
干旱胁迫下蒭雷草的生理响应
2023-03-09 02:13陈意兰刘东明王俊简曙光王发国
热带亚热带植物学报 2023年1期
陈意兰, 刘东明*, 王俊, 简曙光, 王发国
干旱胁迫下蒭雷草的生理响应
陈意兰1,2, 刘东明1,2*, 王俊1,2, 简曙光1,2, 王发国1,2
(1. 中国科学院华南植物园,中国科学院海岛与海岸带生态修复工程实验室,中国科学院南海生态环境工程创新研究院,广州 510650; 2. 华南国家植物园, 广州 510650)
为探讨蒭雷草()在热带珊瑚岛干旱环境下的适应能力,对干旱胁迫下蒭雷草叶片抗逆生理指标的变化进行了研究。结果表明,干旱胁迫初期,叶片的丙二醛(MDA)含量随胁迫程度增加的差异不显著;随胁迫时间的延长,除重度胁迫下先迅速增加后急速下降外,其余处理的变化较小。超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性随干旱胁迫程度增加而升高;随时间延长,SOD活性不断上升,POD活性基本稳定,CAT活性则先下降后上升。除轻度胁迫下可溶性蛋白(SP)含量低于对照外,其余处理均随干旱胁迫程度增加而增加;干旱胁迫下的脯氨酸(Pro)含量随时间延长呈先上升后下降的变化趋势,但在处理第18天时不同胁迫程度间均差异不显著。因此,蒭雷草具有较强的抗旱能力,可用于南海诸岛的人工植物群落构建和植被恢复以营造良好生态环境。
蒭雷草;热带珊瑚岛;干旱胁迫;生理响应
蒭雷草()是我国热带珊瑚岛常见禾本科(Gramineae)多年生草本植物,对南海诸岛防风固沙、营造良好生态环境等具有重要作用[1]。热带珊瑚岛属于典型的热带和赤道带海洋季风气候;日照时间长,辐射强烈,热量充足,终年高温,季风盛行;土壤呈强碱性,盐分含量较高;自然环境恶劣,植被生态系统非常脆弱;生长在该区域的植物应具有耐盐、耐高温、耐旱、喜钙等特殊功能性状以适应生境[2–3]。恢复南沙岛礁的植被生态系统,需要筛选出耐旱性强的植物并进行引种[4]。目前,有关蒭雷草的研究主要集中在花的结构发育[5]、珊瑚礁沙土栽培保育[6]、叶片水势和渗透溶质与环境间的关系[7]等方面,对其抗逆生理生化响应等为适应逆境的研究较少,缺乏支撑其具有适应热带珊瑚岛生境的依据。因此,研究蒭雷草对热带珊瑚岛生境干旱的适应能力,揭示其耐旱机理,对南沙诸岛植被恢复引种具有重要意义。
丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、可溶性蛋白(SP)和脯氨酸(Pro)在植物代谢活动中起着重要作用,是衡量植物抗逆性强弱的重要生理指标[8–10]。通过膜脂过氧化产物MDA含量、抗氧化酶SOD、POD、CAT活性和渗透调节物质SP、Pro含量分析植物耐旱性的研究报道很多,如考来木()[11]、粗壮女贞()[12]、绣球(sp.)[13]等。本文为探讨蒭雷草在热带珊瑚岛高温、季节性干旱环境下的适应能力,揭示其耐旱机理,从西沙东岛采集蒭雷草,通过分株繁殖, 利用梯度控水控制土壤湿度,模拟热带珊瑚岛生境, 研究干旱胁迫下蒭雷草叶片的抗逆相关指标的生理响应,以期为蒭雷草在南沙诸岛人工群落构建、植被恢复应用和营造良好生态环境研究提供参考。
