
不同倍性紫果西番莲“满天星”形态、叶绿素及荧光参数差异比较
2023-01-12 06:02王雪朱雅静陈诗杨振欣罗茜许玉兰王德新蔡年辉
山东农业大学学报(自然科学版) 2022年5期
王雪,朱雅静,陈诗,杨振欣,罗茜,许玉兰,王德新*,蔡年辉*
不同倍性紫果西番莲“满天星”形态、叶绿素及荧光参数差异比较
王雪1,2,朱雅静1,2,陈诗1,2,杨振欣1,2,罗茜1,2,许玉兰1,2,王德新1,2*,蔡年辉1,2*
1. 西南林业大学 西南山地森林资源保育与利用教育部重点实验室, 云南 昆明 650224 2. 西南林业大学西南地区生物多样性保育国家林业局重点实验室, 云南 昆明 650224
为了探寻两个倍性植株的形态、叶绿素及叶绿素荧光参数的差异,揭示不同倍性光合能力的变异规律,本研究以二倍体和三倍体的紫果西番莲“满天星”()的苗木叶片为材料,测定植株形态及叶绿素含量,用高速连续激发式荧光仪Handy PEA测定其叶绿素荧光参数及叶绿素荧光诱导动力学曲线。结果表明,三倍体植株表现出矮壮、叶柄更长、叶片更宽的现象,三倍体较二倍体叶绿素a、叶绿素b、叶绿素总含量、类胡萝卜素含量大,三倍体对环境的光照辐射适应性较好、不易受到光抑制,光合速率更高。在电子传递过程中,三倍体比二倍体变化波动小,量子比率和热耗散的量子比率方面的分配比较稳定,PSⅡ结构的能量传递较稳定,适应性更强。在不同倍性植株中,二倍体和三倍体的OJIP一致,三倍体的荧光强度明显高于二倍体,说明三倍体的荧光产量更高,接受光量子的能量更好,因此光合碳代谢速率更高。
紫果西番莲; 生物学; 叶绿素; 荧光
紫果西番莲()是西番莲科(Passifloraceae)西番莲属()的多年生常绿藤本攀援植物,其具有较高的食用性、观赏性、药用性,是近年来研究较多的藤本植物[1]。我国主要栽培的品种有“黑美人”、“台农1号”、“黄金杂交种1号”、“满天星”、“芭乐味黄金果”等[2]。近年来西番莲种植业的生产经济效益大幅度增加、栽植区域面积等也都迅速和扩大,但因为我国西番莲产业发展得较晚,种质资源仍相对缺乏,品质保持优良、抗性尚好的优良品种就尤其缺乏。“满天星”是紫果西番莲中的一个品种,具有较厚的紫红色果皮及比较大的果实,适合加工利用[3]。
叶绿体中的色素包含叶绿素和类胡萝卜素,其中叶绿素又包括叶绿素a和叶绿素b,他们的作用分别是转化光能和收集光能,在逆境中植物通过提高光能的转化效率来保证碳的同化,因而叶绿素的含量常被用作评判植物抗性强弱的指标。在太子参多倍体诱导及离体快繁的研究中发现,四倍体太子参的叶绿素含量较二倍体提高了11.11%[4]。对平陆百合试管苗的诱导鉴定中发现,四倍体平陆百合叶片和鳞茎中的叶绿素a、叶绿素b、总叶绿素含量均高于二倍体脱毒试管苗[5]。在对滇杨多倍体的研究中也有同样的结论,滇杨四倍体植株的叶绿素含量是二倍体植株的1.5倍[6]。
叶绿体荧光参数通过叶绿素荧光参数的变化特性来间接反映出光合作用周期中PSⅡ系统对叶绿素光能量的快速吸收、传递、耗散、分配能力等特点[7],于是叶绿素荧光参数同时也常常被拿来作为用来研究多倍体植物优良性、抗性的一个优良参数。在对甜瓜属异源多倍体的研究中发现与二倍体种相比,异源四倍体的PSⅡ电子传递效率(ΦPSⅡ)和光化学淬灭系数(qP)较二倍体种显著升高[8]。杜琳等[9]在对白桦四倍体与二倍体的光合特性比较中也得到了相同的论点,即四倍体白桦叶绿素荧光参数中的PSⅡ中心非环式电子传递量子产额较二倍体提高了13.28%,显著高于二倍体。四倍体与二倍体木薯相比叶绿素荧光参数与叶绿素均升高,四倍体木薯叶片的PSII反应中心捕光能力增强、光化学转化效率升高,参与代谢、光合作用等途径相关蛋白质表达水平上调,叶片的光合速率高于二倍体植株[10]。叶绿素荧光参数已被用作探寻植物光合过程的可靠指标,使我们能更准确的了解植物的光合作用过程。叶绿素荧光参数的差异主要是由于品种之间差异所致,因此,可以通过测定不同品种的叶片荧光参数的差异,作为辅助选育和鉴定优良玉米和棉花品种的重要指标[11]。
目前关于紫果西番莲“满天星”的报道主要集中在种质资源性状评价、多倍体诱导[12]等方面,尚无“满天星”及其多倍体的叶绿素及叶绿素荧光参数方面的报道,本研究以紫果西番莲“满天星”二倍体、三倍体植株为研究材料,通过对其部分形态指标和叶绿素及叶绿素荧光的测定,探寻二倍体、三倍体植株的差异,揭示不同倍性光合能力的遗传变异规律,为其后续生理生化研究及高效栽培技术提供理论依据。
