
Polysaccharide extract from Rosa laevigata fruit attenuates inflammatory obesity by targeting redox balance and gut interface in high-fat diet-fed rats
2023-01-21 05:02XuejiaoZhangChenzhongJinHanLiuYihongHuYunyunZhouWeiguoWuSiQin
食品科学与人类健康(英文) 2023年2期
Xuejiao Zhang,Chenzhong Jin,Han Liu,Yihong Hu,Yunyun Zhou,Weiguo Wu,Si Qin
a Lab of Food Function and Nutrigenomics,College of Food Science and Technology,Hunan Agricultural University,Changsha 410128,China
b College of Food and Chemical Engineering,Shaoyang University,Shaoyang 422000,China
c Collaborative Innovation Center for Field Weeds Control,Hunan University of Humanities,Science and Technology,Loudi 417000,China
Keywords:Rosa laevigata Polysaccharides Multi-omics Redox Gut microbiota
ABSTRACT Low-molecular-weight polysaccharides (RLPs) extracted from Rosa laevigata fruits showed multiple biofunctions in Asia.This study aimed to investigate its anti-inflammatory obesity effect in high-fat dietfed rats and further elucidated the underlying molecular mechanism by multi-omics methods.The results showed that RLPs administration had significantly restored immune organ indexes and reduced body weight gain.RNA-seq revealed that the effect of RLPs was partially attributed to its regulation on PPARs signaling by increasing the expressions of Scd,Acox3 and Hmgcs2,and on other redox-related pathways by decreasing the expressions of Cyp2e1,Il1-r1 and Lbp.Moreover,16S rRNA sequencing coupled with metagenome sequencing showed that RLPs had significantly reduced the ratio of Firmicutes/Bacteroidetes from 8.01 to 2.37,and significantly increased the relative abundances of Alistipes,Prevotella,and Akkermansia from 0.36%,1.10% and 2.61% to 0.65%,2.37% and 4.42%,respectively.Spearman correlation analysis result indicated that the abundances of Lachnospiraceae,Prevotella and Bacteroidales were significantly negatively correlated with obesity phenotype,liver function and inflammatory factors.These results revealed that RLPs exerted significant anti-inflammatory obesity property partially via regulation on gut microbiota interface and the redox balance.Therefore,RLPs could be a promising functional food resource with the potential for redox imbalance-related diseases chemoprevention.
1.Introduction
Obesity is deemed as a global health issue currently,which can increase the risk of chronic metabolic diseases including chronic inflammation,diabetes,hypertension,etc.,leading to an extensive burden to society and the economy [1,2].Although genetic and environmental factors are proposed to main reasons for obesity,the basic cause was attributed to an imbalance between dietary intake and energy consumption [3].Scientists are paying more and more attention on effective therapies,among which,natural polysaccharides were used as potential functional ingredients or drug candidate with potent anti-obesity and anti-inflammation activities [4].Obesity was found to correlate with inflammatory response and human intestinal microbiota in previous studies [5-7].It is prone to induce chronic and mild inflammation associated with an improved risk of obesity after disturbance of the gut microbiota and transforms in gut permeability [8].The most of microbial dwellers have the functions of digesting food and regulating immunity,which is due to the formation of a stable microbial ecosystem [2].The disorder of intestinal microbiota is not only related to the prevalence of obesity,but also closely related to metabolic syndrome [2].
Apart from starch and glycogen,natural polysaccharides are long-chain monosaccharide polymeric carbohydrates,which exist many functionalities [9].They could reduce oxidative stress [10],attenuate inflammation [7],improve immunity and obesity [11],and ameliorate compositions of intestinal microbiota [12].In particular,functional polysaccharides from traditional Chinese medicinal plants,such asGanoderma lucidum[13],Morinda citrifolia[14],Hippophae rhamnoides[15],andArctium lappa[6],showing various tonic effects.The polysaccharide isolated from theOphiopogon japonicusroots has potent effect on weight loss and hypoglycemic,mainly via decreasing Firmicutes/Bacteroidetes ratio and adjusting the disordered intestinal microbiota to the normal one with complicated biotransforms [16].
Rosa laevigataand its fruits have been safely used for thousands of years as drugs or foods in eastern countries especially in southern regions of China [17,18].Recent studies showed that the special fruits possess multiple pharmacological effects bothin vitroandin vivo,including antioxidant profiles [19],hypolipidemic effect [18],anti-inflammatory [20],immunomodulatory [21],cardiovascular protection [22],and neuroprotective effect [23],which were reported to be attributed to the presence of various bioactive compounds such as triterpenoids,steroids,phenolics,tannins,organic acid,and polysaccharides [17].Previous studies demonstrated that the polysaccharides isolated fromR.laevigatafruits had hypolipidemic [24],antioxidant [18],immunomodulatory [21],antitumor [25],and neuroprotective activities [23].For example,Zhan et al.[21]revealed a novel acid polysaccharide isolated from theR.laevigatafruits had immunomodulatory activities by activating MAPK and NF-κB signaling pathways.Zhang et al.[25]demonstrated a neutral watersoluble polysaccharide extracted and purified from the fruits ofR.laevigatashowed immunomodulatory activities and significantly antitumor activities by inhibiting tumor cell proliferation and migration and blocking angiogenesis.
The characterization of homogeneous polysaccharide structure inR.laevigatafruits and the biological screening are crucial for the further development of the fruits ofR.laevigata.Combined with these previous studies,the activity of low-molecular-weight polysaccharides from the fruits ofR.laevigatahave been reported rarely.In the previous study,low-molecular-weight polysaccharides (RLPs) fromR.laevigatafruits were extracted and their structure was characterized [24].The present study aimed to evaluate the effect of RLPs on inflammatory obesity and gut microbiota balance in high-fat diet (HFD)-fed rats,and further elucidate the underlying molecular mechanisms by multi-omics methods.
