
经典表观遗传学现象分析
2023-01-10 10:37黄发享
中学生物学 2022年11期
黄发享 渠 源
(华中师范大学第一附属中学 湖北武汉 430060)
生物体中形态结构和功能特性各异的细胞的出现本质上是因为基因的选择性表达。基因在何时表达、在哪些组织器官中表达以及表达量的高低均受到严格的调控,表观遗传调控在其中发挥着重要的作用。与经典遗传相比,表观遗传也能够使基因的表达和生物的表型发生可遗传的改变,但不同的是表观遗传学视域下基因的碱基序列并不发生改变,并且所带来的改变具有不稳定性和可逆性。表观遗传学在解释基因的选择性表达、细胞分化、拓展经典遗传学边界等方面具有重要意义,因此这一领域的研究成为了近年来生命科学研究的热点。
为紧跟生物学发展的步伐、更好地培养学生的生物学学科核心素养,2019年版人教版高中生物学《必修2·遗传与进化》中也加入了表观遗传的概念及相关实例。但是因为表观遗传的概念与经典遗传有很大不同,表观遗传学这个学科也处于蓬勃发展之中,很多现象和问题还亟待解决,因此对表观遗传概念和作用的理解成为了教学中的难点。如何在教学中使学生们明晰表观遗传学中的概念是需要不断探索的问题。下面介绍位置效应花斑、副突变、X染色体失活、基因组印记这四种经典表观遗传学现象的含义和其内在原理。
1 位置效应花斑
1.1 现象的发现及含义
细胞中间期染色质按其形态特征、活性状态和染色性能可分为常染色质和异染色质。与常染色质相比,由于异染色质处于聚缩状态,无法完成正常的基因转录,因此异染色质区表现出了显著的遗传惰性,而位置效应花斑现象的出现正与异染色质的这一特性有关。
位置效应花斑现象最早是由科学家缪勒在用X射线诱导果蝇突变体时发现的。果蝇中最早发现的突变基因是white基因,该基因使果蝇呈现白眼性状,而不是野生型的红眼性状。但是在诱变过程中,缪勒发现有些果蝇的眼色一部分呈现白色,一部分呈现红色,表现为花斑状。后来科学家通过对这种果蝇的多线染色体进行观察发现,这种现象的出现不是因为white基因的碱基序列发生了改变,而是因为果蝇染色体中出现了染色体倒位,染色体断裂的位点一个在染色体臂间的异染色质区,一个邻近white基因(图1)。由于果蝇花斑眼性状的出现是因为white基因位置改变,进而影响了基因的表达,所以这种现象被称为位置效应花斑。

图1 位置效应花斑原理示意图(引自Elgin and Reuter 2013)
1.2 基本原理
位置效应花斑的出现是由于染色体倒位导致white基因的位置发生改变,由原来位于常染色质区域变为靠近异染色质区域,异染色质区域能够将其染色质聚缩的状态扩展至周围染色质区段,进而导致white基因的转录受到抑制,无法正常表达。