1 材料和方法
1.1 研究地点概况
西沙东岛位于海南省三沙市西沙群岛的东部, 位于112°43′13″~112°44′22″ E,16°39′34″~16°40′31″ N,陆地面积1.7 km2,呈椭圆形,是西沙群岛三大珊瑚岛之一[14–15],面积超过1.5 km2,年均温26 ℃~ 27 ℃,年降雨量1 500 mm,雨季集中在6—11月,5—10月和12月干旱少雨,干湿季分明,季节性干旱现象明显,土壤为富含钙和磷质的石灰土和滨海盐,保水能力差[16–17]。
1.2 方法
2018年4月在西沙东岛采集蒭雷草()鲜活植株带回广东省广州市中国科学院华南植物园实验大棚内,在30 cm×25 cm的花盆内进行分株繁殖,基质为珊瑚砂:红壤土:椰糠:泥炭土:有机肥=80:10:5:3:2。2019年7月,选取48株长势一致的健壮植株进行干旱胁迫试验,2株/盆, 共24盆。随机分成4组,每组6盆。以控制土壤含水量设置4个干旱胁迫程度(表1):CK (对照)土壤含水量40%~50%,正常供水;S1土壤含水量30%~40%,轻度胁迫;S2土壤含水量20%~30%,中度胁迫; S3土壤含水量小于20%,重度胁迫。每天用Delta-T便携式土壤水分仪测定土壤含水量,并浇水50~200 mL以控制土壤含水量。定期在各组内随机调换植株位置,尽量保证各组植株除水分外其他生长条件一致,减少试验误差。试验开始后每隔3 d采摘成熟叶片(第3~8位功能叶),带回实验室进行生理指标测定,每样品测量3次,取平均值,共采样7次。
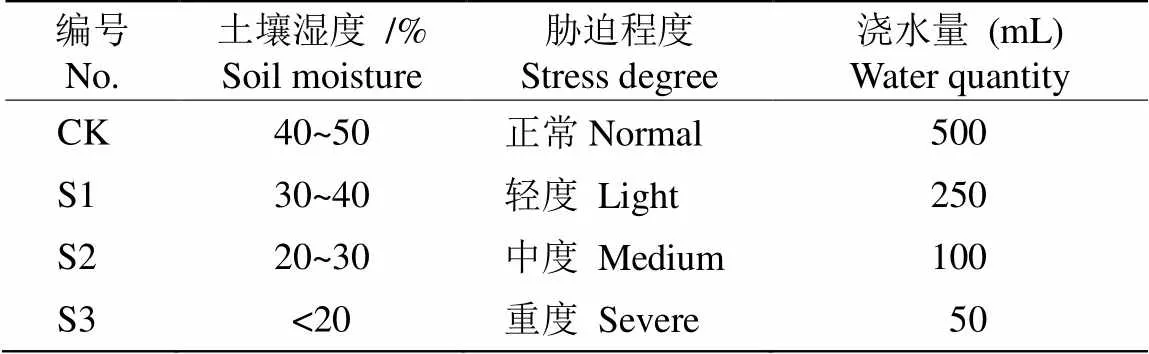
表1 盆栽苗土壤体积含水量
1.3 测定指标及方法
丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法[18]测定;超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)光化还原法[19]测定,过氧化氢酶(CAT)活性采用紫外吸收法[20]测定,过氧化物酶(POD)活性采用愈创木酚显色法[21]测定,可溶性蛋白(SP)含量采用二喹啉甲酸(BCA)法[22]测定,脯氨酸(Pro)含量采用酸性茚三酮染色法[21]测定。
1.4 数据的统计分析
