
基于转录组测序筛选鹿茸胸腺素促成骨细胞增殖基因
2023-02-25 03:20薛东明慧芳孙天霞姜英男赵雨
中国骨质疏松杂志 2023年1期
薛东明 慧芳 孙天霞 姜英男 赵雨
长春中医药大学人参科学研究院,吉林 长春130117
胸腺素是一种淋巴生成因子,由Goldstein等[1]在1966年从小牛胸腺组织中分离提取出来。它广泛存在于各个组织器官中,根据其等电点的不同分为3种[2-3](α、β和γ)。其中Tβ4是由43个氨基酸组成,分子质量为4921Da的β族胸腺素。Tβ4的主要功能是通过与G-actin结合调节肌动蛋白的聚合和解聚[4-8],在维持细胞骨架的动态平衡中起重要作用。以这个功能为基础,Tβ4在生物体中行使各种生物学功能,如促进血管生长、修复受损心肌细胞和创面、抗纤维化和神经保护作用等。
研究显示,Tβ4具有促进骨再生及骨生成的作用。将Tβ4应用于大鼠拔牙创面后,一些成骨相关细胞因子的表达增高,可观察到骨小梁增多[9]。除此之外,Tβ4可以通过上调成骨相关细胞因子的表达来修复大鼠颅骨缺损,促进骨折端骨痂的形成与矿化[10-11]。在鼠齿上皮细胞敲除Tβ4后发现,Tβ4可通过激活Smad与PI3K/Akt通路激活Runx2的上游启动子,增加Runx2的表达量,促进牙齿发育[12];另一方面,Tβ4可通过促进新生血管形成来调控骨再生和骨生成[13-14]。
根据前期实验对梅花鹿鹿茸初生期、快速生长期和骨化期转录组测序数据的分析结果,发现Tβ4基因在3个时期高度表达,根据该基因表达情况和鹿茸的快速生长特性,推测Tβ4基因对骨骼的生长也有着影响作用。
