
多糖类成分抗胰腺癌作用机制的研究进展 Δ
2023-01-14 07:20刘光宇王琦苏玲吉林农业大学食药用菌教育部工程研究中心长春10118吉林农业大学中药材学院长春10118吉林农业大学植物保护学院长春10118
中国药房 2023年1期
刘光宇 ,王琦 ,苏玲 (1.吉林农业大学食药用菌教育部工程研究中心,长春 10118;2.吉林农业大学中药材学院,长春 10118;.吉林农业大学植物保护学院,长春 10118)
胰腺癌作为一种发病率逐年上升的恶性肿瘤,隐匿性强,一般在病程进入晚期时方才确诊,因此治愈率低,在全球癌症病死率中排名第7位,患者5年生存率为9%[1]。在我国,胰腺癌的病死率位居癌症病死率第6位,且患者5年生存率低于癌症患者平均水平,仅为7.2%[2]。由于胰腺癌隐匿性强且转移性高,80%~85%的患者在确诊时已经失去手术机会[3]。化疗是转移性胰腺癌的主要治疗手段,但常见的化疗药物存在心、肝、肾毒性和过敏反应等副作用,且耐药性会使疗效降低。近年来,天然产物及其衍生物以低毒、高效的特性受到广泛关注,逐渐成为治疗癌症的潜在药物。
多糖是一类高分子聚合物,通常由10个及以上的单糖基通过α(1,4)、α(1,2)、β(1,3)和β(1,6)等糖苷键聚合、脱水形成,分子量分布跨度较大,几千至数百万均有存在,其作为药用活性成分普遍存在于各种天然产物中。目前,已有100余种多糖被证实具有抑制肿瘤细胞生长的活性[4],这些多糖对机体无副作用,且与化疗药物联用可减缓后者所致的机体免疫抑制效应。近年来,国内外学者已陆续开展多项多糖类成分抗胰腺癌的相关研究。基于此,本课题组通过整理国内外相关文献,系统归纳与总结多糖类成分抗胰腺癌的作用机制,以期为抗胰腺癌药物的研发提供新思路与新方向。
1 多糖类成分抗胰腺癌的作用机制
1.1 诱导胰腺癌细胞凋亡
在癌症发生的过程中,正常细胞转化为癌细胞,而癌细胞可以通过调节多种细胞因子表达来逃避细胞凋亡。抗凋亡蛋白B细胞淋巴瘤2(B-cell lymphoma-2,Bcl-2)与促凋亡蛋白Bcl-2相关X蛋白(Bcl-2-associated X protein,Bax)在许多肿瘤中均呈高表达状态;胱天蛋白酶(Caspases)与抑癌基因p53也是凋亡信号传导通路中的重要成员。远志多糖RP02-1可通过调节Caspase-3、Bax和Bcl-2表达诱导胰腺癌细胞AsPC-1和BxPC-3凋亡[5]。云芝多糖PSK可通过调控Bax和细胞周期蛋白依赖性激酶抑制蛋白p21WAF/Cip1,诱导胰腺癌细胞PANC-1凋亡[6]。牛樟芝多糖可通过线粒体途径促进胰腺癌细胞BxPC-3凋亡,同时改变Bcl-2家族蛋白的表达情况,促进细胞色素C释放,激活Caspase-9、Caspase-3和多聚腺苷二磷酸核糖聚合酶(poly ADP-ribose polymerase,PARP)的蛋白水解,诱导染色质凝聚和DNA片段化[7]。