采用Excel 2010对数据进行整理,用SPSS Stati- stics 17软件对数据进行统计分析。统计分析前采用Kolmogorov Smirnov正态性检验和Levene’s方差齐性检验进行评估,当数据不符合正态性和方差齐性假设时,对其进行对数变换。采用单因素方差分析(One- Way ANOVA)和最小显著差异法(LSD)分析干旱胁迫对生理指标的影响,处理时间和胁迫程度对生理指标的影响采用双因素方差分析,同时用重复测量方差分析进行检验,采用Duncan’s法在<0.05水平上进行多重比较。利用Excel 2010和Photoshop绘图。
2 结果和分析
2.1 丙二醛(MDA)含量的变化
从图1: A可见,干旱胁迫初期(0~3 d),随胁迫程度增加MDA含量的差异不显著;干旱胁迫12 d,S3处理的MDA含量达到最高,为66.99 mmol/g, 比CK上升了166.12%,达差异显著水平,其余处理的MDA含量变化较小。这表明干旱胁迫初期蒭雷草叶片还未发生膜脂过氧化损伤,MDA未积累; 干旱胁迫中后期(6~18 d),MDA含量呈上升-下降-上升的变化趋势,表明蒭雷草叶片细胞开始积累MDA,且相同处理时间下随胁迫程度增加MDA含量增加,在中、轻度胁迫下MDA含量变化较小。
2.2 抗氧化酶活性的变化
SOD活性 蒭雷草叶片的SOD活性随干旱胁迫时间延长呈逐渐上升趋势;相同处理时间SOD活性以S3>S2>S1>CK。CK的SOD活性变化较小, S1处理的呈上升-下降-上升的变化趋势,干旱胁迫第9天达到最高,为270.94 U/g,比CK上升了42.72%, 差异显著;S2、S3处理的呈逐渐上升的变化趋势(图1: B)。
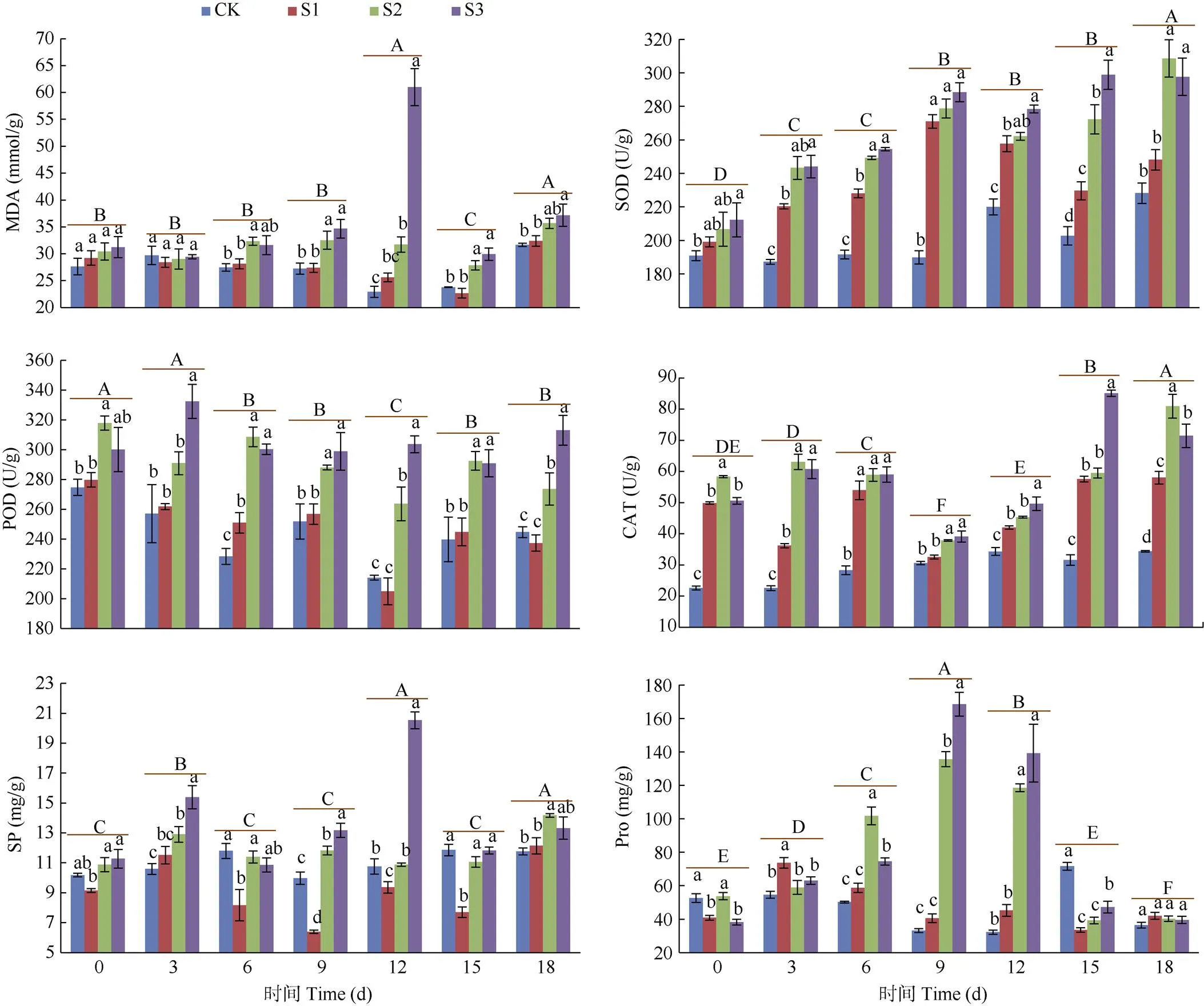
图1 干旱胁迫下叶片的MDA、SP、Pro含量和SOD、POD、CAT活性的变化。CK: 对照; S1: 轻度干旱; S2: 中度干旱; S3: 重度干旱。柱上不同小写字母表示相同时间不同处理间差异显著(P<0.05),不同大写字母表示相同处理不同时间差异显著(P<0.05)。
POD活性 从图1: C可见,POD活性总体上随处理时间的延长变化较小。相同处理时间,除第6天S1与CK的POD活性差异显著外,其余处理时间差异不显著;S3与S2处理除第3天、第12天和第18天的差异显著外,其余均不显著。S3处理第3天的POD活性迅速上升,表明重度干旱胁迫下蒭雷草能迅速反应,提高POD活性以抵抗胁迫伤害。
CAT活性 从图1: D可见,CK的CAT活性随时间延长缓慢上升,干旱胁迫处理的CAT活性变化规律不一致。S1、S2、S3处理第9天的CAT活性均下降至最低, 分别为32.54、37.86和39.15 U/g,分别比CK上升了6.16%、23.52%和27.75%,随后又上升;S1、S2、S3处理第18天的CAT活性分别为58.03、80.94和71.48 U/g,分别比CK上升了68.78%、135.41%和107.87%,均差异显著。总体来看,干旱胁迫下CAT活性呈先下降后上升的变化趋势,说明胁迫初期CAT活性变化不大,随着胁迫时间增加,CAT活性上升。