本研究为了系统评价梅花鹿胸腺素Tβ4在骨发育中的作用,体外合成梅花鹿胸腺素Tβ4蛋白。使用梅花鹿胸腺素Tβ4蛋白处理小鼠成骨细胞(MC3T3细胞)后进行转录组测序,筛选出骨发育相关基因和通路,为阐明梅花鹿胸腺素Tβ4蛋白对成骨细胞活性的影响及其机制提供理论依据。
1 材料与方法
1.1 材料
细胞增殖及细胞毒性检测试剂盒(武汉博士德生物,AR1160-500),MC3T3小鼠成骨细胞(由本实验室保存),梅花鹿源胸腺素Tβ4蛋白(生工生物公司合成),Trizol试剂盒(北京全式金生物,ER501-01),DMEM培养基(上海Vivacell公司,01-052-1ACS),牛血清(上海Vivacell公司,04-001-1ACS),双抗(上海Vivacell公司,03-031-1BCS),酶标仪(Infnite 200 PRO plate reader,Life Sciences,USA)。
1.2 方法
1.2.1梅花鹿胸腺素β4蛋白的合成及鉴定:本研究采用Fmoc固相合成法合成梅花鹿胸腺素β4蛋白,首先将肽链的C-端氨基酸的羟基通过共价键与Wang树脂相连,再进行脱氨基保护基反应,接长肽链。经过“缩合-洗涤-去保护-中和和洗涤-下一轮缩合”等步骤的重复,将肽链延伸至目标长度。最后,从Wang树脂上将目标肽链裂解下来,对纯化后得到的目的多肽进行鉴定。
1.2.2细胞活性检测:取对数生长期的MC3T3细胞,调整细胞悬液的浓度为5×104个/mL,将100 μL细胞悬液加入到96孔板中。培养4 h后,加入稀释好的Tβ4药液,对照组加入不含药物的等体积培养基。37 ℃,5% CO2培养48 h。弃培养基,加入90 μL培养基和10 μL CCK8试剂,反应25 min。用酶标仪在450 nm波长下测定吸光度,计算细胞存活率。