2.Materials and methods
2.1 Materials
The wildR.laevigatafruits were picked in Wugang city of Hunan province,China,and they were identified as ripeR.laevigatafruits by Professor Zefa Liu of Agricultural Research Institute of Hunan Province.RLPs were isolated by extraction,separation and purification,which contained three low-molecularweight polysaccharides RLP-1a ((90.78 ± 2.56)%),RLP-2a((86.34 ± 2.31)%),and RLP-3a ((82.61 ± 1.92)%) with 9 004,8 761,and 7 571 Da,respectively,and their structures were characterized as described in our previous paper [24].Briefly,structural characterization showed that RLP-1a was consisted of rhamnose,arabinose,xylose,glucose,and galactose with the ratio of 3.14:8.21:1:1.37:4.90,RLP-2a was composed of rhamnose,mannose,glucose,and galactose with the ratio of 1.70:1:93.59:2.73,and RLP-3a was composed of rhamnose,arabinose,xylose,mannose,glucose,and galactose with the ratio of 6.04:26.51:2.05:1:3.17:31.77.The NMR analyses revealed that RLP-1a,RLP-2a and RLP-3a contained 6,4 and 6 types of glycosidic linkages,respectively.
RLPs was extracted in a constant temperature bath of distilled water (90 °C) for 2 h,and then precipitated overnight with aqueous ethanol,and the final concentration was 80% (V/V).Then the precipitates were purified on columns packed with anion exchange resin (DEAE-52,Whatman,Sanford,UK) and Sepharose resin(Sepharose CL-4B,Whatman,Sanford,UK) continuously,after dialyzed and lyophilized,RLPs were obtained.
2.2 Animals and treatments
A total of 40 male rats with the age of 8-week-old were purchased from Hunan SJA Laboratory Animal Co.,Ltd..They were given adaptive feeding for one week before the experiment with a constant temperature and humidity environment.The animal experiments were conducted based on the National Research Council’s Guidelines for the Care and Use of Laboratory Animals and were approved by the Committee on the Ethics of Animal Experiments of Hunan Agricultural University,China (protocol code: 2020-43).
The rats were randomly separated into 4 groups with 10 rats in each,including control (CN) group,HFD group,HRLP(HFD+800 mg/kg BW RLPs) group and LRLP (HFD+200 mg/kg BW RLPs) group.Sample food and water consumption were assigned 3 times/week/cage and the averages of food and water consumed were calculated weekly.The rats were administrated continuously for 12 weeks and sacrificed after anesthesia.Serum and internal organs were immediately collected,rapidly frozen with liquid nitrogen,followed by storage at -80 °C until analysis.At the end of the experiment,the length from nose to anus and the total body weight of the rats were measured.Oral glucose tolerance test (OGTT) was performed the day before the completion of the experiment.Lee’s index was calculated according to the following formula (1) [26].The formula(2) listed below has been used for calculating the thymus and spleen indices [27],as well as the formula (3) has been used for operating the homeostasis model assessment of insulin resistance (HOMA-IR) [28]:

2.3 Histologic analysis
Tissues of the liver,epididymal adipose,and colon were fixed with formalin,then were dewatered gradually with 70% -100% ethanol.After dewatering,these tissues were transparent with xylene,embedded by paraffin and cut into slices by a frozen slicer (Yamato,Saitama,Japan).Liver,epididymal adipose,and colon sections were stained with hematoxylin-eosin staining followed by observations under a microscope (Nikon,Tokyo,Japan) at 100 ×.
2.4 Determination of biochemical indexes
The levels of serum fasting blood glucose (FBG),fasting insulin(FINS),aspartate transaminase (AST),alanine transaminase (ALT),γ-glutamyl transpeptidase (GGT),total cholesterol (T-Cho),and high-density lipoprotein (HDL) were measured using an automatic biochemical analyzer (XinRui,XR220PLUS,Zhongshan,China).The level of glutathione oxidase (GSH-Px) in liver,as well as secretion of IL-1β,IL-6,TNF-α,MCP-1,and IFN-γ cytokines in serum were analyzed by ELISA Kits (Zecen Biotechnology Co.,Ltd.,Jiangsu,China) method.
2.5 Transcriptome analysis of rat livers
The total RNA was obtained from liver tissue by Trizol Kit(Invitrogen company) based on the manufacturer’s instructions.Nanophotometer along with Bioanalyzer 2100 system (Agilent Technologies,CA,USA) was used to detect the purity of RNA and the integrity of the RNA,respectively.Illumina sequencing was performed after the library passed the quality inspection,as we covered in the previous article [24].The pathway with FDR (false discovery rate) ≤ 0.05 was defined as the pathway with significant enrichment among the differentially expressed genes.
2.6 Short-chain fatty acids analysis
SCFAs were extracted from feces in line with the methods previously described [7,28].The instrument used to determine the contents of SCFAs was gas chromatography-mass spectrometry(GC-MS,QP2010SE) (Shimadzu Co.,Kyoto,Japan) equipped with a DB-Wax capillary column (30 m × 0.25 mm).
2.7 Gut microbiome and metagenome analysis
Gut microbial analysis was performed by 16S rRNA gene sequencing as well as metagenome sequencing with specialized software.The PowerSoil®DNA Isolation Kit (Mobio,San Diego,CA,USA) was conducted to extract total genomic DNA of intestinal microbiota from feces samples.The primers,which used for amplification of V4 variable region of the 16S rRNA gene by PCR,were 515F (GTGCCAGCMGCCGCGGTAA) and 806 R(GGACTACHVGGGTWTCTAAT) [18].The 16S rRNA gene sequencing was carried out on the Illumina HiSeq Ion S5TMXL platform (Thermo Fisher Scientific,MA,USA) based on the producer’s method.In the metagenome sequencing,Qubit®dsDNA Assay Kit in Qubit®2.0 Fluorometer (Life Technologies,CA,USA)was used to measure DNA concentration.A total amount of 1 μg DNA per sample was used for the purpose of import material for the DNA sample preparations.
2.8 Statistical analysis
All experimental results were presented as mean ± standard deviation (SD) unless otherwise stated,and SPSS 20.0 (SPSS Inc.,Chicago,IL,USA) was used for variance and significant difference analysis,as well as Tukey test.P<0.05 was considered statistically significant.