2.3 渗透调节物质的变化
SP含量 从图1: E可见,S1处理相同时间,除第3和18天SP含量比CK的高,但差异不显著外,其余时间均低于CK。S2和S3处理的SP含量高于CK或差异不显著,且随干旱胁迫程度的加重而增加。总体来看,SP含量随处理时间的延长变化较大,但SP含量最低时仍与初始时差异不显著, 这说明干旱胁迫下蒭雷草通过维持SP水平以保证细胞内外渗透势的平衡。
Pro含量 从图1: F可见,S1处理除第3天Pro含量高于CK且差异显著外,其余处理时间低于CK或差异不显著。S2和S3处理第9天Pro含量达到最大,分别为135.66和168.53 mg/g,分别比CK上升了309.90%和409.19%,均差异显著; 干旱胁迫第18天,Pro含量分别下降至40.22和39.48 mg/g, 与CK的差异不显著。总体来看,Pro含量随处理时间延长呈先上升后下降的变化趋势, 干旱胁迫第18天各组Pro含量均差异不显著,说明干旱胁迫下蒭雷草能维持Pro含量以保证细胞内外渗透势的平衡。
2.4 方差分析统计检验
从表2可见,处理时间对蒭雷草叶片的MDA和SP含量、POD和CAT活性有显著影响(<0.05),但对SOD活性、Pro含量没有显著影响(0.05); 胁迫程度对蒭雷草叶片的MDA、SP和Pro含量、SOD、POD和CAT活性均有显著影响(<0.05);处理时间和胁迫程度对这些指标产生显著的交互作用(<0.05)。

表2 处理时间和胁迫程度对生理指标的方差分析
3 讨论和结论
3.1 干旱胁迫对叶片MDA含量的影响
持续干旱致使植物体内的活性氧含量增加,而活性氧的积累会引起膜脂过氧化作用[23]。MDA是膜脂过氧化产物之一,其含量反映膜脂过氧化损伤的程度[24]。本研究中,干旱胁迫前期,不同胁迫程度蒭雷草叶片的MDA含量差异不显著, 说明短期胁迫处理未造成蒭雷草发生膜脂过氧化损伤;随处理时间延长,除重度胁迫外,MDA含量的变化较小,说明蒭雷草对干旱胁迫表现出较强的防御能力,将MDA含量维持在较低水平, 减缓干旱导致的伤害[25]。这与热带珊瑚岛的长春花()[26]、莲叶桐()[27]一样,对干旱、强光、盐碱等环境具有良好的适应性。
3.2 干旱胁迫对叶片抗氧化酶活性的影响
干旱胁迫下叶片细胞内活性氧积累,导致发生膜脂过氧化损伤,抗氧化酶活性的增强有利于清除细胞内的活性氧[28]。SOD可使超氧阴离子发生歧化反应生成H2O2,POD、CAT再将H2O2转化为H2O,从而减缓膜系统的损伤[29]。本研究中,蒭雷草叶片SOD活性随胁迫时间延长持续上升,超氧阴离子不断被清除,膜脂过氧化作用减缓[30],使MDA含量在一定时期内维持在较低水平。POD活性保持较低水平,与李捷等[31]的研究结果不一致,可能与蒭雷草叶片细胞启动合成POD的机制不敏感有关。