1 材料及方法
1.1 材料
试验以二倍体(2=2=18)、三倍体(2=3=27)紫果西番莲“满天星”苗木为研究对象,试验材料由西南林业大学林木遗传与繁育重点实验室提供,试验在西南林业大学苗圃中进行。
1.2 方法
自2021年7月中开始对5月移栽的苗木生长量进行测量,采用皮尺和直尺(cm)及游标卡尺(mm),精确至0.01,测量两种倍性紫果西番莲的苗高、地径、叶片宽、叶柄长,每种倍性各测定3株苗木,测定至12月止。其中叶片及叶柄长选取植株的中上部成熟叶进行测量。
叶绿素含量测定使用乙醇法,称取剪碎的新鲜样品0.1 g,放入研钵中,加入少量石英砂及少量的95%乙醇,研成均浆,静置后将溶液倒入加了滤纸的25 mL定容瓶中,用95%乙醇的乙醇冲洗定容。把叶绿体色素提取液倒入光径1 cm的比色杯内。以95%乙醇为空白,在分光光度计下分别以波长663 nm、645 nm下或470 nm测定吸光度。测定从2021年7月至12月止。
参照朱景乐等[13]叶绿素荧光参数测量方法。使用高速连续激发式荧光仪Handy PEA对两种倍性植株的叶片进行测定,先将植株叶片利用叶夹夹住暗适应处理后使用机器上的探头进行测量。于2021年7月至12月选取生长较一致的两种倍性植株的顶端下数第3片长势稳定的叶片进行测定,采用完全随机区组处理,3个重复。
PSⅡ原初光化学效率和初始荧光参数测定。叶片暗处理30 min后测定初始荧光(F)、最大荧光(F)、可变荧光(F)、PSⅡ最大光化学效率(F/F)、PSⅡ电子传递速率(F/F)、PSⅡ潜在活性(F/F)、单位反应中心吸收的能量(/)、单位反应中心耗散的能量(/)、单位反应中心捕获的用于还原QA的能量(/)、单位反应中心捕获的用于电子传递的能量(/)、单位面积反应中心的数量(/)、电子传递的量子产额(Φ)、原初光化学反应的最大光化学效率(Φ)、捕获的激子能导致电子传递的比率(Φ)。按照Genty B等[14]、高玉等[15]、夏建荣等[16]的计算方式计算:
单位反应中心吸收的能量:/=M/V/(F/F)。
单位反应中心耗散的能量:/=/-/。
单位反应中心捕获的用于还原QA的能量:/=M/V。
单位面积反应中心的数量:/=(F/F)×(V/M)×F。
电子传递的量子产额:Φ=[1-(F/F)×(1-V)]。
原初光化学反应的最大光化学效率:Φ=/。
捕获的激子能导致电子传递的比率:Φ=1-V。
1.3 数据分析
使用SPSS1625.0和EXCEL 2010软件进数据处理和分析。
2 结果与分析
2.1 二倍体和三倍体紫果西番莲“满天星”形态学分析
如表1所示,随着时间的增加两种倍性的‘满天星’的苗高与地径均呈现增长的状况,且增长的幅度相似,8月至9月有较大幅度的增长,在苗高方面二倍体均显著高于三倍体,在地径方面从9月份开始,三倍体均高于二倍体。两种倍性‘满天星’的叶柄长度随月份的变化呈增长趋势,且从9月份开始三倍体显著大于二倍体,在叶片宽度方面,两种倍性‘满天星’整体体现出先增大后减小的趋势,在10-12月出现增减反复的情况,但整体三倍体的叶片宽度大于二倍体。总体而言,三倍体相比于二倍体的紫果西番莲‘满天星’呈现矮壮、叶柄更长、叶片更宽的的现象。
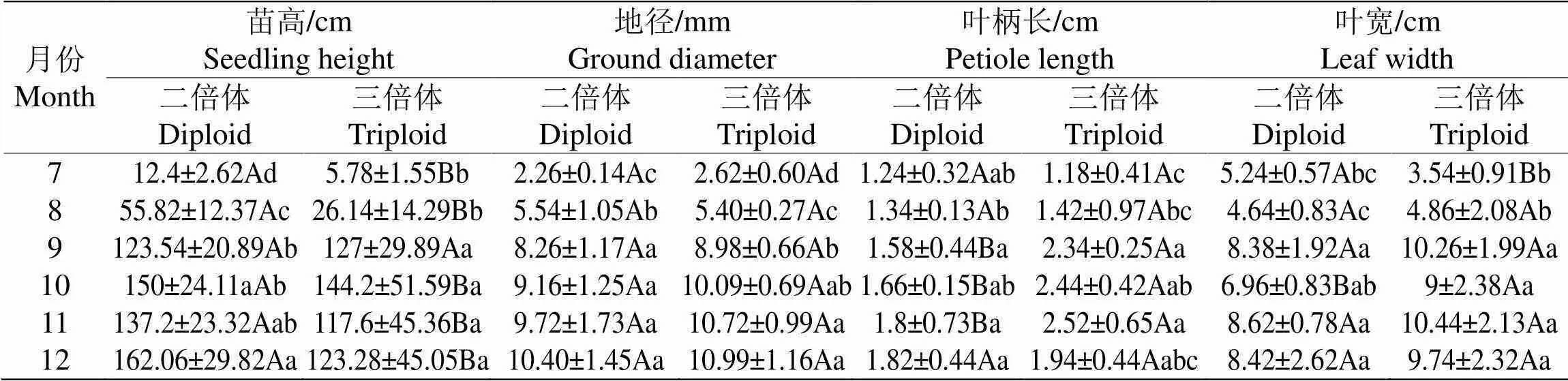
表1 二倍体和三倍体紫果西番莲‘满天星’形态学差异
注:不同大写字母表示各月份中两种倍性‘满天星’在<0.05水平下差异显著,不同小写字母表示各倍性‘满天星’在月份间<0.05水平下差异显著,下同。
Note: Different capital letters indicate that there is a significant difference between the two ploidy 'Mantianxing' in each month at the level of<0.05, and different small letters indicate that there is a significant difference between the ploidy 'Mantianxing' in each month at the level of<0.05, the same below.