3.Results
3.1 LPs affected weight gain and insulin resistance
Being overweight is a crucial clinical symptom of obesity.The body weight gain of HFD was (316 ± 10.71) g,which is significantly higher than CN,HPLP,and LRLP group with (186 ± 6.63),(226.33 ± 6.14),and (252 ± 7.93) g,respectively.The model group showed a significant increase in Lee’s and liver index level compared to those of the normal group,and these were alleviated by treatment of RLPs,as shown in Fig.1.Besides,the spleen index was found to be decreased significantly but the thymus was increased by RLPs treatment (Fig.1C).We have analyzed the contents of TG,TC and LDL-C in serum,which were reported in our previously published papers [24].The analysis results are simplified as follows:compared with the HFD group,the HRLP group reduced TG(P <0.05),TC (P<0.05),FFA (P<0.05),and LDL-C (P<0.05)levels by 16.3%,47.0%,27.8%,and 14.8%,respectively,and the HDL-C levels significantly increased by 35.0% (P<0.05),indicating that RLP with higher dosage intervention had stronger hypolipidemic activities in rats,and the hypolipidemic activity of LRLP was weaker than that of HRLP.FBG level showed a significant difference between the HFD group with the other groups (Fig.1D,P<0.05),and OGTT showed that a high dose of RLPs improved the glucose tolerance which was impaired in HFD-induced rats (Fig.1E,P<0.05).FINS level and HOMA-IR index was (2.39 ± 0.31) μg/L and 15.58 ± 2.75 respectively in the treatment of high dosage RLPs,which significantly decreased when we made a comparison with the HFD group (Fig.1F,P<0.05).These data suggested that RLPs exerted their anti-obesity effect via bodyweight control,lowering HFD-induced serum insulin level and improving insulin resistance.

Fig.1 Effects of RLPs on the body weight gain (A),Lee’s index (B),organ index (C),FBG (D),OGTT (E),FINS (F) and HOMA-IR index (G).The values represent mean ± SD (n ≥ 5),and showed significant difference at the P <0.05 levels.
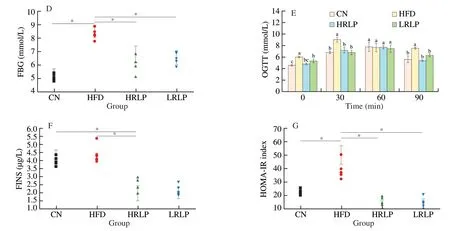
Fig.1 (Continued)
3.2 RLPs reduced lipids accumulation,peroxidation,and liver injury
Inflammatory obesity-related tissue and organ lesions are key indicators for clinical diagnosis.Therefore,hepatic,epididymis adipose,and colonic H&E staining results were conducted to investigate the clinical effect of RLPs on rat tissues.As shown in Fig.2A,HFD led to increasing lipid droplets accumulation and bubble-like degeneration of hepatocytes,whereas this result was reversed after RLPs intervention dose-dependently.In Fig.2B,HFD caused epididymis adipocyte enlargement and less quantity,and RLPs supplementation alleviated this effect.In the HFD group,the epithelial cells of intestinal mucosa were damaged and disappeared continuously and the glands were distorted,but RLPs treatment significantly improved the pathological changes of the colon (Fig.2C).
The levels of ALT,AST,and GGT are clinically detected as biomarkers for liver damage.HFD group showed increased ALT,AST,GGT,and RLPs treatment showed an opposite trend in serum(P<0.05) (Figs.2D-F).Furthermore,by comparison with the CN group (P<0.05),the activities of GSH-Px in the liver were markedly reduced in the HFD group.However,the activity of GSH-Px was notable raised again markedly by RLPs treatment,shown in Fig.2G (P<0.05).It can be seen from the above results that RLPs could mitigate HFD-guided liver injury and lessened oxidative stress in rats.
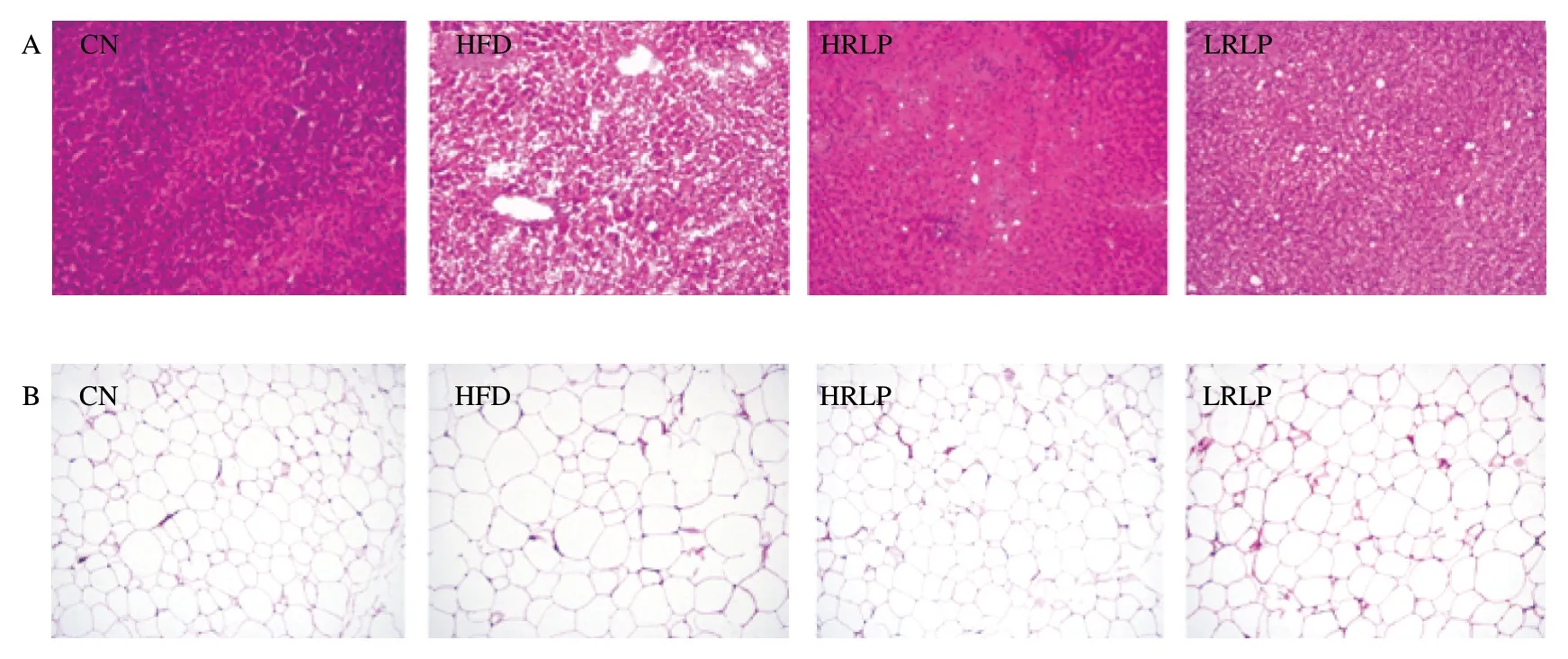
Fig.2 RLPs reduced hepatic lipid droplets (A),epididymal adipocyte enlargement (B),colonic mucosa damage (C) and serum levels of ALT (D),AST (E) and GGT (F),and liver levels of GSH-Px.Hematoxylin-eosin (H&E) staining was performed for each group of sliced tissue,and magnified 100 times by fluorescence microscope (Nikon,Tokyo,Japan).The data were reported as mean ± SD (n≥5),and showed significant differences at the P <0.05 levels.