1.2.3mRNA文库构建:以8 μg total RNA起始量建库。利用mRNA转录组文库构建试剂盒(Hieff NGSTMMaxUp Dual-mode mRNA Library Prep Kit for Illumina®)中分离磁珠分离mRNA,并合成双链cDNA、末端补平、3’端加A、连接index接头。根据测得的片段大小,使用分选磁珠纯化文库扩增产物。对检测合格的文库利用Illumina平台进行高通量测序。
1.2.4基因表达分析及差异基因的筛选与富集:一个基因的表达水平反映了该基因转录本的丰度情况,本研究使用Salmon[15]计算基因的表达量。使用DESeq2(V1.6.3)[16]软件进行基因的差异分析,将获得的所有差异基因比对到GO和KEGG数据库,通过超几何检验进行GO和KEGG功能富集分析。
1.2.5实时荧光定量PCR分析:对提取的细胞总RNA进行反转录后,随机选择4个基因进行qRT-PCR以检验转录组数据的准确性,根据2-ΔΔCT法进行相对定量分析,引物序列见表1。
1.2.6蛋白互作网络(PPI)构建及Hub基因筛选和富集:使用 STRING 数据库(https://cn.string-db.org/)预测差异基因的PPI网络,分析蛋白质之间的功能性相互作用有助于阐明梅花鹿Tβ4促成骨细胞增殖的机制。导出 STRING 数据库构建的差异基因PPI网络至Cytoscape软件,使用cytohubba和BiNGO插件筛选中枢基因(hubgene,Hub)并对其进行GO富集分析。Hub基因的KEGG富集分析使用了在线软件DAVID(https://david.ncifcrf.gov/)。
2 结果
2.1 合成多肽的质谱鉴定分析