2.2 二倍体和三倍体紫果西番莲“满天星”叶绿素含量差异
叶绿素含量的高低常被用来判断植物光吸收能力的强弱,在7-12月对田间生长的两种倍性‘满天星’的叶绿素含量进行测量,结果如表2所示,两种倍性‘满天星’的叶绿素a含量自7月份起持续降低,保持一致的趋势,三倍体‘满天星’的叶绿素a含量始终高于二倍体,说明三倍体‘满天星’吸收光能的能力更强。叶绿素b也有相似的结果,三倍体‘满天星’的叶绿素b含量始终高于二倍体,但与叶绿素a含量的变化趋势不同,两种倍性‘满天星’的叶绿素b含量呈先下降又上升的趋势,在11月两种倍性‘满天星’的叶绿素b含量最低,且在12月时有较大幅度的增加,12月时两种倍性‘满天星’叶绿素b含量较其他月份都要高,这可能是由于环境的变化引起的变化。总叶绿素含量是叶绿素a与叶绿素b含量的总和,三倍体‘满天星’的总叶绿素含量高于二倍体,符合大部分多倍体的特性。两种倍性‘满天星’的类胡萝卜素含量均呈逐渐下降的趋势,在8-11月时三倍体‘满天星’的类胡萝卜素含量都大于二倍体,但在12月时三倍体类胡萝卜素含量大幅降低,三倍体‘满天星’的类胡萝卜素含量仅在12月份低于二倍体。
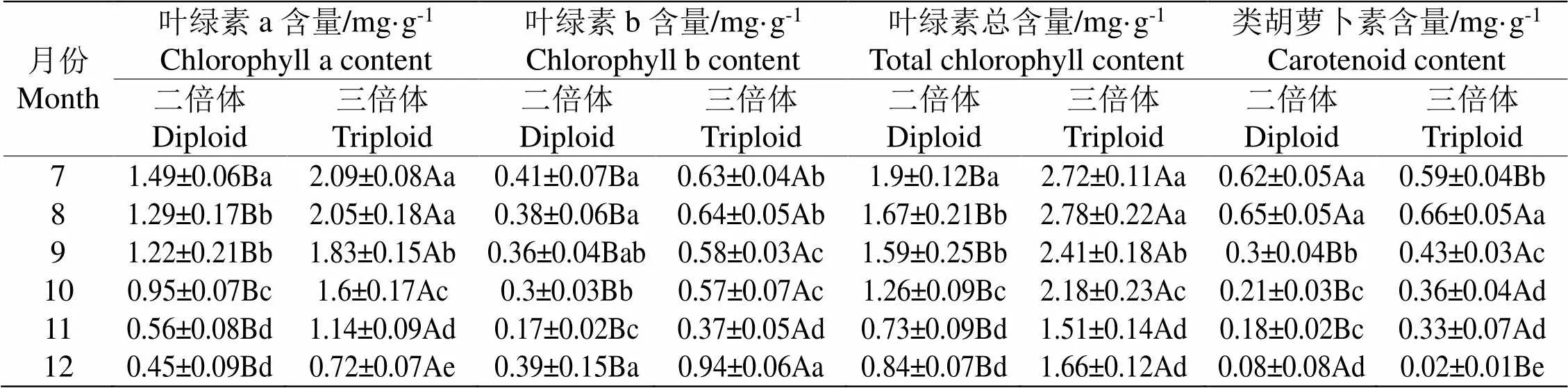
表2 二倍体和三倍体紫果西番莲‘满天星’叶绿素含量差异
2.3 二倍体和三倍体紫果西番莲“满天星”PSⅡ原初光化学效率和初始荧光参数
叶绿素荧光参数在测量光合作用时植物需先经过光暗处理,它的实际上意义是指植物光合作用状态从休止性到活化性状态间的变化,测量可以揭示整个植物在光合作用中产生的荧光动态的变化,因而认为它可能包含了某些重要的生物光合信息。从图1-a可以看出,三倍体的初始荧光F随着月份的变化较小,二倍体初始荧光F则呈现增减反复的趋势,分别在8月和11月达到峰值并高于三倍体。图1-b、图1-c可知,二倍体的最大荧光F和可变荧光F变化趋势基本一致,都随着时间变化呈现先升高后降低的趋势;三倍体的最大荧光F和可变荧光F变化趋势基本一致,在10月前都保持不显著的变化,10月后开始显著降低;三倍体的F、F都大于二倍体,并在7、9、11、12月都显著大于二倍体,且二倍体的下降趋势更显著。图1-d、图1-e、图1-f可知,两种倍性的“满天星”PSⅡ最大光化学效率(F/F)、PSⅡ电子传递速率(F/F)、PSⅡ潜在活性(F/F)的变化趋势基本一致,10月前变化波动较小,10月后显著降低,并在9、11、12月显著大于二倍体,二倍体的下降趋势也更显著。
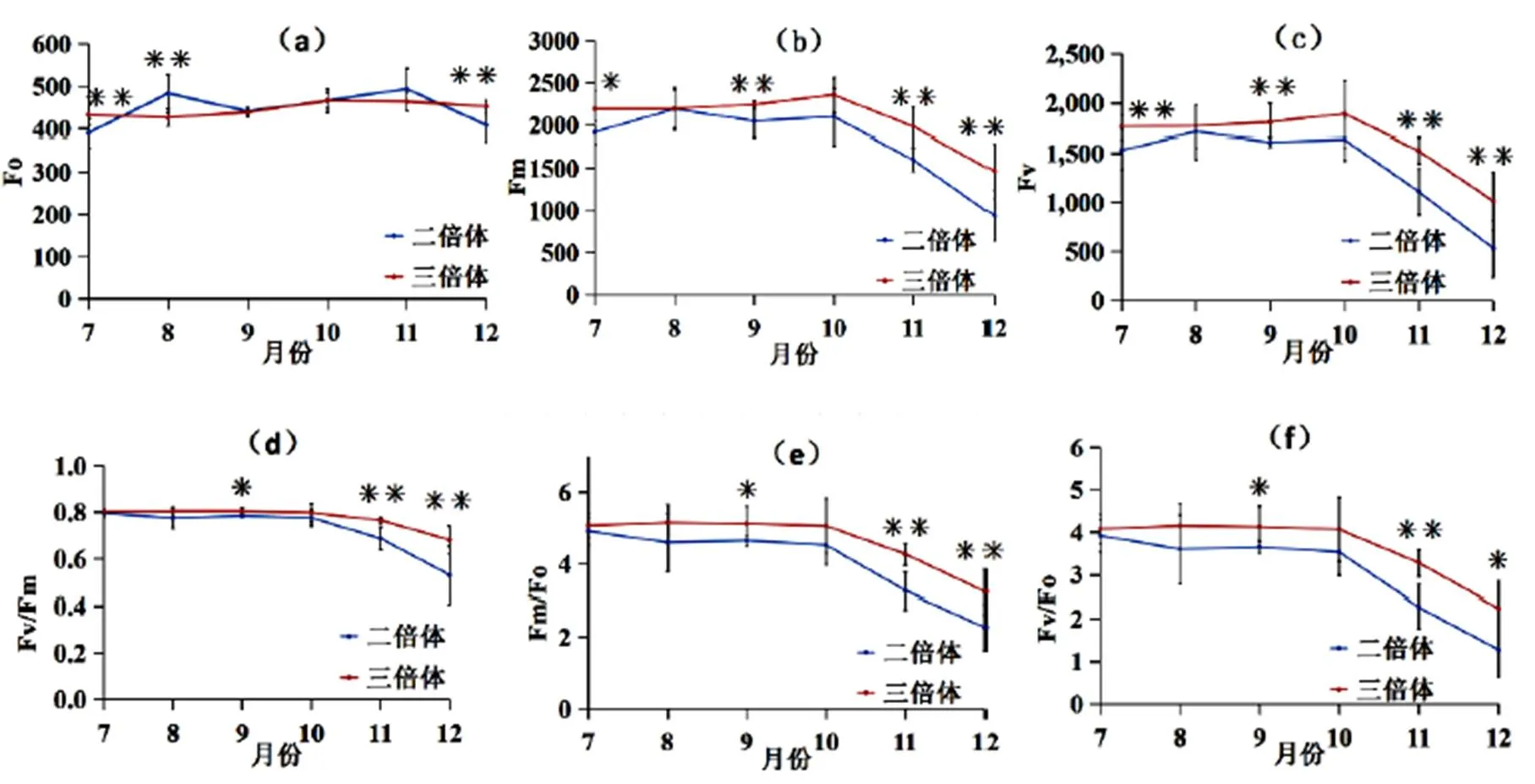
图1 二、三倍体紫果西番莲“满天星”PSⅡ原初光化学效率和初始荧光参数差异图
2.4 二倍体和三倍体紫果西番莲“满天星”PSⅡ反应中心能量流动分配
从图2-a可知,在两种倍性“满天星”中,单位反应中心吸收的能量(/)均呈现先下降后上升的趋势,在10月降到最低,在12月达到最大;二倍体“满天星”的/在7~12月均大于三倍体,且9~12月显著大于三倍体;二倍体“满天星”的/在10~12月波动变化大于三倍体。从图2-a可知,两种倍性的“满天星”单位反应中心耗散的能量(/)变化趋势相同,10月前无显著变化,10月后呈上升趋势;二倍体的/均大于三倍体,且9~12月显著大于三倍体;二倍体“满天星”的/在10~12月波动变化较三倍体大。从图2-c、图2-d可知,两种倍性的“满天星”单位反应中心捕获的用于还原QA的能量(/)和单位反应中心捕获的用于电子传递的能量(/)变化趋势基本一致,且二倍体的月变化显著大于二倍体。
从图2-e可知,两种倍性的“满天星”单位面积反应中心的数量(/)的月变化相同,先增加后降低,三倍体的/的月变化显著大于二倍体。从图2-f可知,在不同倍性“满天星”中,电子传递的量子产额(ΦE)均呈现10月无显著变化,10月后显著下降,11~12月二倍体的ΦE的变化量显著大于三倍体。从图2-g、图2-h可知,两种倍性的原初光化学反应的最大光化学效率(ΦP)、捕获的激子能导致电子传递的比率(Φ)的月变化趋势基本一致,呈现反复增减的趋势并在9月达到最小,三倍体的整体变化较二倍体平稳,二倍体波动较大。