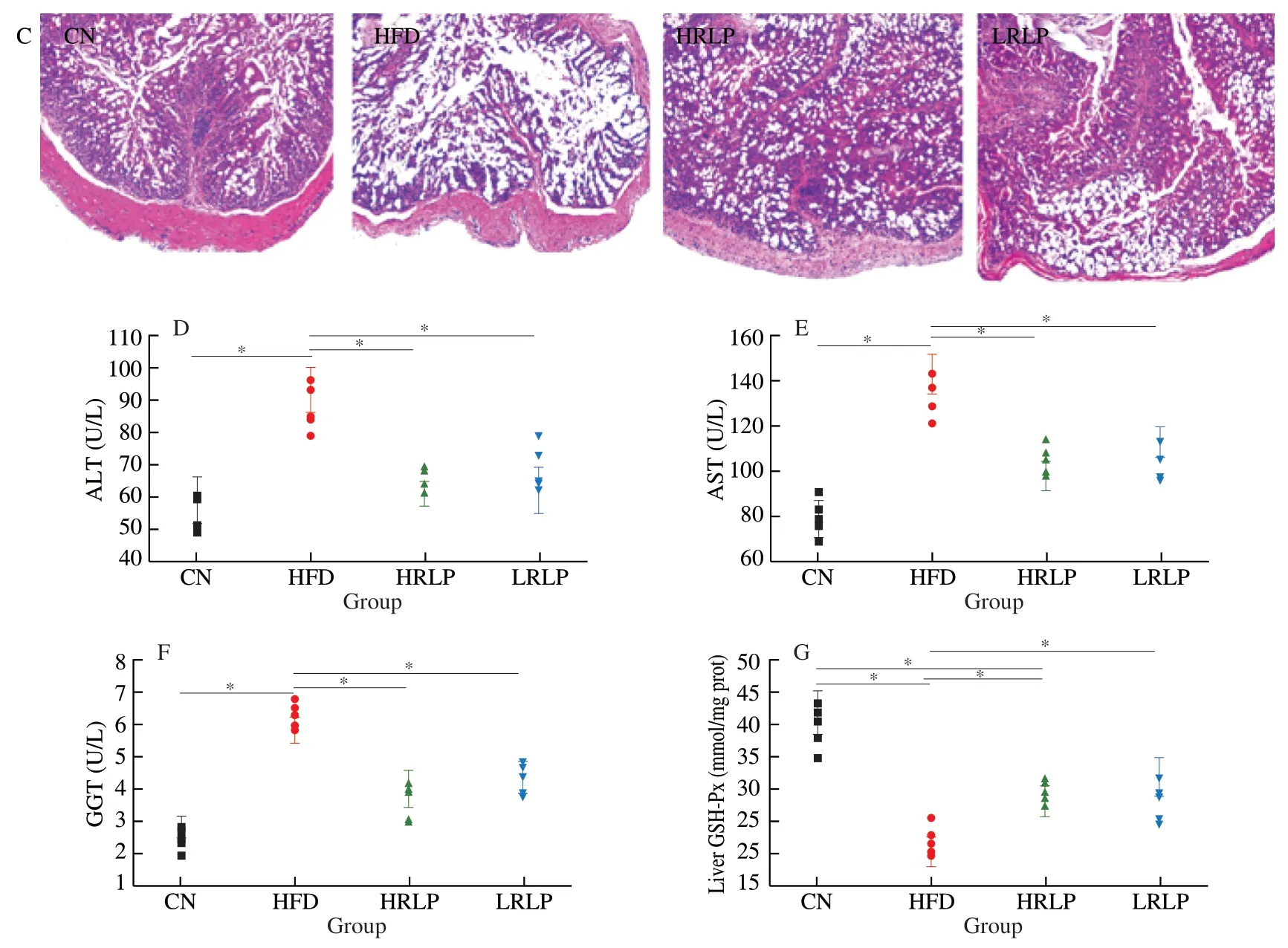
Fig.2 (Continued)
3.3 Effects of RLPs on inflammation-related cytokines in serum
As pro-inflammatory cytokines play an important role in the prosses of inducing inflammation [29],five kinds of cytokines were determined in serum to evaluate the anti-inflammatory action of RLPs.The results were represented in Fig.3,serum levels of TNF-α,IL-1β,IL-6,and MCP-1 were significantly increased but IFN-γ was decreased in the HFD group (P<0.05),indicating that a HFD can trigger an internal inflammatory response.After the intervention of RLPs at the two tested dosages,the promotions of TNF-α,IL-1β,IL-6,and MCP-1 were effectively suppressed.Polysaccharides are reported to exert anti-inflammatory effect via decreasing the plasma levels of inflammatory cytokines including IL-6,IL-1β and TNF-α [30,31].As shown in Fig.3E,in contrast to the HFD group,the secretion of IFN-γ was remarkably raised in RLPs treated groups,which could induce T cell proliferation and differentiation into Th1 cells,and participate in cellular immunity and eliminate intracellular pathogens [32].Moreover,RLPs also significantly down-regulated the expressions of IL-1 receptor 1 (Il-1r1,P<0.001) and LPS-binding protein (Lbp,P<0.005) (Fig.3F),which are upstream genes of the NF-κB pathway.

Fig.3 Effects of RLPs on the inflammatory cytokine levels of TNF-a (A),IL-1β (B),IL-6 (C),MCP-1 (D),IFN-γ (E),and rats liver gene expression of Lbp and Il-1r1 (F).The rats were three for each group,and the data were expressed as mean ± SD,and significantly different at the P <0.05 levels.The mRNA expressions were determined by RNA-Seq,and set the fold change of the control group to 1 and then calculate the fold change for each group (n ≥ 5).