CAT活性在干旱胁迫前期呈下降趋势,导致重度胁迫第12天的活性氧没有迅速清除,MDA含量达到最高;随后CAT活性持续上升,与其他抗氧化酶协同作用清除活性氧,减缓膜脂过氧化损伤,MDA含量又迅速下降[29],之后虽有上升,但上升速率较缓。有研究表明,植株通过提高抗氧化酶活性来提高抵御干旱的能力,当超过一定阈值后,抗氧化酶活性会逐渐下降[32]。CAT活性先降低后增加,与单叶蔓荆(var.)[33]受到干旱胁迫时的变化不一致,可能是干旱胁迫前期植株启动合成CAT的机制不敏感,CAT没有积累,干旱胁迫后期蒭雷草叶片细胞机体迅速作出反应,CAT活性上升[34]。
3.3 干旱胁迫对叶片渗透调节物质的影响
干旱胁迫下,植物通过提高SP和Pro含量以降低细胞渗透势,使细胞膨压稳定,以维持正常的生理反应,保证细胞结构与功能正常[35]。本研究中, 轻度胁迫下SP、Pro含量变化不大,可能是因为轻度干旱胁迫没有造成细胞渗透势的变化,细胞内外水分仍处于动态平衡。随着干旱胁迫程度加重,SP、Pro含量不断升高以增加细胞渗透势,从而保持细胞水分不流失,将细胞内外水分保持动态平衡[36]。有研究表明,植物细胞的SP不只参与调节细胞渗透势,还参与各种代谢[37]。本研究中,轻度胁迫的SP含量显著低于CK,中度胁迫的SP含量变化较小,重度胁迫的SP含量显著高于CK,说明当干旱胁迫对细胞没有造成威胁时,SP既参与调节细胞渗透势,也参与各种代谢;而干旱胁迫对细胞造成威胁时,细胞积累的SP主要参与调节细胞渗透势, 从而提高植物细胞保水能力,这与李鸿博等[38]的研究结果一致。
综上,为清除干旱胁迫产生的活性氧,蒭雷草会迅速开启防御系统,激活抗氧化酶。轻度干旱胁迫下,蒭雷草叶片细胞的SP、Pro含量变化不大, 随胁迫程度加重,SP、Pro不断积累以提高细胞渗透势,维持细胞内外水分平衡。因此,在抗氧化酶及渗透调节物质的保护下,蒭雷草表现出了较强的防御能力,具有较强抗旱能力,可引种到南海诸岛进行植被生态系统构建与恢复。
[1] JIAN S G. Vegetation of tropical coral islands in China [J]. Guihaia, 2020, 40(3): 443. [简曙光. 中国热带珊瑚岛植被 [J]. 广西植物, 2020, 40(3): 443.]
[2] HUANG Z G, ZHANG W Q. A discussion on the quaternary climate record from the coral reef in tropical China [J]. Trop Geogr, 2008, 28(1): 11–15. [黄镇国, 张伟强. 中国热带珊瑚礁的第四纪气候记录 [J]. 热带地理, 2008, 28(1): 11–15. doi: 10.3969/j.issn.1001-5221.2008.01.003.]