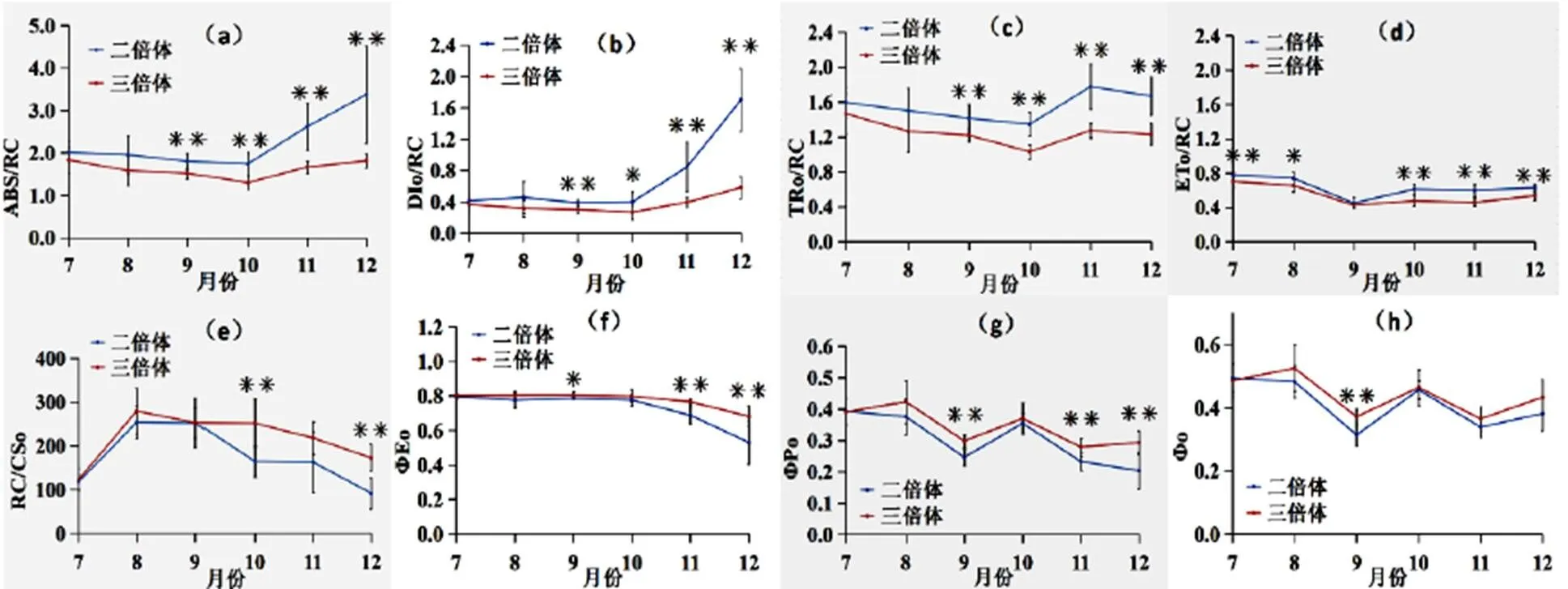
图2 二倍体和三倍体紫果西番莲“满天星”PSⅡ反应中心能量流动分配的月变化
2.5 二倍体和三倍体紫果西番莲“满天星”叶绿素荧光诱导动力学曲线
当绿色植物暴露于在强可见光照射时,含丰富叶绿素基质的绿色组织上会诱导发出强度在不断产生改变的红色荧光,荧光强度也随暴露时间长短的而变化,这种由不同波长变化的强度而形成的动力学曲线即称为植物叶绿素荧光强度诱导动力学曲线[17]。一般情况下,刚暴露在光下的初始荧光定义为O点,荧光最大值定义为P点,叶绿素荧光诱导动力学曲线就是反映O点到P点的荧光变化过程。从图3可以看出,不同倍性的“满天星”的测定曲线趋势基本一致,荧光产量随着时间增加均有所上升,其中OJIP曲线的所有点(O、J、I和P)均有所增加;三倍体OJIP曲线所有点(O、J、I和P)都大于二倍体,在O点、J点、I点和P点荧光值均表现为三倍体>二倍体。

图3 二倍体和三倍体紫果西番莲“满天星”叶绿素荧光诱导动力学曲线
注:(a)为线性时间坐标(b)为对数时间坐标。
Note: (a) linear time coordinate. (b)logarithmic time coordinate.
2.6 二倍体和三倍体紫果西番莲“满天星”各项指标相关性分析
将测得的4个形态指标、叶绿素含量与PSⅡ原初光化学效率和初始荧光参数进行了相关性系数分析。初始荧光(F)与最大荧光(F)呈正相关,与叶绿素a、叶绿素b、叶绿素总含量、类胡萝卜素呈负相关,但仅与叶绿素总含量显著相关性。最大荧光(F)、可变荧光(F)、PSⅡ最大光化学效率(F/F)、PSⅡ电子传递速率(F/F)、PSⅡ潜在活性(F/F)与叶绿素a、叶绿素b、叶绿素总含量、类胡萝卜素呈正相关,且除叶绿素b以外均为极显著关系,与叶柄长、叶宽呈极显著负相关。叶绿素a与叶绿素b、叶绿素总含量、类胡萝卜素均呈极显著正相关关系,相关系数分别为0.348、0.956、0.809,与叶柄长呈显著负相关,相关系数为0.333。叶绿素b与叶绿素总量、叶绿素总含量与类胡萝卜素呈极显著正相关关系。叶柄长与除初始荧光(F)、叶绿素b、地径、苗高以外的指标均呈显著性负相关关系。苗高与地径、叶宽、叶柄长均呈显著正相关关系。
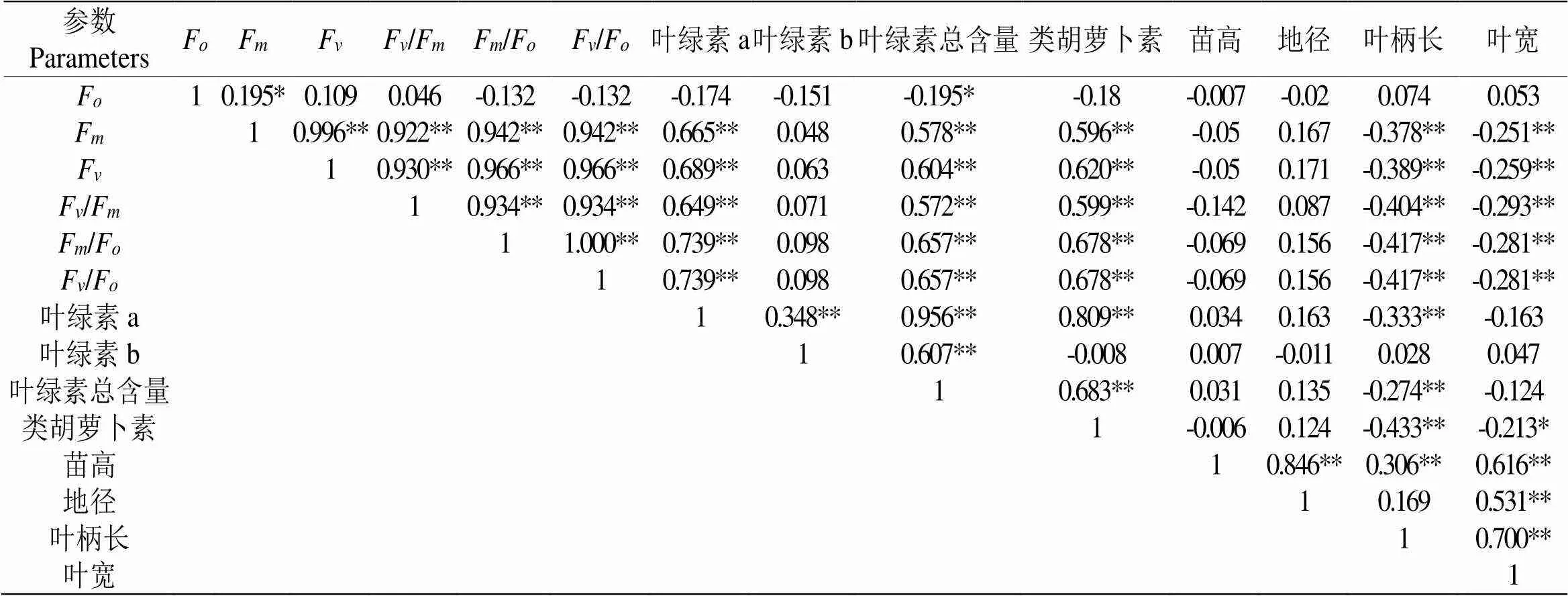
表3 二倍体和三倍体紫果西番莲“满天星”各项指标相关性分析
注:*在0.05级别(双尾),相关性显著,**在0.01级别(双尾),相关性显著。
Note: *At 0.05 level (double tail), the correlation is significant, * * at 0.01 level (double tail), the correlation is significant.
3 讨论
在植物的生长发育过程中,资源的分配策略随时间和环境的变动而改变,植物因资源分配不均导致植株两种不同性状的相对生长速率产生差异,这是一种异速生长现象[18],即植株在自身的两个部分分配不同的资源去生长,树高与地径是最常用于对比植物表型变化的生长指标,能够直接反映苗木的生长状况。对二倍体和三倍体‘满天星’的生长情况进行观测,发现二倍体‘满天星’的苗高大于三倍体,地径小于三倍体,这与蒋卉等[19]的研究结果相似,研究表明多倍体植物通常具有更粗壮的茎[20],如在对象鼻兰的诱导试验中发现,四倍体象鼻兰较二倍体粗壮[21]。本研究中三倍体‘满天星’也拥有较粗壮的茎。在对多倍体枇杷的叶部形态研究中发现,三、四倍体枇杷的叶长叶宽叶柄均大于二倍体枇杷[22],这与本研究相似。