3.4 Effects of RLPs on hepatic lipids metabolism and redox balance
RNA-seq analysis on differentially expressed genes (DEGs) in liver tissue of rats fed with HFD as shown in Fig.4A,a total number of 1 566 DEGs were gained from the HFD group and the CN group,including 783 up-regulated DEGs and 783 down-regulated DEGs,and the DEGs between HRLP and HFD groups were 1 109,among which 579 were up-regulated DEGs and 530 were down-regulated DEGs.The Venn map reflects the number of DEGs across three comparison groups,which were the HRLP vs HFD group,the LRLP vs HFD group,and the HRLP vs LRLP group (Fig.4B).The number of unique DEGs in these three comparison groups was 695,278,and 333,respectively,and the number of common DEGs was 12.For DEGs between HRLP and HFD,49 biological processes and 7 molecular functions were significantly enriched in GO terms.The 49 biological processes were mainly classified into the oxidation-reduction process,lipid metabolism process,small molecule metabolism process,carbohydrate metabolism process,and ATP metabolism process,etc.,and the 7 molecular functions mainly includes oxidoreductase activity and DNA binding of the regulatory region.These results showed that treatment with RLPs altered the expression of genes related to the lipid metabolism and redox processes.With the analysis by Kyoto Encyclopedia of Genes and Genomes (KEGG),the differential genes were mainly in the peroxisome proliferator-activated receptors(PPARs) signaling pathway,steroid biosynthesis pathway,fatty acid metabolism,unsaturated fatty acid biosynthesis,metabolism of xenobiotics by cytochrome P450,and non-alcoholic fatty liver disease(NAFLD) pathway (Fig.4C and 4D).PPARs plays a significant role in adjusting liver lipid metabolism.PPARs including PPARα,PPARβ,and PPARγ,are associated with fat metabolism with different functions.PPARαis mainly participated in the regulation of fatty acid uptake,lipogenesis,fatty acid oxidation,cholesterol metabolism,and lipid transport in liver fat metabolism [33].As shown in Fig.5,the expression of the fatty acid oxidation geneAcox3(P<0.05)and the stearoyl-coenzyme A desaturase geneScd(P<0.001) were significantly increased in the HRLP group in contrast to the HFD group,indicating RLPs can enhance fatty acid oxidation and regulate the metabolism of long-chain fatty acids.
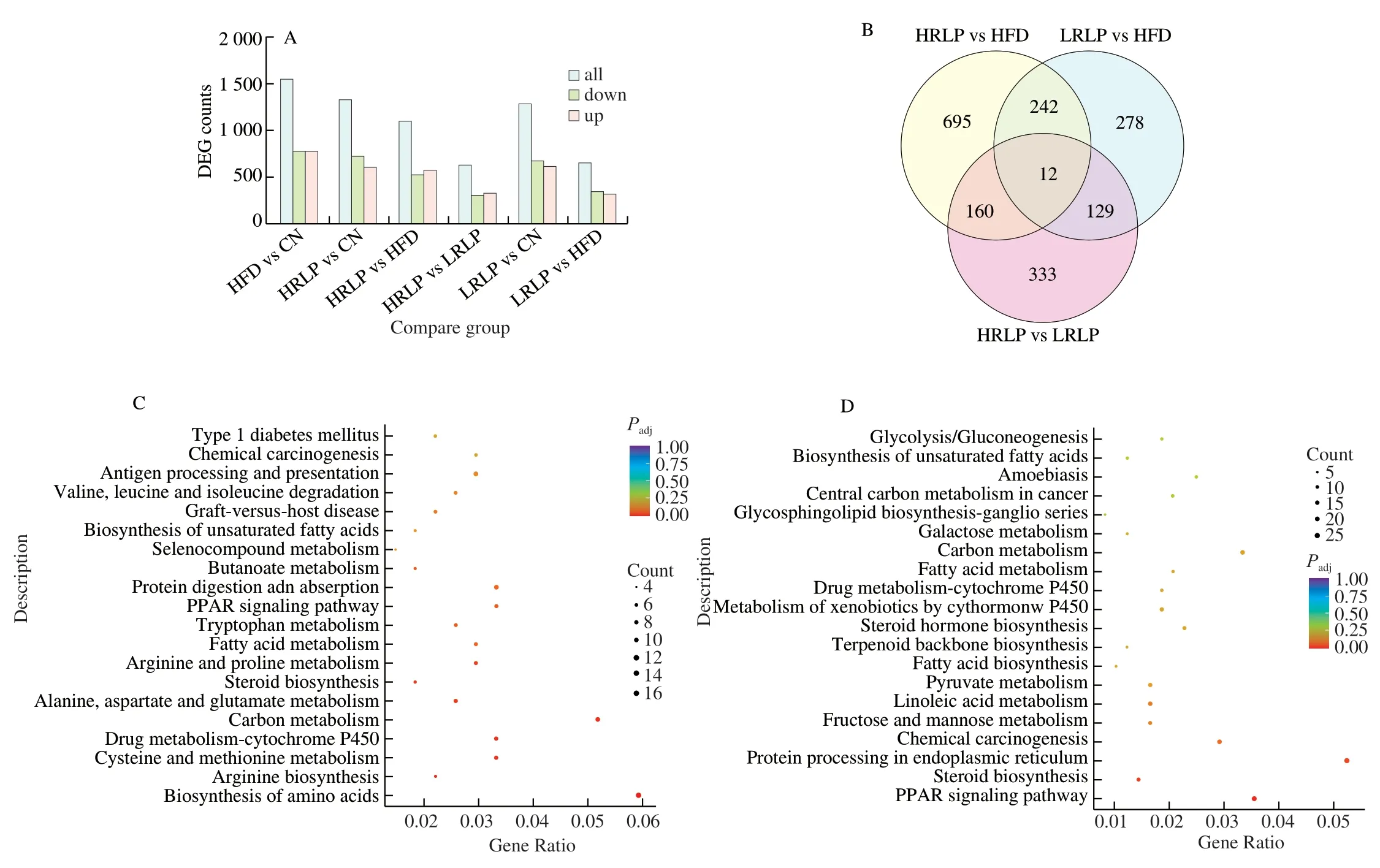
Fig.4 Effects of RLPs on related pathways and liver gene expression.(A) The number of DEGs across six comparison group;(B) Venn map of DEGs across three comparison group;(C) KEGG analysis of DEGs between LRLP and HFD groups;(D) KEGG analysis of DEGs between HRLP and HFD groups.
HFD could induce oxidative stress inin vivolevel,andCyp2e1is considered to be the main cause of the overproduction of oxidative stress markers ROS in the life system [34].As shown in Fig.4C,HFD indeed increased the expression ofCyp2e1,intervention with RLPs significantly decreased that (P<0.001).This result implied that a HFD successfully lead to oxidative stress in rats,and RLPs improved lipid metabolism by alleviating oxidative stress to maintain the redox balance.