茯苓多糖在干预胰腺癌细胞PANC-1后,可使细胞中的Caspase-3、Caspase-9、p53蛋白水平升高,Bcl-2蛋白水平降低,诱导细胞发生变圆缩窄、膜起泡、细胞融合减少、细胞核浓缩而碎裂等典型的凋亡行为[8]。喜树果实多糖可显著增加胰腺癌细胞BxPC-3的收缩百分比,诱导染色质浓缩、碎片化及凋亡小体的产生[9]。麦冬多糖ROH05的乙酰化物能显著抑制胰腺癌细胞BxPC-3和PANC-1增殖,诱导细胞核破碎、皱缩等细胞凋亡现象的发生[10]。此外,香菇多糖与吉西他滨联用可增强对胰腺癌细胞AsPC-1的抑制作用,下调凋亡抑制蛋白survivin的表达水平,其联合用药效果优于吉西他滨单药治疗[11]。
核因子κB(nuclear factor-κB,NF-κB)是炎症和癌症的关键调节因子,也是赋予胰腺癌细胞凋亡抗性的关键因素之一[12]。研究表明,多糖会干扰NF-κB介导的信号通路,在肿瘤细胞凋亡中发挥关键作用。黑枸杞多糖LRP1-S2在体外和体内对胰腺癌细胞AsPC-1、BxPC-3和PANC-1均有抑制作用,其可通过抑制p38蛋白及丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)/NF-κB 信号通路活性来促进BxPC-3细胞凋亡,进而发挥抗胰腺癌活性[13]。海胆多糖SEP在体内和体外均可通过与Toll样受体4(Toll-like receptor 4,TLR4)结合并上调细胞外调节蛋白激酶(extracellular regulated protein kinases,ERK)、c-Jun氨基末端激酶(c-Jun N-terminal kinase,JNK)、p38蛋白表达和NF-κB磷酸化来激活自然杀伤(natural killer,NK)细胞,从而有效抑制胰腺癌细胞SW1990和BxPC-3的生长[14]。褐藻中分离出的岩藻多糖可激活Caspases依赖的内源性和外源性凋亡途径,抑制NF-κB信号通路,促进肿瘤抑制因子p53的表达,从而通过靶向p53/NF-κB通路,诱导胰腺癌细胞PANC-1凋亡[15]。
由上文可知,多糖类成分主要通过调控Caspases、细胞凋亡因子Bax、Bcl-2以及NF-κB信号通路发挥诱导胰腺癌细胞凋亡的作用。
1.2 诱导胰腺癌细胞自噬
在癌症发展的各个时期,正常的肿瘤细胞自噬可视为肿瘤抑制,一旦自噬减少或发生异常就会导致氧化应激的发生,促进癌症发展。由此,自噬可分为4种主要形式,即细胞保护性(细胞存活)、细胞毒性(细胞死亡)、细胞抑制性(生长停滞)和非保护性(对细胞死亡或存活没有贡献)[16]。