[3] XING F W, DENG S W. Flora of the South China Sea Island [M]. Beijing: China Forestry Press, 2019. [邢福武, 邓双文. 中国南海诸岛植物志 [M]. 北京: 中国林业出版社, 2019.]
[4] LI X Y, LIU D M, JIAN S G, et al. Biological characteristics of drought resistance of[J]. Guihaia, 2021, 41(6): 914–921. [李晓盈, 刘东明, 简曙光, 等. 海岸桐的抗旱生物学特性 [J]. 广西植物, 2021, 41(6): 914–921. doi: 10.11931/hiuhaia.gxzw201908017.]
[5] KUOH C S, HSIAO L C, LIAO G I. Comparison of upper floret development in bisexual and male spikelets of(Gramineae) with scanning electron microscopy [J]. Taiwania, 1998, 43(3): 235–245.
[6] WANG F, JIAN S, LIU D, et al. Constructing coral reef shelter by leveling coral reef sand, planting trees comprising arbor, shrubs, grass vine and herbaceous plant on coral reef sands, and performing conser- vation process: CN, 108605572-A; CN 108605572-B [P]. 2018.
[7] ALLAWAY W G, PITMAN M G, STOREY R, et al. Water relations of coral cay vegetation on the great barrier reef: Water potentials and osmotic content [J]. Aust J Bot, 1984, 32(4): 449–464. doi: 10.1071/B T9840449.
[8] LIANG F, TAN X H, DENG X, et al. Growth and physiological responses of semi-mangrove plantto water- logging and salinity stress [J]. Guihaia, 2021, 41(6): 872–882. [梁芳, 檀小辉, 邓旭, 等. 半红树植物玉蕊对淹水-盐度胁迫的生长及生理响应 [J]. 广西植物, 2021, 41(6): 872–882. doi: 10.11931/guihaia. gxzw202007036.]
[9] LIU Z B, CHENG R M, XIAO W F, et al. Growth and physiological responses ofseedlings to autumn and winter flooding [J]. J Lake Sci, 2016, 28(2): 405–413. [刘泽彬, 程瑞梅, 肖文发, 等. 中华蚊母树()幼苗对秋、冬季淹水的生长及生理响应 [J]. 湖泊科学, 2016, 28(2): 405–413. doi: 10.18307/ 2016.0221.]
[10] TANG D, CHENG P, YANG J J, et al. Physiological responses of plants to drought stress in the northern piedmont, Tianshan Mountains [J]. Arid Zone Res, 2021, 38(6): 1683–1694. [汤东, 程平, 杨建军, 等. 天山北坡山前植物对干旱胁迫的生理响应 [J]. 干旱区研究, 2021, 38(6): 1683–1694. doi: 10.13866/j.azr.2021.06.20.]
[11] ZHANG Z L, LIU Z H, CHEN M H, et al. Physiology response ofseedlings to combined elevated temperature and drought stress [J]. N Hort, 2017(19): 81–88. [张志录, 刘中华, 陈明辉, 等. 高温干旱胁迫下考来木幼苗的生理响应 [J]. 北方园艺, 2017(19): 81–88. doi: 10.11937/bfyy.20170134.]
[12] WU L L, WANG D L, LI Z Y. Physiological response ofto continuous drought and rewatering [J]. Plant Physiol J, 2018, 54(5): 837–844. [吴丽丽, 王德炉, 李自玉. 粗壮女贞对持续性干旱及复水的生理响应 [J]. 植物生理学报, 2018, 54(5): 837–844. doi: 10.13592/j.cnki.ppj.2017.0570.]
[13] REN Q Q, SUN J X, ZHANG D S, et al. Physiological response and drought resistance evaluation of differentunder drought stress [J]. Acta Agric Univ Zhejiang, 2021, 33(10): 1852–1860. [任倩倩, 孙纪霞, 张德顺, 等. 干旱胁迫下不同绣球品种生理响应与抗旱性评价 [J]. 浙江农业学报, 2021, 33(10): 1852–1860. doi: 10. 3969/j.issn.1004-1524.2021.10.08.]