3.5 RLPs affected the production of SCFAs
SCFAs produced by gut microbiota,which play critical roles in maintaining host health.The levels of SCFAs were determined in the feces of the four groups of rats (Table 1).Overall,our results represented that the HFD decreased SCFAs levels compared to normal control rats,which corresponded to former reports [35].By Comparison with the HFD group,rats treated with RLPs showed a significant increase in the content of propionic acid,butyric acid,isovaleric acid,and valeric acid (P<0.05).These results corresponded to the recent research of purple sweet potato polysaccharides producing high contents of SCFAs in colitis mice [36].
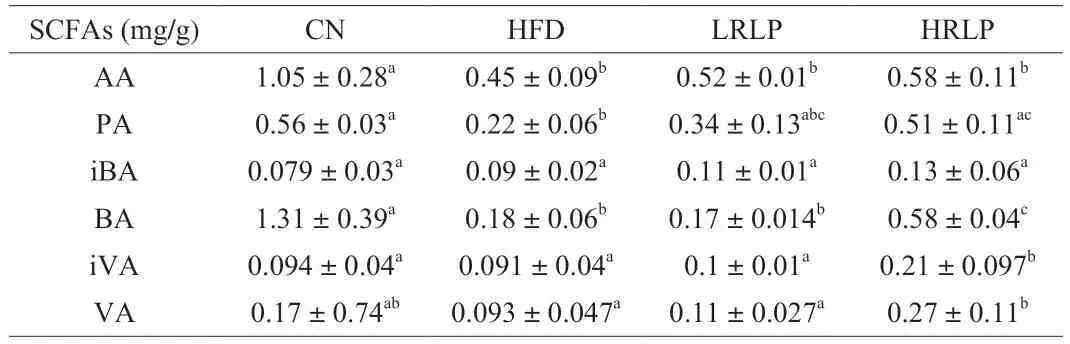
Table 1 Effects of RLPs on production of SCFAs.
3.6 Effects of RLPs on gut microbiota composition
16S rRNA gene sequencing and metagenome gene sequencing were used to analyze the composition,characteristics,and relative abundance of gut microbiota.The diversity and richness of microbial community can be reflected by alpha-diversity analysis.Alphadiversity is reflected by Observed Species,Chao,and Shannon indices.Among them,Chao and observed species indices represents the richness of gut microbiota and higher values mean higher richness,while Shannon index both represent the diversity of gut microbiota,lower values of Shannon index mean higher diversity.As shown in Table 2,compared with CN group,HFD significantly reduced Chao index and observed specifications index (P<0.05),and also significantly reduced Shannon index,suggesting HFD had caused the decrease in richness and diversity of gut microbiota in rats.The intervention of RLPs increased the Chao index and observed species index,but there was no significant difference (P>0.05).These results suggest that RLPs can increase the richness of intestinal flora in rats fed with high-fat diet to a certain extent.In addition,Coverage index was high with average value of 0.998 in four groups,suggesting sequence similarity was >97% and the sequencing results well reflected the actual status of intestinal flora in our study.
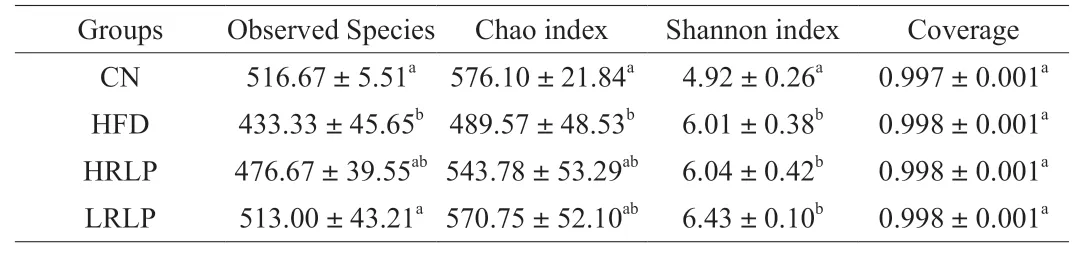
Table 2 Alpha-diversity index in each group.
After treated with HFD,the intestinal barrier of rats was damaged and the intestinal environment was altered,which will result in changes of gut microflora compositions.the taxonomic composition at the phylum level mainly contained Bacteroidetes,Firmicutes,and Proteobacteria and their distribution patterns in four groups were different.As shown in Fig.6A,in comparison with the CN group,the relative abundance of Firmicutes was elevated increased in the HFD group,while the abundances of Bacteroidetes and Proteobacteria were decreased.However,supplementation with RLPs reversed this result.Some predominant taxa in each group were listed in Table 3.Family levels of Bacteroidaceae,Akkermansiaceae,Prevotellaceae,and Oscillospiraceae were increased by high-dose RLPs supplementation,while the abundance of Clostridiaceae and Ruminococcaceae were dose-dependently decreased by RLPs supplementation (Fig.6B).The Firmicutes/Bacteroidetes ratio was ascended in the HFD group but descended with RLPs supplementation (Fig.6C).Further analysis at the genus level showed that RLPs raised the relative abundances ofAkkermansia,Alistipes,andPrevotella,and reduced the relative abundances ofDoreaandBlautia(Fig.6D).
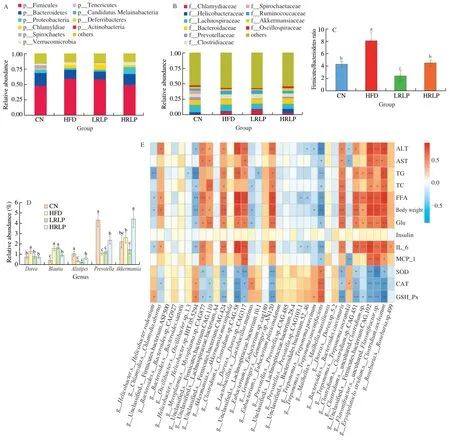
Fig.6 RLPs modified gut microbiota community by 16S rRNA sequencing coupled with metagenome sequencing and its correlation analysis with blood parameters.(A) Relative abundance of fecal microbiota from each group of mice at phylum level.(B) Relative abundance of the fecal microbiota from each group of mice at family level.(C) Firmicutes to Bacteriodetes rate of each treatment.(D) Relative abundance of Dorea,Blautia,Alistipes,Prevotella and Akkermansia muciniphila in each group.The data represent mean ± SD (n ≥ 3) and significantly different at the P <0.05 levels.(E) Spearman correlation analysis heat map(Metagenome sequencing) between key intestinal microbial taxa at species level and metabolic parameters of host at the end of the experiment.Red means a positive correlation and blue means a negative correlation,* indicates significance (P <0.05),** indicates extremely significant differences (P <0.01).