叶绿素含量常被看做能反映植物光合作用强弱的重要指标,在对两种倍性‘满天星’叶绿素含量测定的中,三倍体‘满天星’的叶绿素含量始终高于二倍体,表明三倍体比二倍体的吸光能力更强。研究表明,在相同环境中不同植物叶绿素荧光特性不同[23],相同植物在不同光照强度下产生的叶绿素荧光特性也不同[24],相同植物不同倍性的叶绿素荧光特性也不同[10]。叶绿素荧光技术通过光照强度对PSⅡ结构和功能的影响来反映植物的光合能力[25,26],进而反映出不同植物、不同强度、不同倍性的适应性。
初始荧光F是PSII反应中心完全开放时的荧光产额,它与叶片叶绿素浓度有关。二倍体F的月变化增减反复,说明二倍体对于环境的光照辐射适应性差,在8月和11月达到峰值并高于三倍体,说明在8月和11月植物以热的形式和荧光的形式散失能量,类囊体膜受损,植物的光合作用受损;三倍体“满天星”F月变化较小,说明三倍体对环境的光照辐射适应性较好。最大荧光F是PSII反应中心完全关闭时的荧光输出,它能反映电子通过PSII的转移,随着月份增加温度的降低,Fm和F呈下降趋势,所以初始F、F数值水平较高,且下降率较低为适应性较好的植株,三倍体的初始F、F大于二倍体,且三倍体下降率较低,说明三倍体‘满天星’PSII反应中心处于完全关闭时的电子传递情况较二倍体好,三倍体较二倍体对环境的适应性强。孙国荣等[27]研究发现高光合速率通常伴随着较高PSⅡ最大光化学效率(F/F)、PSⅡ电子传递速率(F/F)、PSⅡ潜在活性(F/F),也可以反映外界环境对PSⅡ的影响[28,29]。结果表明,二、三倍体“满天星”的F/F、F/F、F/F均在12月时最低,说明在12月温度最低时受到的光抑制最强。且二倍体的F/F、F/F、F/F始终小于三倍体植株,说明二倍体植株更易受到光抑制[30],三倍体植株的光合速率更高。
PSⅡ中单位反应中心光能的转换和通过电子传递情况可以通过/、/、/、/、/、ΦE、ΦP和Φ的变化反应。二倍体的/、/、/、/的月变化幅度大于三倍体,且二倍体的值始终大于三倍体,说明二倍体PSⅡ结构更易受到温度和辐射的影响,能量大多以热量和荧光的形式散失,以减少温度和辐射对PSⅡ系统反应中心造成的伤害[31]。PSⅡ中的能量分配比率与/、ΦE、ΦP和Φ荧光参数有关[32],两种倍性植株的/、ΦE随着月份增加呈下降趋势,且二倍体下降的趋势大于三倍体,说明二倍体的光合作用中能量分配受到的影响更大,二、三倍体ΦP和Φ呈现增减反复的趋势,且三倍体关系,则而四倍体的的ΦP和Φ波动变化小于二倍体,说明,PSⅡ结构的能量传递较稳定,在电子传递过程中,量子比率和热耗散的量子比率方面的分配比较稳定,适应性更强。
快速叶绿素荧光诱导动力学曲线以生物膜流动为基础,通过氧化还原反应和PSⅡ中电子的传递来反映PSⅡ的供体侧、受体侧及PSⅡ反应中心对荧光的响应[33]。在不同倍性“满天星”中,三倍体的荧光强度明显高于二倍体,说明三倍体的荧光产量较高,接受光量子的能量较好,因此光合碳代谢速率最好。在不同倍性植株中,二倍体和三倍体的叶绿素荧光动力学曲线一致,三倍体的OJIP曲线所有点(O、J、I和P)都明显大于二倍体,说明三倍体的荧光产量较高,接受光量子、电子从QA向QB的传递量较高。
叶绿素荧光参数是鉴定植株光合特性的一个很好的指标,叶绿素荧光和叶绿素含量的关系,前人研究中表明F与叶绿素含量呈正相关[34],本研究也有相同的结论,在史宝胜等[35]的研究中也表现出叶绿素含量越高的品种,光化学效率就越高。本研究叶绿素荧光参数与叶绿素含量、叶柄长、叶宽呈现极显著相关关系,说明各项指标较大的三倍体“满天星”的光合效率较二倍体植株好,说明三倍体比二倍体植株优良。
4 结论
(1)紫果西番莲“满天星”的三倍体较二倍体的呈现矮壮、叶柄更长、叶片更厚的现象;
(2)紫果西番莲“满天星”的三倍体的叶绿素a、叶绿素b、叶绿素总含量、类胡萝卜素较二倍体含量更高;
(3)紫果西番莲“满天星”的三倍体对环境的光照辐射适应性较好、不易受到光抑制,光合速率更高。
[1] 丰国蕊,赵李姗,朱雅静,等.紫果西番莲根尖染色体制片技术研究[J].山西农业大学学报(自然科学版),2021,41(3):42-49