多位科学家对异染色质状态的维持以及常染色质转变为异染色质的机制进行了深入研究,目前形成的调控模型如图2所示。正常情况下,常染色质区域具有H3K4me2/3(组蛋白H3第4位赖氨酸二甲基化或三甲基化)、H3K9Ac(组蛋白H3第9位赖氨酸乙酰化)、H3S10(组蛋白H3第10位丝氨酸)磷酸化等修饰,这些修饰能够促进染色质的开放和基因的表达。而当常染色质转变为异染色质时,这些修饰首先会分别被LSD1(一种组蛋白去甲基化酶)、HDAC1(一种组蛋白去乙酰化酶)和磷酸酶除去(图2A)。随后,SU(VAR)3-9和SUV4-20(组蛋白甲基转移酶)会分别使H3K9(组蛋白H3第9位赖氨酸)和H4K20(组蛋白H4第20位赖氨酸)发生甲基化修饰(图2B),这两个位点的修饰会促进染色质聚缩,进而抑制基因表达。正是在这些表观遗传调控蛋白和修饰方式的调控下,常染色质区域转变为异染色质区域,white基因的表达受到抑制,果蝇出现了位置效应花斑现象。
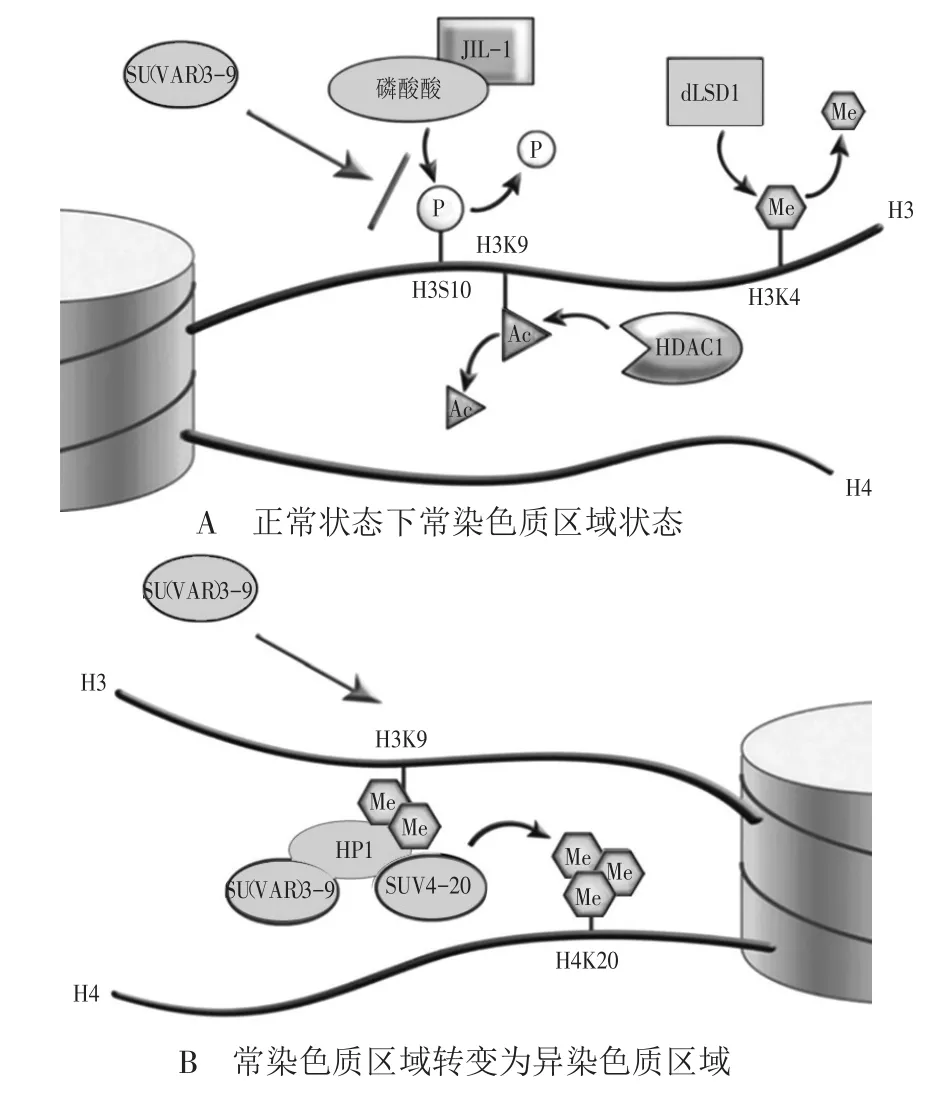
图2 常染色质转变为异染色质的机制(引自Elgin and Reuter 2013)
2 副突变
2.1 现象的发现及含义
副突变最先是由美国威斯康星大学的Brink教授在玉米中发现的。Brink教授发现在玉米R基因的基因座位上存在两个等位基因Rst和Rr,这两个等位基因的纯合个体表现出不同的性状,但是Rst和Rr杂合子自交后代中的Rr纯合子并不表现为Rr所控制的性状,反而表现Rst所控制的性状,但是这一表型不稳定,几代之后又会回复为Rr所控制的性状。由于这种现象不同于突变,但是与突变相似,因此Brink教授将此现象命名为副突变。
后来,科学家在玉米中发现了另一个副突变位点b1。b1基因的表达产物为转录因子,能够激活花青素的生物合成通路,进而使玉米植株呈现出紫色,因此当b1基因的表达发生变化时会造成玉米颜色的改变。在b1基因座位上存在两个特殊的等位基因B-I和B’。研究发现B-I和B’纯合植株分别表现为紫色和绿色,二者杂交产生的子一代表现为绿色,说明B’相对于B-I基因来说为显性。子一代与隐性纯合子进行测交,若符合孟德尔遗传定律,子二代中绿色植株与紫色植株的比例应为1∶1,但实际上子二代植株全部呈现绿色。深入分析发现,B-I和B’基因在碱基序列上并没有区别,启动子区域和编码区的甲基化程度也没有差别。因此,推测可能存在表观遗传调控方式使得B’/B-I杂合子中B-I变为了B’*,进而使子二代中全部表现为B’所控制的性状。
2.2 基本原理
为了进一步分析B-I基因转变为了B’*的内在机制,科学家对B-I和B’基因及其上下游序列进行了分析。结果发现,与正常的b1基因相比,在B-I和B’基因转录起始位点(b1基因座位)上游100 kb处有一个长约6 kb的区域,该区域存在7个853 bp长的串联重复序列(图3)。研究者发现B-I和B’基因中的串联重复序列正义链和反义链都能够转录,并能够形成双链RNA。在B-I和B’基因纯合时,B’基因上游的串联重复序列转录出的双链RNA能够与多种蛋白结合,抑制其自身的表达,因此与B-I基因相比,B’基因的b1基因座位表达量较低,植株呈现绿色。在B’/B-I杂合子中,B’上游串联重复序列转录出的双链RNA能够使B-I转变为B’*,b1基因座位的表达量下降,因此杂合子的测交后代中植株全部表现为绿色。在未来的研究中还需要进一步探讨副突变中RNA介导的转录沉默的机制,明确参与这一过程的关键蛋白。