本研究采用LC-MS对多肽进行质谱分析,结果见图1。可以看出合成多肽的分子量为4 964.40,与梅花鹿胸腺素β4的理论分子量4 963.426一致,证明合成多肽即为目的产物。
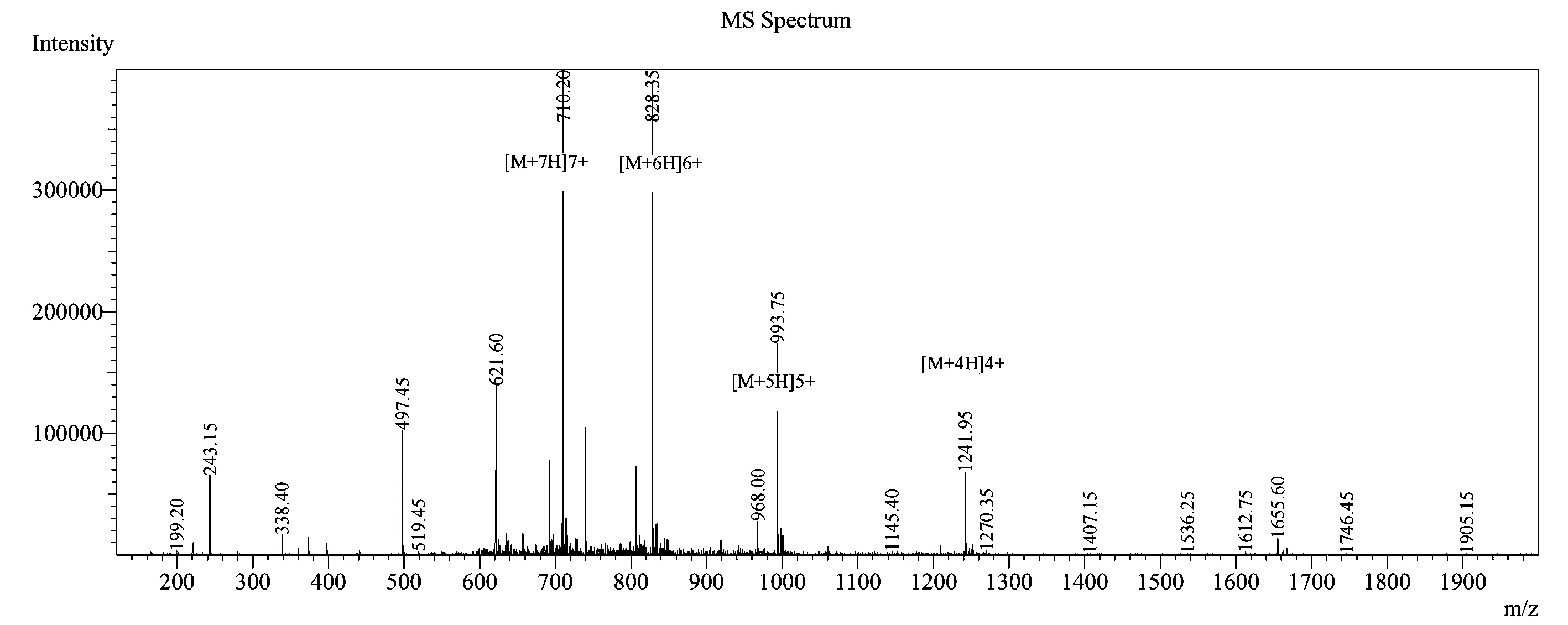
图1 合成Tβ4蛋白质谱鉴定图Fig.1 Synthetic Tβ4 protein mass spectrum identification diagram
2.2 细胞活性检测
使用不同浓度Tβ4蛋白处理成骨细胞后48 h,检测各组细胞的活性。如图2所示,Tβ4蛋白处理后的成骨细胞存活率明显高于对照组细胞,且差异具有统计学意义。在使用浓度为150 μg/mL的Tβ4蛋白处理细胞时,促进细胞生长的效果最明显(P<0.001)。说明Tβ4蛋白处理成骨细胞后显著促进了细胞的增殖活性(表2)。
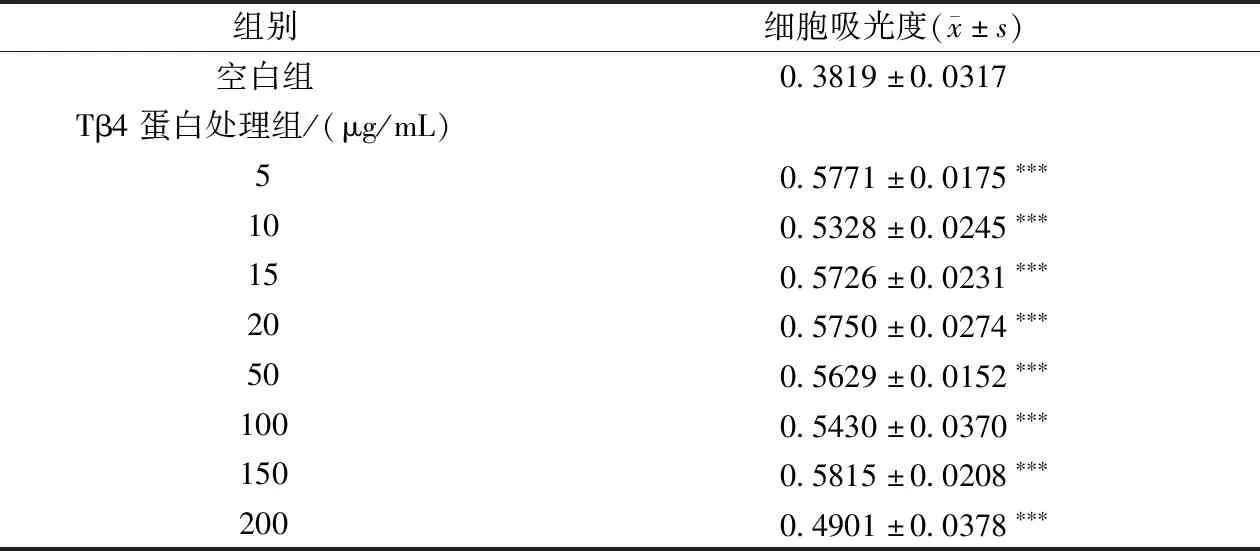
表2 不同浓度Tβ4蛋白处理成骨细胞48 h后细胞吸光度Table 2 Cell absorbance of osteoblasts treated with different concentrations of Tβ4 liquid for 48 h
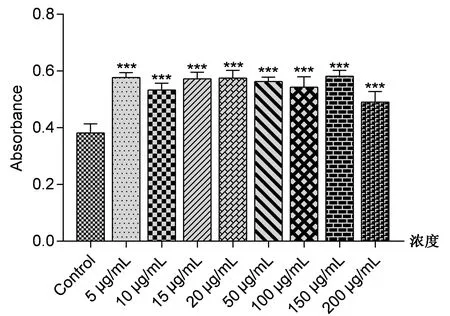
图2 不同浓度Tβ4药液对成骨细胞存活率的影响Fig.2 Effects of different concentrations of Tβ4 liquid on the survival rate of osteoblasts
2.3 qRT-PCR验证