[2] 刘文静,潘葳,吴建鸿.5种百香果品系间氨基酸组成比较及评价分析[J].食品工业科技,2019,40(24):237-241
[3] 章希娟,许玲,魏秀清,等.5份西番莲种质资源性状评价[J].东南园艺,2017,5(6):5-8
[4] 洪晓云.太子参离体快繁及多倍体诱导的研究[D].福州:福建农林大学,2013
[5] 王鹏丽.平陆百合多倍体的诱导与鉴定[D].太谷:山西农业大学,2018
[6] 陈杰,周军,孙正海,等.组织培养结合秋水仙素诱导滇杨多倍体的研究[J].云南农业大学学报(自然科学),2013,28(2):251-256
[7] Rohacek K, Bartak M. Technique of the modulated chlorophyll fluorescence: Basic concepts, usefu1 parameters, and some applicadons [J]. Photosynthetlea, 1999,37:339-363
[8] 陈友根,陈劲枫,李为观,等.甜瓜属不同倍性异源多倍体光合特性及超微结构研究[J].南京农业大学学报,2010,33(1):32-36
[9] 杜琳,李永存,穆怀志,等.四倍体与二倍体白桦的光合特性比较[J].东北林业大学学报,2011,39(2):1-4
[10] 安飞飞,凡杰,李庚虎,等.华南8号木薯及其四倍体诱导株系叶片蛋白质组及叶绿素荧光差异分析[J].中国农业科 学,2013,46(19):3978-3987
[11] 徐洪文,宋凤斌,朱先灿,等.不同生育时期玉米苞叶叶绿素荧光特性差异分析[J].华北农学报,2009,24(6):74-77
[12] Antoniazzi CA, Faria R, Carvalho P,. In vitro regeneration of triploid plants from mature endosperm culture of commercial passionfruit (Sims ) [J]. Scientia Horticulturae, 2018,238:408-415
[13] 朱景乐,杨超伟,杜红岩,等.2个杜仲无性系叶绿素荧光参数对比[J].河南农业大学学报,2014,48(5):575-578
[14] Genty B, Briantais JM, Baker NR. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence [J]. Biochimica et Biophysica Acta General Subjects, 1989,990:87-92
[15] 高玉,高志奎,张晓慧,等.通过快速荧光动力学曲线探测白黄瓜光系统Ⅱ的热激胁迫效应[J].生态学报,2009,29(6):3335-3341
[16] 夏建荣,邹定辉.利用OJIP叶绿素a荧光评估干出对石莼()光系统Ⅱ的影响[J].海洋通报,2007(4):50-55
[17] 李鹏民,高辉远,Strasser RJ.快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J].植物生理与分子生物学 学报,2005(6):559-566
[18] McConnaughay KDM, Coleman JS. Biomass allocation in plants:ontogeny or optimality?A test along three resource geadients [J]. Ecology, 1999,80(8): 2581-2593
[19] 蒋卉,袁欣,冯乃馨,等.矮牵牛同源多倍体诱导及其初期表型差异分析[J].河南农业科学,2019,48(9):111-116
[20] 郭启高,宋明,梁国鲁.植物多倍体诱导育种研究进展[J].生物学通报,2000(2):8-10
[21] 吴婷,葛红,杨树华,等.秋水仙素诱导象鼻兰种子产生多倍体[J].北方园艺,2022(2):71-78