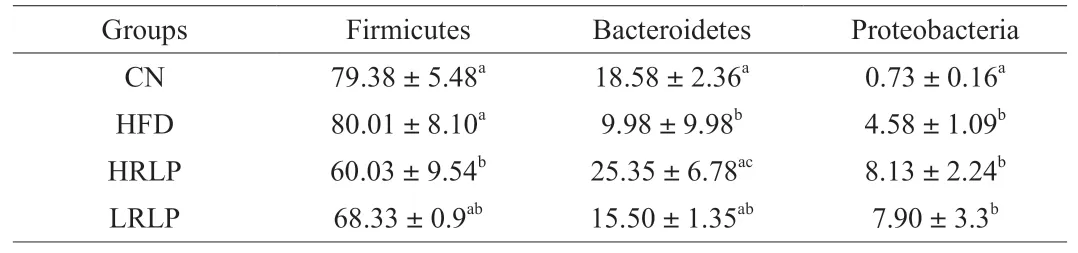
Table 3 Differences in gut microbiota at phylum levels of each group.
Finally,metagenomic sequencing identified bacteria of fecal samples at the species level.Spearman’s association analysis was used to investigate the correlation between gut microbiome community and serum biochemical characteristics.The results were represented in Fig.6E,there was a significant negative correlation among the serum levels of TG,TC,FFA,ALT,AST,glucose,IL-6,and MCP-1 with the relative abundances ofBacteroidalesbacterium 52-46,Prevotellasp.CAG.1031,Lachnospiraceae bacterium 10.1,Lactobacillus murinus,Helicobacter rodentium,Helicobactersp.MIT.05.5294,Treponema succinifaciens,andTreponema porcinum.Additionally,there was a significant positive correlation among the serum levels of TG,TC,FFA,ALT,AST,glucose,IL-6,and MCP-1 with the relative abundances ofClostridiumsp.CAG.451,Clostridiumsp.CAG.58,Clostridium cocleatum,Firmicutes bacterium CAG.102,Firmicutes bacterium CAG.110,Firmicutes bacterium CAG.424,Doreasp.CAG.317,Gemmigersp.An120,Roseburiasp.499,Bacteroides caecimuris,andMycoplasmasp.CAG.877.
4.Discussion
The present study applied multi-omics methods to explore the potential mechanism of the anti-inflammatory property of RLPs in HFD rats,and the results showed a protective effect of RLPs against inflammatory obesity by controlling body weight,lowering blood lipid and glucose indexes,regulating redox signaling pathways,and modulating gut microbiota.These data revealed that RLPs could be potent and promising drug candidates with the potential chemoprevention on redox-related disorders.
Both obesity and overweight not only increases the risk of developing type 2 diabetes,hyperlipidemia,fatty liver and cardiovascular disease,but also increases organ burden [37].Elevation of pro-inflammatory biomarkers levels have been found to be related to obesity,including IL-6,IL-1β and TNF-α [38].After 12 weeks’RLPs intervention,the serum levels of IL-1β,IL-6,MCP-1 and TNF-α were moderately but remarkably reduced,which probable partly give rise to the reduced activation of the TLR4-mediated inflammatory pathway.It has been reported that ROS was an inflammatory signaling molecule and mediator [39].Oxidative stress plays a crucial role in the pathogenesis of hyperlipidemia,which can bring about tissue damage through anti-oxidants imbalance [40].Hyperglycemiainduced increasing level of ROS is the main cause for various syndromes of DM [41].In current study,RLPs restrained oxidative stress caused by extreme cholesterol via enhancing the activity of antioxidant enzyme GSH-Px.High levels of TNF-α and IL-1β induced by HFD led to the occurrence of oxidative stress,which corresponded to a decrease in liver GSH-Px and an increase in liver ALT,AST,and GGT.Therefore,reducing oxidative stress induced by HFD can take precautions against a liver injury.Cyp2e1is the key factor in the production of oxidative stress and the most critical enzyme in the microsomal ethanol oxidation system.The addition of alcohol can lead to an increase inCyp2e1levels,which can accelerate the overproduction of ROS [42].In our research,the expression of theCyp2e1was down-regulated by the intervention of RLPs.Polysaccharides fromSchisandra chinensiswas reported to reduce theCyp2e1-dependent oxidative stress by inhibiting the up-regulation ofCyp2e1protein in HepG2 cells [43].
LPS can be handled by TLR4-mediated inflammatory pathways through handling and transporting byLbpand activation of macrophages [44].RLPs intervention down-regulated the expression of LBP and IL-1R1,which were reduced in HFD-fed rats may be partly by reducing activation of the TLR4-mediated inflammatory pathway.In addition,a boost in fatty acid oxidation by activating PPAR signaling with increasingAcox3andScdexpression.Thus,we considered that RLPs could activate theCyp2e1,toll-like receptor signaling pathways,and PPAR signaling pathways to regulate hepatic lipids metabolism and redox-related disorders.
Polysaccharides with different polarity,molecular weight,and structure extracted fromR.laevigatahave different biological activities.Such as a neutral water-soluble polysaccharide extracted and purified from the fruits ofR.laevigata,mainly consisted of glucose,arabinose,and galactose,and consisted of →5)-α-L-Araf-(1→,→2,5)-α-L-Araf-(1→,→3,5)-α-L-Araf-(1→,→4)-α-DGlcp-(1→,6)-α-D-Glcp-(1→,→3,6)-β-D-Glcp-(1→,→4)-α-DGalp-(1→,→6)-β-D-Galp-(1→,→2)-β-D-Xylp-(1→,terminalα-L-arabinose,and terminalβ-D-mannose,which showed immunomodulatory and antitumor activities [25].An acid polysaccharide with themwof 137 123 Da and a triple-helix conformation isolated from the pulp ofR.laevigatafruit,consisted of rhamnose (7.6%),arabinose (26.5%),xylose (3.5%),mannose (0.9%),glucose (5.7%),galactose (31.9%) and galacturonic acid (23.9%),and contained 16 types of glycosidic linkages,which showed antiinflammatory and immunomodulatory activity [21].