为检验转录组数据是否准确、可信,随机选取4个差异基因进行qRT-PCR检测。结果表明qPCR测定结果与转录组测序结果得到的差异基因表达变化趋势一致(图3),表明本研究中转录组测序得到的数据是可信的。
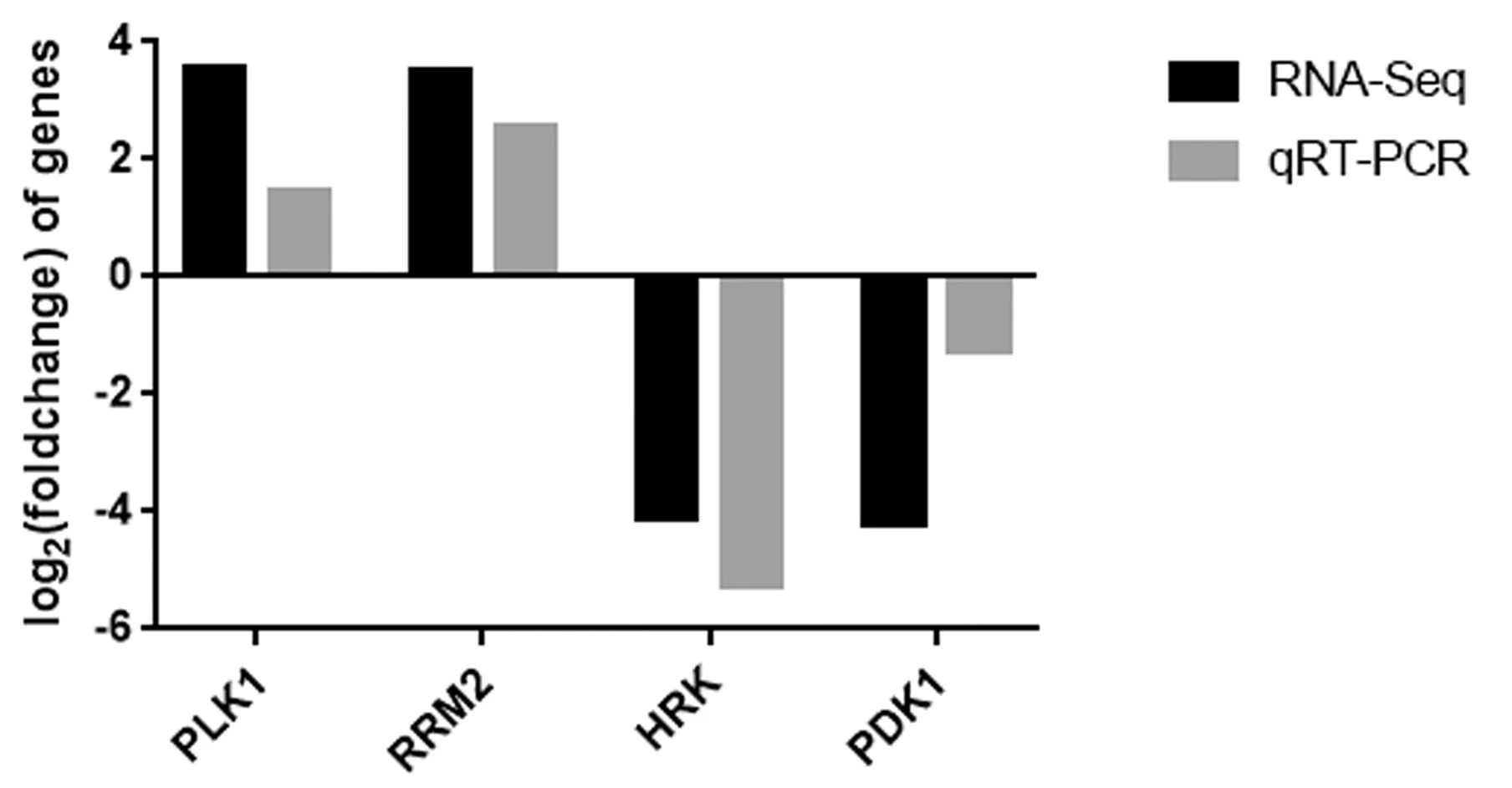
图3 转录组测序结果的qRT-PCR验证Fig.3 qRT-PCR verification of transcriptome sequencing results
2.4 样本间相关性及差异基因富集分析
为了后续的差异基因分析对各组进行了相关性分析,根据图4可以看到组内及组间样品的相关性,两组样本的组内皮尔森系数较高,组内重复性好。两组样品的组间皮尔森相关系数为0.932 1~0.889 1,表明两组样品间存在差异,可以用于后续差异基因的分析。
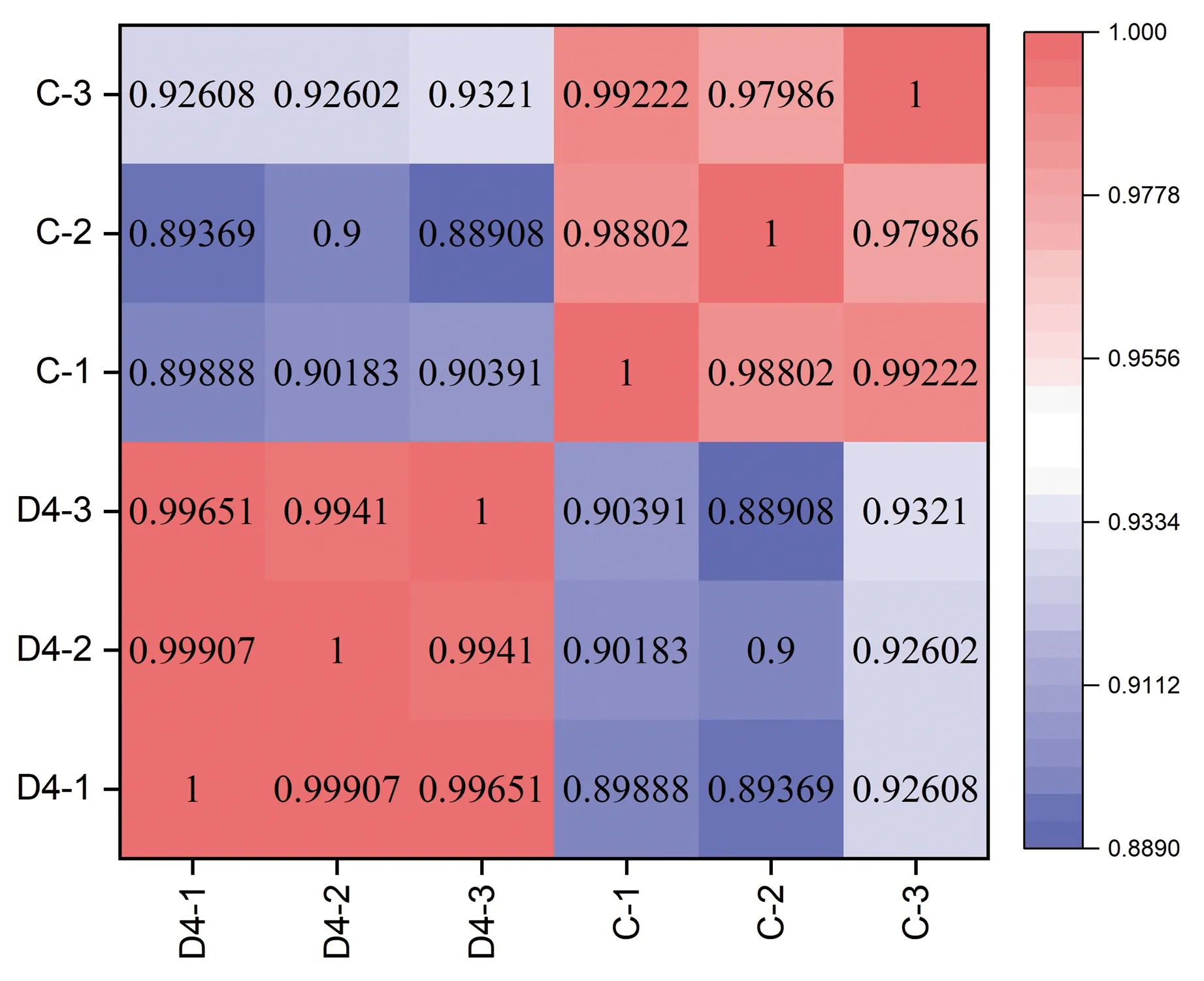
图4 样本间相关性分析热图Fig.4 The heat map of inter-sample correlation analysis
本研究以P<0.05且|Log2FC|≥2为筛选条件,以差异基因的P值及Log2FC值为输入数据,绘制火山图,Tβ4处理组和对照组的差异基因筛选结果如图5,相对于对照组,Tβ4处理组共获得差异基因6 106个,其中1 979个基因为上调,4 127个基因为下调。
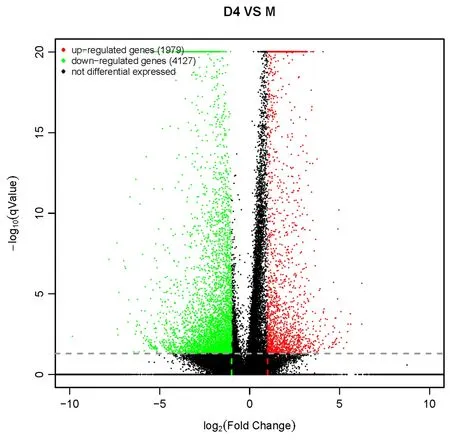
图5 差异基因火山图Fig.5 Volcano map of differential genes
对差异基因的GO和KEGG富集分析结果以校正后的P<0.05作为阈值,满足此条件的GO条目及KEGG通路存在显著富集的情况,选择富集程度最高的前30个GO条目及KEGG通路进行绘图。富集结果如图6,可以看出富集程度较大的GO功能包括细胞因子产生的调节、肽基-酪氨酸磷酸化的正调控和蛋白酪氨酸激酶活性的正调控等。