[22] 温国,孙皓浦,党江波,等.多倍体与二倍体枇杷叶片特征及抗旱性初步分析[J].果树学报,2019,36(8):968-979
[23] 李西文,陈士林.遮荫下高原濒危药用植物川贝母()光合作用和叶绿素荧光特征[J].生态学 报,2008(7):3438-3446
[24] 敖金成,苏文华,张光飞,等.不同光强下对马耳蕨叶绿素荧光参数的日变化[J].南京林业大学学报(自然科学 版),2011,35(1):135-138
[25] Baker NR. A possible role for photosystem Ⅱ in environmental perturbations of photosynthesis [J]. Physiol, Plant, 1991,81:563-570
[26] 宋春雨,刘晓冰,金彩霞.高温胁迫下光合器官受损及其适应机理[J].农业系统科学与综合研究,2002(4):252-256
[27] 孙国荣,刘文芳,肖翊华.杂交水稻灌浆期光合生理特性的研究[J].武汉植物学研究,1990(4):341-347
[28] 李磊,李向义,林丽莎,等.两种生境条件下6种牧草叶绿素含量及荧光参数的比较[J].植物生态学报,2011,35(6):672-680
[29] 张静,张元明.冻融过程对生物结皮中齿肋赤藓叶绿素荧光特性的影响[J].中国沙漠,2011,31(6):1479-1487
[30] 姚春娟,郭圣茂,陈兰兰,等.草决明叶绿素荧光特性的秋季日变化[J].农业与技术,2015,35(20):27-28
[31] 杨建伟,赵丹,孙桂芳,等.干旱胁迫对小紫珠光合生理的影响[J].西北植物学报,2018,38(4):733-740
[32] 贾浩,郝建博,曹洪波,等.遮荫对‘保佳红’桃树叶片快速叶绿素荧光诱导动力学曲线的影响[J].西北植物学报,2015,35(9):1861-1867
[33] Maxwell K, Johnson GN. Chlorophyll fluorescence-a practical guide [J]. Journal of Experimental Botany, 2000,51:659-668
[34] 张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999(4):444-448
[35] 史宝胜,徐继忠,马宝焜,等.SH系砧木光合作用特性的研究[J].河北农业大学学报,2001(4):54-57
Comparison of Morphology, Chlorophyll and Chlorophyll Fluorescence Parameters ofwith Different Ploidy
WANG Xue1,2, ZHU Ya-jing1,2, CHEN Shi1,2, YANG Zheng-xin1,2, LUO Xi1,2, XU Yu-lan1,2, WANG De-xin1,2*, CAI Nian-hui1,2*
1.650224,2.650224,
In order to explore the difference of morphology, chlorophyll and chlorophyll fluorescence parameters between two ploidy plants and reveal the variation rule of different ploidy photosynthetic capacity, this study used the seedling leaves of two ploidy"Mantianxing" as materials to determine the plant morphology and chlorophyll content, and measured the