[14] TONG Y, JIAN S G, CHEN Q, et al. Vascular plant diversity of the Paracel Islands, China [J]. Biodiv Sci, 2013, 21(3): 364–374. [童毅, 简曙光, 陈权, 等. 中国西沙群岛植物多样性 [J]. 生物多样性, 2013, 21(3): 364–374. doi: 10.3724/SP.J.1003.2013.11222.]
[15] WANG S H, ZHU Y J, WANG Y F, et al. Effect of vegetation types on soil physicochemical property in East Island and Yongxing Island of Xisha Islands [J]. J Trop Subtrop Bot, 2019, 27(4): 383–390. [王森浩, 朱怡静, 王玉芳, 等. 西沙群岛主要岛屿不同植被类型对土壤理化性质的影响 [J]. 热带亚热带植物学报, 2019, 27(4): 383–390. doi: 10.11926/jtsb.4013.]
[16] LIU X T, GE C D, ZOU X Q, et al. Carbon, Nitrogen geochemical characteristics and their implications on environmental change in the lagoon sediments of the Dongdao Island of Xisha Islands in South China Sea [J]. Acta Oceanol Sin, 2017, 39(6): 43–54. [刘晓瞳, 葛晨东,邹欣庆, 等. 西沙群岛东岛潟湖沉积物碳、氮元素地球化学特征及其指示的环境变化 [J]. 海洋学报, 2017, 39(6): 43–54. doi: 10.3969/j. issn.0253-4193.2017.06.005.]
[17] XING F W, WU D L, LI Z X, et al. Investigation of plant resources in Xisha Islands [J]. J Plant Resour Environ, 1993, 2(3): 1–6. [邢福武, 吴德邻, 李泽贤, 等. 西沙群岛植物资源调查 [J]. 植物资源与环境, 1993, 2(3): 1–6.]
[18] WANG X K, HUANG J L. Principles and Techniques of Plant Physio- logical and Biochemical Experiments [M]. 3rd ed. Beijing: Higher Education Press, 2015: 131–133. [王学奎, 黄见良. 植物生理生化实验原理和技术 [M]. 第3版. 北京: 高等教育出版社, 2015: 131–133.]
[19] CHEN J X, WANG X F. Experimental Guidance of Plant Physiology [M]. Guangzhou: South China University of Technology Press, 2002. [陈建勋, 王晓峰. 植物生理学实验指导 [M]. 广州: 华南理工大学出版社, 2002.]
[20]LU W J, LI Y S. Experimental Course of Plant Physiology [M]. Beijing: China Forestry Press, 2012. [路文静, 李奕松. 植物生理学实验教程 [M]. 北京: 中国林业出版社, 2012.]
[21] LI H S. Principles and Techniques of Plant Physiological Biochemical Experiment [M]. Beijing: Higher Education Press, 2000. [李合生. 植物生理生化实验原理和技术 [M]. 北京: 高等教育出版社, 2000.]
[22] WU S H, CHEN H W, JIAN S G, et al. The biological characteristics ofon tropical coral island in China [J]. Ecol Sci, 2017, 36(6): 57–63. [吴淑华, 陈昊雯, 简曙光, 等. 中国热带珊瑚岛橙花破布木()的生物学特性 [J]. 生态科学, 2017, 36(6): 57–63. doi: 10.14108/j.cnki.1008-8873.2017.06.008.]
[23]CHEN J. Response of activated oxygen metabolism to drought stress of four ornamental species [D]. Hangzhou: Zhejiang Agric For University, 2013. [陈珺. 四种观赏植物的活性氧代谢对干旱胁迫的响应 [D]. 杭州: 浙江农林大学, 2013.]
[24] TSIKAS D. Assessment of lipid peroxidation by measuring malondial- dehyde (MDA) and relatives in biological samples: Analytical and biological challenges [J]. Anal Biochem, 2017, 524: 13–30. doi: 10. 1016/j.ab.2016.10.021.
[25] JI Y, ZHANG X Q, PENG Y, et al. Effects of drought stress on lipid peroxidation, osmotic adjustment and activities of protective enzymes in the roots and leaves of orchardgrass [J]. Acta Pratacult Sin, 2014, 23(3): 144–151. [季杨, 张新全, 彭燕, 等. 干旱胁迫对鸭茅根、叶保护酶活性、渗透物质含量及膜质过氧化作用的影响 [J]. 草业学报, 2014, 23(3): 144–151. doi: 10.1168/cyxb20140316.]