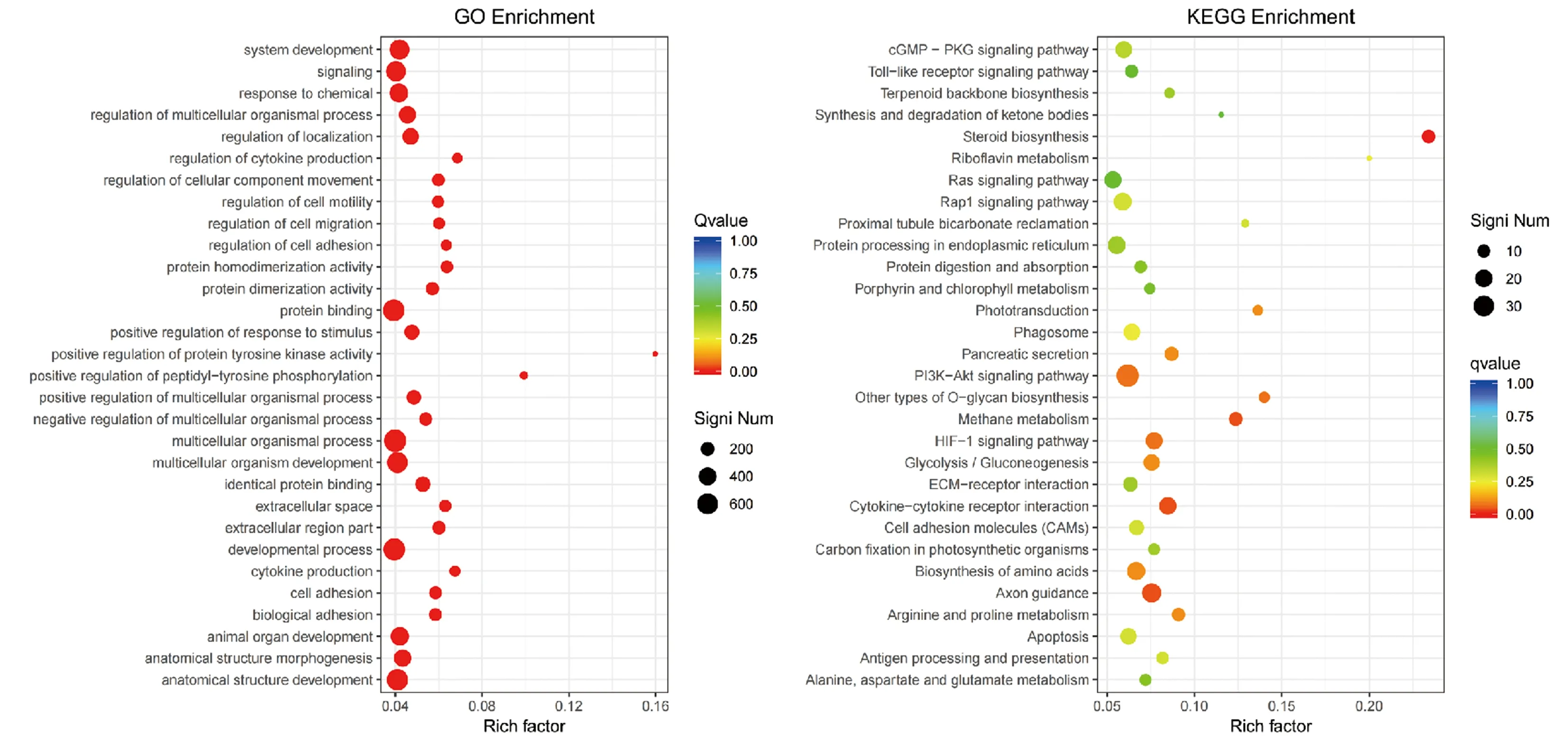
图6 差异基因GO及KEGG功能富集散点图Fig.6 Scatter plot of functional enrichment of differential genes go and KEGG注:纵轴为功能名称,横轴为Rich Factor,Rich factor的值越大,则富集的程度越大,图中点的大小则表示富集到的差异基因个数,点越大,富集到的差异基因越多。
差异基因的KEGG富集中富集程度较大的信号通路包括甾体生物合成、核黄素代谢和萜类骨架生物合成等。
2.5 促成骨细胞增殖候选基因及通路的筛选
将差异基因筛选条件设置为|Log2FC|≥2,Q值<0.001且表达量>5,继续筛选促进成骨细胞增殖候选基因,共获得60个差异基因,其中39个基因显著上调,21个基因显著下调。
利用STRING数据库构建60个差异基因的蛋白互作网络,节点数为59,边数为414;平均节点度为14,平均局部聚类系数为0.709。
使用Cytoscape软件中的cytohubba插件进一步筛选Hub基因,差异基因中前10位的Hub基因包括CDK1、PLK1、CDC20、BUB1B、BUB1、RRM2、MAD2L1、NCAPG、NCAPH、FEN1(表3)。
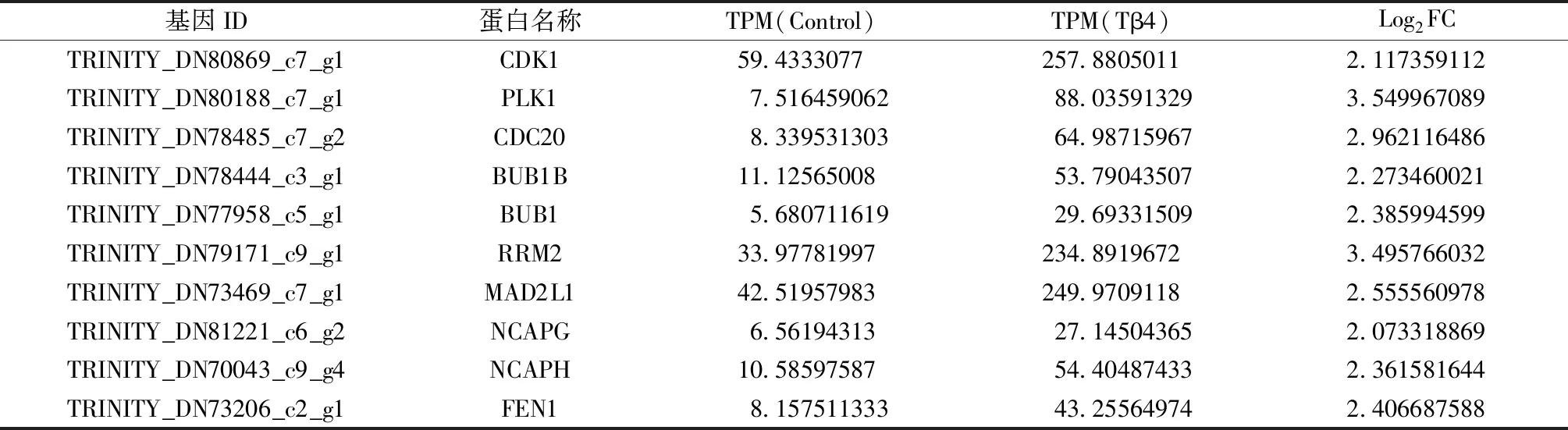
表3 Hub基因表达量Table 3 The expression level of Hub gene