在蒲公英根部中多糖成分的作用下,胰腺癌细胞BxPC-3、PANC-1的线粒体膜电位明显降低、活性氧含量增加,进而在死亡前发生自噬[17]。黄芪多糖可通过上调胰腺癌细胞AsPC-1的自噬特异性标志物MAP1LC3(LC3)的表达诱导癌细胞自噬[18]。芦荟多糖可使胰腺癌细胞BxPC-3自噬启动蛋白1抗体(unc-51 like autophagy activating kinase 1,ULK1)的表达上调,苄氯素1(Beclin-1)、Bcl-2的表达下调,诱导肿瘤细胞产生自噬体或自噬溶酶体,从而发生自噬,进而发挥抗癌效果[19]。远志多糖RP02-1可通过干扰Beclin-1、自噬蛋白5(autophagy related 5,Atg5)和LC3B来抑制胰腺癌细胞BxPC-3自噬,从而抑制其生长[5]。
由上文可知,自噬可发生在癌症代谢过程的多个组成部分中,多糖类成分能通过调节自噬因子,从不同角度靶向自噬及自噬相关途径,从而达到治疗胰腺癌的目的。
1.3 抑制胰腺癌细胞增殖
癌细胞的快速增殖是癌症迅速恶化的原因之一,因此对癌细胞增殖的调控可有效抑制癌症的发展。研究发现,刺参黏多糖可以改善转录共激活因子相关蛋白(Yes-associated protein,YAP)的磷酸化和细胞核-细胞质易位并抑制下游基因转录,通过调控Hippo-YAP通路来抑制胰腺癌细胞SW1990增殖[20]。杭菊花、怀菊花和亳菊花中分离出的6种中性均一多糖均对胰腺癌细胞PANC-1具有明显的增殖抑制作用,其中杭菊花多糖CMTA0S3和怀菊花多糖 CMJA0S2对胰腺癌细胞的抑制率最高(接近70%)且呈现出良好的浓度依赖性,其作用机制可能与调控NF-κB通路有关[21]。三七多糖可以通过直接靶向半乳糖凝集素3(galectin-3,Gal-3)、表皮生长因子受体(epidermal growth factor receptor,EGFR)、骨形态发生蛋白受体(bone morphogenetic protein receptors,BMPRs)并阻断EGFR/ERK/矮小相关转录因子1(runt-related transcription factor 1,Runx1)、BMPR/母亲DPP同源物(mothers against decapentaplegic homologue,SMAD)/DNA结合抑制因子3(inhibitor of DNA binding 3,Id-3)、整合素(integrin)/黏着斑激酶(focal adhesion kinase,FAK)/JNK信号通路而有效抑制胰腺癌细胞BxPC-3的生长[22]。此外,亮菌多糖ATPS可显著增加吉西他滨对胰腺癌细胞BxPC-3、SW1990的增殖抑制作用,其作用机制可能与调控自杀相关因子(factor associated sui-cide,Fas)/Fas-L介导的死亡受体通路和Bax/Bcl-2相关的线粒体通路有关[23]。姬松茸多糖结合硒可以协同改善炎症反应和脂代谢情况,抑制白细胞介素1(interleukin-1,IL-1)、IL-6及肿瘤坏死因子 α(tumor necrosis factor-α,TNF-α)的表达,从而减缓肿瘤生长[24]。更多的细胞实验结果显示,天麻多糖、北极海参多糖、忍冬多糖均对胰腺癌细胞PANC-1具有抑制作用[25];玉竹多糖也可增强顺铂对胰腺癌细胞PANC-1的抑制作用[26];泥螺多糖因其具有较强的抗氧化活性,从而表现出对胰腺癌细胞SW1990的显著抑制作用[27]。