图3 B-I和B’基因的结构(引自Arteaga-Vazquez and Chandler 2010)
3 X染色体失活
3.1 现象的发现及含义
在XY型性别决定的生物中,雌性个体细胞中含有两条X染色体,雄性个体细胞中含有一条X染色体,若没有相应的调控机制,雌性个体中X连锁基因的表达量将是雄性个体的两倍。但是,实际上雌雄个体中X连锁基因的表达量基本一致。这种现象是通过剂量补偿实现的。剂量补偿有3种方式:①雌性个体每条X染色体上基因的表达量减半,这种机制存在于线虫中;②雄性个体X染色体上基因的表达量是雌性个体每条X染色体上基因表达量的两倍,这种机制存在于果蝇中;③在哺乳动物中,雌性个体的两条染色体中只有其中一条染色体的基因正常表达,这种现象又被称为X染色体失活。
早在1949年,科学家巴尔就在间期细胞核中发现了染色较深、处于聚缩状态的染色质小体,并将其命名为巴氏小体。随后为了解释这一现象,科学家Lyon提出假说:在胚胎发育早期雌性哺乳动物细胞中有一条染色体会失去活性,形成巴氏小体的结构,而且X染色体的失活是随机的。
3.2 基本原理
X染色体失活的直接原因是由于X染色体处于聚缩状态,基因无法正常表达,但是根本原因是非编码RNA Xist的转录以及其带来的X染色体上DNA和组蛋白修饰的变化。在X染色体上有一段序列被称为X染色体失活中心,从X染色体失活中心中会转录出非编码RNA Xist(图4中虚线),Xist能够结合在X染色体上,招募一系列蛋白,使DNA和组蛋白的修饰发生改变(图4中实心圈线),导致染色体聚缩,随后进一步招募其他类型的染色质修饰分子,增加染色质的修饰(图4中空心圆圈),进一步巩固异染色质的结构,使X染色体失活,抑制基因的表达。
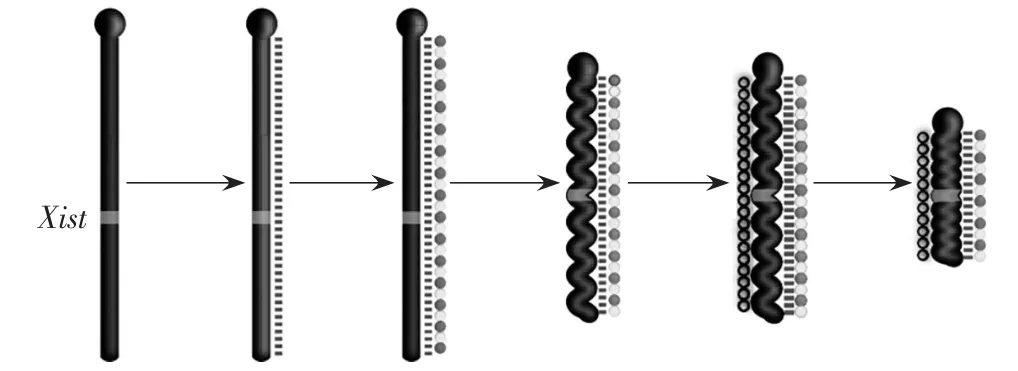
图4 Xist RNA诱导X染色体失活的大致过程(引自Brockdorff and Turner 2015)
4 基因组印记
4.1 现象的发现及含义
对于二倍体哺乳动物来说,体细胞中均含有一半来自父源的染色体和一半来自母源的染色体,因此对于每个基因来说都含有两个拷贝,若父源和母源的基因表达情况相同,那么一个受精卵中仅含有母源的染色体或仅含有父源的染色体仍然可以发育为正常个体。但实际情况并非如此。
有科学家做了这样一个实验:将受精卵中的雌原核或雄原核取出,利用显微注射核移植技术移入一个雄原核或雌原核。结果发现,具有两个雌原核或两个雄原核的胚胎会出现致死现象,只有雄原核和雌原核同时存在的胚胎才能存活。这说明父源和母源染色体上基因的表达并不一致,父源和母源基因的表达对于胚胎的正常发育均至关重要。因此,细胞中可能存在基因组印记,只有父源或母源一方的基因会正常表达,另一方的基因会处于沉默状态。