chlorophyll fluorescence parameters and chlorophyll fluorescence induction kinetics curve with the high-speed continuous excitation fluorescence meter Handy PEA. The results showed that triploid plants were short and strong, with longer petioles and wider leaves. Compared with diploid plants, triploid plants had more chlorophyll a, chlorophyll b, total chlorophyll content and carotenoid content. Triploid plants had better adaptability to light and radiation, were not easily inhibited by light, and had higher photosynthetic rate. In the process of electron transfer, triploid has less fluctuation than diploid, the distribution of quantum ratio and heat dissipation quantum ratio is relatively stable, and the energy transfer of PS Ⅱ structure is relatively stable and more adaptive. In different ploidy plants, the OJIP of diploid and triploid is the same, and the fluorescence intensity of triploid is significantly higher than that of diploid, indicating that triploid has higher fluorescence yield, better energy to receive light quanta, and therefore higher photosynthetic carbon metabolism rate.
; biology; Chlorophyll; fluorescence
S667.9
A
1000-2324(2022)05-0755-08
2022-05-10
2022-06-27
云南省万人计划青年拔尖人才项目(YNWR-QNBJ-2019-075);云南省研究生导师团队建设项目(2022-97);西南林业大学第一批预研基金(01108-18200133)
王雪(1997-),女,在读硕士研究生,主要研究方向林木遗传育种. E-mail:1334091850@qq.com
通讯作者:Author for correspondence. E-mail:woshixin2007@126.com; cainianhui@sohu.com
猜你喜欢
热带作物学报(2022年5期)2022-06-01
科学养鱼(2021年11期)2022-01-11
山西农业科学(2021年3期)2021-12-06
热带农业科技(2020年3期)2020-12-15
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
农民致富之友(2020年6期)2020-04-08
食品与生活(2019年12期)2019-05-15
世界热带农业信息(2018年11期)2018-01-17
中国果业信息(2015年12期)2015-01-24