由上文可知,多糖类成分能通过靶向EGFR/ERK/Runx1、BMPR/SMAD/Id-3等信号通路,且多以浓度依赖形式抑制胰腺癌细胞增殖。
1.4 调控胰腺癌细胞周期
细胞周期包括G1、S、G2、M 4个时期,细胞正常的分裂、增殖、分化、衰老维持着机体自身的稳定,而癌细胞可逃脱细胞周期的调控,不受限制地分裂、增殖。多糖类成分可通过调节由细胞分裂周期蛋白(cell division cycle,CDC)、细胞周期蛋白依赖性激酶(cyclindependent kinases,CDK)和细胞周期蛋白依赖性激酶抑制因子(CDK inhibitor,CKI)组成的蛋白激酶复合物阻滞胰腺癌细胞周期[28];激活后的CDK抑制剂p21和p27可通过与调控G1期的细胞周期蛋白(Cyclin)-CDK复合物结合并诱导其失活[29],同时影响Cyclin A的表达,从而抑制S期的DNA复制。
褐藻多糖可通过调控胰腺癌细胞PANC-89、PancTu1、PANC-1的细胞周期抑制剂 p57、TP53INP1、Cyclin E2、E2F 转录因子 1(recombinant E2F transcription factor 1,E2F1)、增殖细胞核抗原(proliferating cell nuclear antigen,PCNA)的表达,抑制下游细胞周期蛋白CDC45、CDC7、CDC25A的表达,诱导细胞周期停滞,从而有效杀死癌细胞[30]。红花多糖HH1-1可降低胰腺癌细胞BxPC-3和AsPC-1中细胞周期蛋白Cyclin A2、蛋白激酶B(protein kinase B,Akt)和CDK2蛋白水平,上调p21表达水平,显著增加S期细胞比例,从而抑制癌细胞增殖[31]。玉米须粗多糖S1干预胰腺癌细胞BxPC-3后,可见癌细胞核皱缩及凋亡小体,该多糖通过阻断EGFR/磷脂酰肌醇 3-激酶(phosphoinositide-3 kinase,PI3K)/Akt/环磷腺苷效应元件结合蛋白(cAMP-response element binding protein,CREB)信号通路将细胞周期阻滞在S期,从而抑制BxPC-3细胞增殖[32]。姬松茸粗多糖能下调胰腺癌细胞MIAPaCa-2、PCI-35的细胞周期促进基因Cyclin D1、Cyclin A2、Cyclin B2和CDK6的表达,诱导G0/G1期细胞阻滞及Caspases依赖性凋亡,从而抑制胰腺癌细胞增殖[33]。
由上文可知,多糖类成分可通过调节CDC、CDK与Cyclin家族细胞因子,将胰腺癌细胞周期阻滞在S期,从而发挥抑癌作用。
1.5 抑制胰腺癌细胞迁移与侵袭