采用Cytoscape软件中的BINGO插件对筛选到的Hub基因进行GO富集,构建GO富集的生物学过程有向无环图。结果显示,富集程度较高的GO条目为:细胞周期的正调控、有丝分裂细胞周期的调节、细胞分解代谢过程的正向调节、细胞周期过程的调节和细胞周期过程的正调控等。
通过在线软件DAVID对Hub 基因进行KEGG通路分析,结果发现有7个候选基因富集到了细胞周期和p53信号通路中(表4)。分别为CDC20(细胞分裂周期分子20)、PLK1(丝氨酸/苏氨酸蛋白激酶)、CDK1(细胞周期蛋白依赖性激酶1)、BUB1B(有丝分裂检查点丝氨酸/苏氨酸蛋白激酶BUB1β)、BUB1(有丝分裂检查点丝氨酸/苏氨酸蛋白激酶BUB1)、MAD2L1(有丝分裂纺锤体组装检查点蛋白MAD2A)、RRM2(核糖核苷二磷酸还原酶亚基M2)。

表4 Hub基因KEGG富集分析Table 4 KEGG enrichment analysis of Hub genes
3 讨论
细胞可以通过完成细胞周期来产生新的细胞,实现细胞增殖[17-18]。细胞周期是一系列严格调控的分子事件,控制DNA复制和有丝分裂,对调控细胞增殖和生长具有重要作用。
为了研究梅花鹿胸腺素Tβ4对骨发育的作用,本研究使用体外合成的梅花鹿Tβ4蛋白处理MC3T3细胞后进行转录组测序,筛选出7个促进成骨细胞增殖的候选基因。其中,CDK1、BUB1B、BUB1和PLK1都是细胞周期检查点的重要调控基因[19],可促进细胞增殖。研究表明,CDK1通过结合周期蛋白A/B形成A/B-CDK1来促进通过G2进入M期[20]。BUB1和BUBR1是纺锤体组装有丝分裂检查点(SAC)的核心组成部分[21],同时也是PLK1的受体。