The intestinal microbiota is known to play an important role in a series of physiological procedures,including inflammatory reactions,immunological functions,and energy balance [45].In recent years,the association between chronic inflammation and intestinal flora imbalance has been confirmed [46].Indeed,certain intestinal bacteria and their metabolites can activate the mucosal immune system,leading to intestinal mucosal dysfunction [47].Dietary complex carbohydrates can be fermented partly or all by large intestine probiotic,and the SCFAs as their fermentative products of can affect microbes and the gut environment,and those also can regulate the efficiency of fat metabolism and energy intake of the host [48].Polysaccharides,as dietary complex carbohydrates and prebiotics,which has been reported alleviate inflammation by regulating the proportions of certain intestinal bacteria to achieve anti-inflammatory effects and immunomodulation [36].The disorder of intestinal microbiota is associated with the prevalence of obesity and metabolic syndrome [2].In our work,RLPs improved SCFAs production,reduced the Firmicutes/Bacteroidetes ratio,and enhanced the relative abundances ofAkkermansiaand Bacteroidetes,which contribute to intestinal barrier function.An early report suggested that the anti-obesity mediated by producing more beneficial SCFAs,which were the pre-dominant end-products of increased complex carbohydrates fermented by gut-related bacterial [16].The previous research has highlighted the importance of SCFAs in promoting colonocyte repair and improving chronic inflammatory diseases [49].Besides,SCFAs regulate the production and function of nature and adaptive immune cells,and reducing the levels of SCFAs may cause an immune imbalance and thus immune-related pathologies [49].In addition,SCFAs may also participate in adjusting fatty acid synthesis,oxidation,and lipolysisin vivo,which affect the development of obesity [50].Kim et al.[51]reported that dietary fiber and resistantstarch can produce SCFAs during microbial fermentation in the colon,stimulate myeloid and lymphocyte proliferation,promote immunity,enhance intestinal barrier function,and then inhibit the invasion of harmful microorganisms into the host to reduce inflammatory diseases.Lately,studies have shown that SCFAs are the signal molecule of PPARγ[52],and the changes of dietary composition can easily lead to the changes of SCFAs content and then the activity of PPARs signaling pathway was changed.Previous studies have also reported that butyric acid can increase the expression of PPARs in adipocytes of pigs [53],thereby inhibiting diet-induced obesity by increasing energy consumption [54].Thus,the antiobesity effect of RLPs may be due to activation of PPARs signaling pathway,increasing SCFAs levels in the intestine,and improving the composition of intestinal bacteria to improve the abnormal lipid metabolism and inhibit the formation of obesity in rats with the HFD.
Propionic acid,the main fermentation product of Bacteroidetes,had an obvious inhibitory effect on fatty acid synthesis and cholesterol in the liver of mice [55].In addition,obese rats with an increase abundance of Firmicutes in the intestinal microbiota,improved the ability to obtain energy from the diet with more efficiently absorbing calories and subsequently gaining weight [56].It is universally acknowledged that a reduction in Firmicutes/Bacteroidetes ratio can avoid the development of obesity in both dietary and genetic models of obesity in rats [56].In this study,the HFD increased the abundance of Firmicutes and decreased the abundance of Bacteroidetes.Supplementation of RLPs in HFD dose-dependently increased the relative abundance of Bacteroidetes,and decreased the abundance of Firmicutes.
Our results suggested that RLPs might selectively adjust certain bacteria relating to inflammation and obesity.Dysbiosis of gut microbiota revealed a significant increase in Firmicutes including Clostridiaceae and Lachnoclostriaceae,whereas the levels of Bacteroidetes includingAlistipes,Butyricimonas,Paraprevotella,andAkkermansiabelonging to the phylum of Verrucomicrobia revealed a significant decrease.Clostridiaceae is the main reason of intestinal infection and diarrhea after antibiotic treatment [57],and the increase of Lachnospiraceae family level resulted in body weight gain in germ-free ob/ob mice [58].In patients with insulin resistance,Butyricimonasspecies showed a significant correlation with plasma glucose levels [59].Moreover,the increased relative levels ofAkkermansiamay play critical roles in preventing obesity,insulin resistance,and intestinal inflammation [60,61].A low proportion ofAkkermansiain the gut weakens the intestinal barrier,making it easier for intestinal toxins to invade the body,leading to thinning of the mucous membranes.Patients with inflammatory bowel disease,obesity,and type II diabetes mellitus have lowerAkkermansiarelative abundance [62].The results from our study showed that the RLPs increased the abundance ofButyricimonasandAkkermansia,and might prevent chronic inflammation.We also found thatPrevotellaandAlistipeswere down-regulated in the HFD group and up-regulated in the HRLPs group.PrevotellaandAlistipesare belonging to Bacteroidetes,which are generally regarded as “good bacteria” as they are negatively correlated with metabolic syndromes and diabetes [63-65].
The dietary functional components can directly affect gene expression,regulate post-metabolic transcription factor activity,or stimulate the signal transduction cascade to induce transcription [66].Nutrigenomics is containing transcriptome/microarray,proteomics,epigenetics,metabolomics,microbiome,and metagenomics,which is considered as the next era for food nutrition and human chronic disease prevention [67,68].
5.Conclusion
In this study,three omics methods were used to investigate the molecular mechanism of RLPs inhibiting inflammatory obesity.After diversity correlation analysis in different dimensions,we found that the intervention of RLPs may be conducive to the stability of some intestinal microbiota,particularly in a diet-induced microbial out of balance,and influence on corresponding metabolic pathways,contributing to keep host health.A further advanced investigation should be performed and more convincing molecular evidence should be obtained to reveal the comprehensive molecular mechanism of the biological function of RLPsin vitroandin vivo,before its clinical application shortly.
Conflicts of interest
The authors declare that there is no conflict of interest.

- 食品科学与人类健康(英文)的其它文章
- Colloidal nanoparticles prepared from zein and casein:interactions,characterizations and emerging food applications
- Biological factors controlling starch digestibility in human digestive system
- Preparation methods,biological activities,and potential applications of marine algae oligosaccharides: a review
- Development of hyaluronic acid-based edible film for alleviating dry mouth
- Mushroom β-glucan and polyphenol formulations as natural immunity boosters and balancers: nature of the application
- Preparation of multicore millimeter-sized spherical alginate capsules to specifically and sustainedly release fish oil