癌症转移是恶性肿瘤的特征之一,肿瘤细胞迁移是转移的先决条件,而后发生侵袭与浸润,最终导致转移。胰腺癌是公认的转移性极强的癌症,80%~85%的患者在确诊时已发生转移[3]。转移性癌细胞及邻近内皮组织中常观察到基质金属蛋白酶家族(matrix metalloproteinase,MMPs)过表达现象。MMPs能降解细胞外基质(extracellular matrix,ECM)中多种蛋白成分,其过表达会破坏基质屏障,使肿瘤细胞能够侵入周围组织和血管[34]。
褐藻多糖可以下调胰腺癌细胞PANC-1中MMP-2和MMP-9的表达,表现出抑制癌细胞迁移和侵袭的能力[35]。云芝多糖PSK在缺氧条件下以配体非依赖性方式抑制缺氧诱导因子1α(hypoxia-inducible factor 1α,HIF-1α)和刺猬(Hedgehog,Hh)信号通路,调控下游MMP-2、MMP-9表达水平,从而降低胰腺癌细胞AsPC-1的侵袭性[34]。体内和体外实验表明,太子参多糖H-1-2可以抑制胰腺癌细胞PANC-1中前梯度蛋白2(anterior gradient 2,AGR2)和HIF-1α的表达,并能抑制小鼠体内胰腺癌异种移植瘤的生长[36]。三七花多糖可抑制骨形态发生蛋白2(bone morphogenetic protein 2,BMP2)信号转导,下调Id-1表达,从而抑制肿瘤血管生成,进而削弱胰腺癌细胞BxPC-3的迁移与侵袭能力[37]。在胰腺癌细胞BxPC-3异种移植小鼠模型中,褐藻中的岩藻依聚糖能抑制肿瘤血管生成,下调磷酸化血管内皮生长因子受体 2(vascular endothelial growth factor receptor 2,VEGFR2)和血小板-内皮细胞黏附分子1(platelet endothelial cell adhesion molecule-1,PECAM-1)的表达,从而发挥抗癌活性[38]。黑枸杞多糖LRP3-S1可抑制胰腺癌细胞BxPC-3、PANC-1和AsPC-1增殖,同时通过调控MAPK信号通路和抑制FAK与Akt、糖原合成酶激酶3β(glycogen synthase kinase-3β,GSK-3β)的下游磷酸化,从而抑制BxPC-3细胞的侵袭性[39]。此外,黄芪多糖与阿帕替尼联合使用后可显著抑制MMP-9、Bcl-2的表达,从而抑制癌细胞迁移能力[17]。
由上文可知,多糖类成分能调节MMPs表达,通过激活Hh、MAPK信号通路发挥抑制胰腺癌细胞迁移与侵袭的作用。
综上,不同来源的多糖类成分抗胰腺癌的作用机制如表1所示。
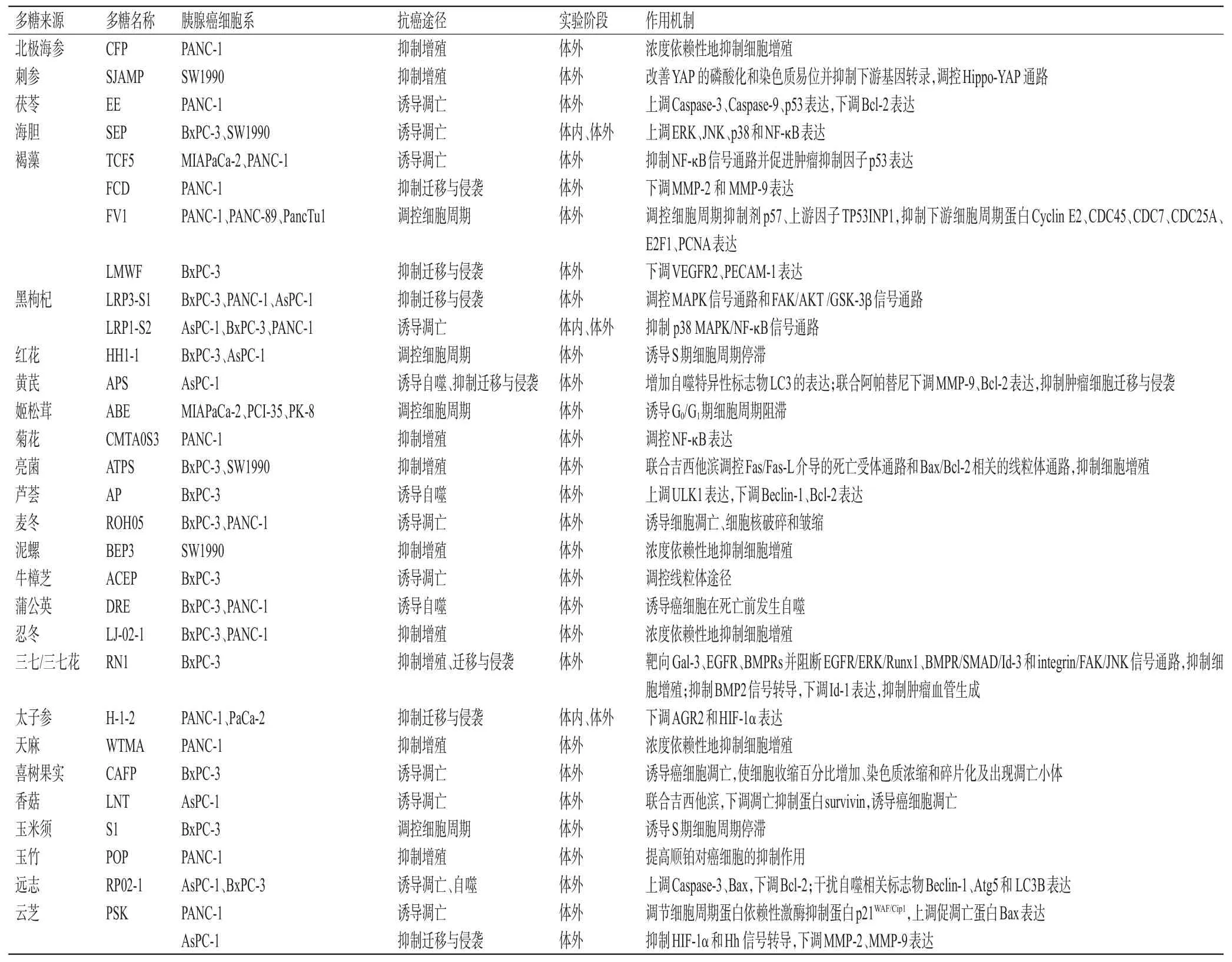
表1 不同来源多糖类成分抗胰腺癌的作用机制
2 总结与展望