印记基因的发现证实了基因组印记的存在,研究发现印记基因通常在染色体上成簇存在,例如Igf2r、Igf2、Kcnq1、Gnas、Dlk1基因簇,印记基因簇的基本结构如图5所示,其中ICE为印记控制元件,在父源(Pat)和母源(Mat)染色体中此区域的甲基化程度有显著差别,IG表示印记基因,IG-NC表示印记非编码RNA,NI表示非印记基因。图5中该基因簇为母源表达的印记基因。
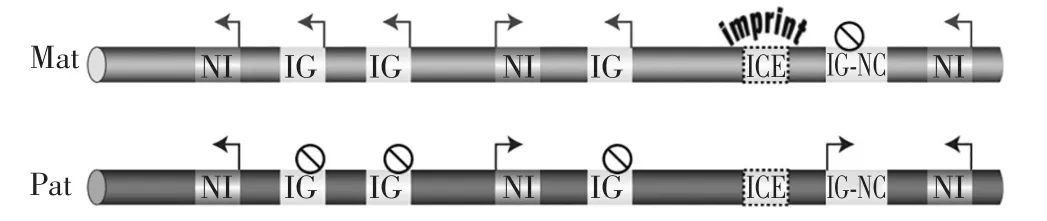
图5 印记基因基本结构(引自Barlow and Bartolomei 2014)
基因组印记与某些人类疾病的发生直接相关。例如,帕德维利综合征和天使综合征的发生均与15号染色体同一片段的缺失有关,父源染色体中这一片段的缺失会导致帕德维利综合征,而母源染色体中这一片段的缺失会造成天使综合征。父源和母源印记基因的缺陷会导致不同类型疾病的发生,在个体中产生不同的表型。
4.2 基本原理
目前基因组印记存在绝缘子模型和长链非编码RNA模型(图6)两种调控机制。
Igf2基因簇为父源表达的印记基因,该印记基因的建立符合绝缘子模型(图6A)。在这一基因簇中,母源染色体的ICE区域不发生甲基化,CTCF蛋白能够结合在ICE区域并组成绝缘子结构,由于绝缘子的存在,增强子(图6中标有E的椭圆)只能激活非编码RNA H19-NC的转录,Igf2和Ins2基因的转录受到抑制。而在父源染色体中ICE区域会发生甲基化,因而无法与CTCF蛋白结合形成绝缘子结构,H19-NC也无法转录,因此增强子能够作用于Igf2和Ins2基因,激活基因表达。
Igf2r基因簇为母源表达的印记基因,该印记基因的建立符合长链非编码RNA(lncRNA)模型(图6B)。在这一基因簇中,母源染色体的ICE区段会发生甲基化,该区段含有非编码RNA Airn的启动子序列,因此ICE的甲基化会抑制Airn的转录,Airn不能发挥转录抑制作用,因而Igf2r等基因得以正常表达。而在父源染色体中,ICE区域不会发生甲基化,因而非编码RNA Airn正常转录,进而抑制了Igf2r等基因的表达。

图6 基因组印记的基本原理(引自Barlow and Bartolomei 2014)
猜你喜欢
畜牧兽医学报(2022年3期)2022-03-30
中国畜牧兽医(2022年1期)2022-02-15
畜牧兽医科技信息(2021年5期)2021-03-05
现代泌尿外科杂志(2019年10期)2019-10-31
生物学通报(2019年2期)2019-06-15
作文周刊·小学二年级版(2019年12期)2019-04-26
畜牧兽医科学(2018年12期)2018-02-18
畜牧兽医科学(2018年20期)2018-02-15
畜牧兽医科技信息(2018年5期)2018-02-13