PLK1在真核细胞内表达,在有丝分裂、细胞的增殖和细胞周期的调控过程中有重要作用[22-23],与CDK1功能相似,可促进BUB1和BUB1B磷酸化[24],并与之结合,发挥SAC的功能[25]。PLK1的敲除会使细胞周期滞留在G2/M期[26],因此PLK1在SAC中是不可或缺的基因。RRM2可以调控核糖核苷酸还原酶的活性,在细胞增殖及DNA合成和修复过程中起到重要作用[27]。
除促增殖作用外,CDK1还可与细胞周期调节因子 B1(cyclin B1)结合来调节线粒体p53的抗凋亡作用[28]。以上信息说明梅花鹿胸腺素Tβ4可能通过这些基因调控细胞周期信号通路和p53信号通路来促进成骨细胞增殖。
目前,对于梅花鹿胸腺素Tβ4研究较少,因此本研究以高通量测序结果为基本,基于生物信息学的预测,筛选出了促成骨细胞增殖的候选基因。根据结果预测胸腺素Tβ4有抗凋亡以及加速细胞周期促进增殖的作用,为研究成骨细胞增殖机制提供了方向。在后续工作中,需要对候选基因进行功能验证及分析,为今后研究胸腺素对骨发育的影响提供理论和实验基础。
猜你喜欢
天津医科大学学报(2021年3期)2021-07-21
幼儿100(2021年15期)2021-05-26
心电与循环(2020年1期)2020-02-27
作文小学中年级(2019年9期)2019-10-14
作文·小学低年级(2019年9期)2019-09-10
江苏农业科学(2017年5期)2017-04-15
中国民族医药杂志(2016年6期)2016-05-09
中国医科大学学报(2015年10期)2015-03-01
湖北农业科学(2014年3期)2014-07-21
中国医学科学院学报(2011年5期)2011-12-01
