干旱的耐受性[38]。经ABA处理后,DST的表达水平没有明显变化,同时野生型和突变体中气孔开放程度也没有明显变化,暗示C2H2型锌指蛋白DST能够以不依赖于ABA的方式增强水稻对干旱的耐受。
DREB/CBF(The dehydration-responsive element binding transcription factors/C-repeat CRT binding transcription factors)能够以非ABA依赖的方式增强植物的抗逆性[52,53]。OsDREB2A是DREB家族的成员能增强水稻对盐和干旱的耐受[54]。ZFP179过表达植株中OsDREB2A表达增加,耐盐性提高,然而ABA处理ZFP179过表达植株,OsDREB2A表达却没有增加,暗示C2H2型锌指蛋白ZFP179也可能通过非ABA依赖的方式调控DREB2A基因的表达从而增强对逆境的耐受。
5 总结与展望
C2H2型锌指蛋白参与了水稻株高、籽粒发育、花期调控等过程,在生长发育中发挥重要功能,同时该类蛋白也参与了对盐、冷、干旱、铝胁迫的应答。虽然OsART1参与了水稻对铝胁迫的耐受调控,但是仍然不清楚C2H2型锌指蛋白是否参与对其他重金属的耐受调控。目前对该类蛋白的研究主要集中在水稻形态和生理变化等方面的描述,对于逆境胁迫的信号转导途径和分子调控网络的研究仍然有待深入。此外,在水稻的生长发育过程中,C2H2型锌指蛋白能够通过改变染色质状态调控花期,然而能否通过改变染色质状态而增强抗逆能力仍然有待探究。
综上,C2H2型锌指蛋白在水稻的生长发育和胁迫的耐受调控中发挥重要作用,其功能和分子机制还有待进一步研究和挖掘。
猜你喜欢
中国农学通报(2022年13期)2022-05-31
理财周刊(2022年4期)2022-04-30
昆明医科大学学报(2022年1期)2022-02-28
现代畜牧科技(2021年4期)2021-12-05
建材发展导向(2021年11期)2021-07-28
林业机械与木工设备(2021年2期)2021-03-11
当代水产(2020年10期)2020-03-17
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
当代水产(2019年8期)2019-10-12