多糖类成分能通过调控细胞因子的表达靶向多条信号通路,以抑制细胞增殖、诱导细胞凋亡与自噬、阻滞细胞周期、抑制细胞迁移与侵袭,从而发挥广泛抗胰腺癌的作用。NF-κB信号通路是多糖类成分发挥抗胰腺癌作用的主要通路,该通路在参与免疫应答等正常生理功能的同时还调控IL、MMPs、VEGF、CDC、CDK等细胞因子,并且能激活下游可以直接杀伤肿瘤细胞的TNF,以多种形式维持机体稳定;Hippo-YAP信号通路参与胰腺腺泡细胞的分化过程,当腺泡细胞被非正常抑制时,组织器官会过度增殖并导致胰腺癌的发展,而多糖类成分能抑制该通路下游多个效应因子的表达,减缓胰腺癌进程;Integrin信号通路主要介导细胞和细胞之间以及细胞和细胞外基质之间的相互识别和黏附,能驱动肿瘤微环境对抗癌药物产生耐药性,而多糖类成分可作为抑制剂阻断该信号通路,减少抗癌药物的耐药性,从而增强抑癌效果。
目前对于多糖类成分调控细胞因子的研究多集中在Bax、Bcl-2、p53等常见凋亡因子上,但如HIF-1α是癌细胞在实体瘤缺氧环境下存活的必要调节因子,YAP可以将癌细胞重新编程为肿瘤干细胞,与肿瘤的发生、发展和转移密不可分,FAK、P13K、SMAD等细胞因子也作为中间递质参与了多条抑制胰腺癌的途径,这些细胞因子间及与信号通路的关联调控模式将是今后明晰多糖类成分抑制胰腺癌作用机制的研究重点(图1)。
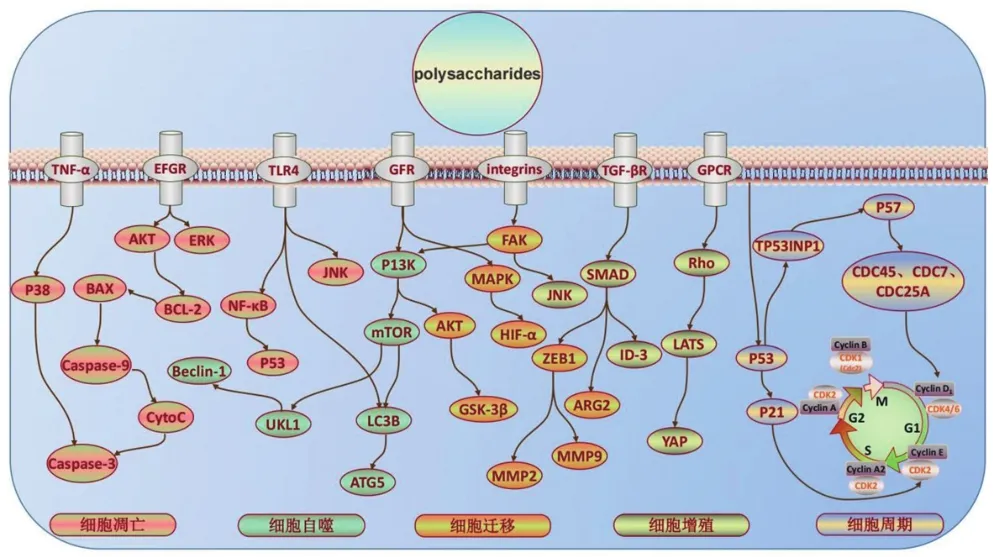
图1 多糖类成分抗胰腺癌作用的细胞因子网络
随着多糖分离纯化及分析技术的进步,对多糖结构及生物学功能的研究也日益深入。研究者在多糖抗癌构效关系的研究中发现,多糖的分子量、电荷数、硫酸化程度与位置均与其抗癌活性密切相关。例如:褐藻中分离出的5个多糖类成分中,硫酸盐与糖醛酸含量最高的组分显示出最佳的抗癌活性[15];分子量与单糖组成会影响枸杞多糖的抗胰腺癌能力[39];泥螺多糖抗胰腺癌活性也推测与其硫酸化程度有关[27]。但是关于多糖类成分抗胰腺癌的构效关系仍需深入研究,如目前尚不知晓多糖如何通过单糖组成、糖苷键、立体结构对胰腺癌产生抑制作用。此外,多糖的羧甲基化、硫酸化、磷酸化、硒化和乙酰化等衍生化产物的抗胰腺癌作用也值得深入探讨。
截至目前,多糖类成分抗胰腺癌的作用机制多是由体外细胞实验验证的,通过对其机制的进一步研究与探索,在体外实验基础上进行体内动物模型验证,两者相互结合,并结合结构修饰与剂型调节,将会为临床胰腺癌治疗药物的开发带来新的希望。
猜你喜欢
保健医苑(2022年6期)2022-07-08
奥秘(创新大赛)(2019年9期)2019-10-09
小哥白尼(趣味科学)(2019年1期)2019-04-12
特别健康(2018年3期)2018-07-04
中成药(2017年9期)2017-12-19
中成药(2017年7期)2017-11-22
奥秘(2017年5期)2017-07-05
天津医药(2016年9期)2016-10-20
中华老年多器官疾病杂志(2016年7期)2016-04-28
中国中医药现代远程教育(2014年22期)2014-03-01