[26] TONG S H, LIU N, WANG J, et al. Ecological and physiological adaptabilities ofto tropical coral island [J]. Guihaia, 2020, 40(3): 384–394. [童升洪, 刘楠, 王俊, 等. 长春花()对热带珊瑚岛生理生态适应性研究 [J]. 广西植物, 2020, 40(3): 384–394. doi: 10.11931/guihaia.gxzw201902010.]
[27] ZHOU W M, LIU N, CAI H Y, et al. Ecophysiological adaptability ofto tropical coral islands [J]. Guihaia, 2021, 41(6): 897–904. [周婉敏, 刘楠, 蔡洪月, 等. 莲叶桐对热带珊瑚岛环境的生理生态适应性 [J]. 广西植物, 2021, 41(6): 897–904. doi: 10.11931/guihaia.gxzw202007030.]
[28] LIU W Y, HE B, YANG F R, et al. Physiological response to drought and re-watering of different quinoa varieties [J]. Pratac Sci, 2019, 36(10): 2656–2666. [文瑜, 何斌, 杨发荣, 等. 不同品种藜麦幼苗对干旱胁迫和复水的生理响应 [J]. 草业科学, 2019, 36(10): 2656– 2666. doi: 10.11829/j.issn1001-0629.2019-0698.]
[29] Ingram J, Bartels D. The molecular basis of dehydration tolerance in plants [J]. Ann Rev Plant Biol, 1996, 47(1): 377–403. doi: 10.1146/annurev.arplant.47.1.377
[30] LI H X. Physiological response mechanism and resistance evaluation of six peony cultivars to drought stress [J]. N Hort, 2021(4): 64–71. [李海霞. 六个牡丹品种对干旱胁迫的生理响应机制及抗性评价 [J]. 北方园艺, 2021(4): 64–71. doi: 10.11937/bfyy.20201252.]
[31] LI J, CUI Y T, BAI Y W, et al. Physiological response and drought resistance evaluation of two kinds wolfberries on drought stress [J]. J Gansu Agric Univ, 2019, 54(5): 79–87. [李捷, 崔永涛, 柏延文, 等. 两种枸杞对干旱胁迫的生理响应及抗旱性评价 [J]. 甘肃农业大学学报, 2019, 54(5): 79–87. doi: 10.13432/j.cnki.jgsau.2019.05.010.]
[32] ZHAO C C, LI X N, ZHANG Y K, et al. Physiological correspondence of four varieties of perennial ryegrass to drought stress [J]. Pratac Sci, 2020, 37(4): 669–677. [赵春程, 李晓宁, 张寅坤, 等. 4个多年生黑麦草品种对干旱胁迫的生理响应 [J]. 草业科学, 2020, 37(4): 669– 677. doi: 10.11829/j. issn.1001-0629.2019-0528.]
[33] WANG L H, MIAO J, ZUO J C, et al. Physiological response ofvar.from different provenances under drought stress and evaluation of drought tolerance [J] J SW For Univ, 2019, 39(1): 69–79. [王连红, 苗杰, 左进城, 等. 不同种源单叶蔓荆对干旱胁迫的生理响应与耐旱性评价 [J]. 西南林业大学学报, 2019, 39(1): 69–79. doi: 10.11929/j.swfu.201804044.]
[34] Wang S, Zhou Q, Zhu Z L. Physiological and biochemical charac- teristics ofseedlings under drought stress [J]. Acta Bot Boreali-Occid Sin, 2013, 33(12): 2459–2466. [王飒,周琦,祝遵凌. 干旱胁迫对欧洲鹅耳枥幼苗生理生化特征的影响 [J]. 西北植物学报, 2013, 33(12): 2459–2466.]
[35]ASHRAF M, AKRAM N A, AL-QURAINY F, et al. Drought tolerance: roles of organic osmolytes, growth regulators, and mineral nutrients [J]. Adv Agron, 2011, 111: 249–296. doi: 10.1016/B978-0-12-387689-8. 00002-3.
[36] ZHANG L P, LIU Y, WANG Y, et al. The effects of NaCl stress on growth and accumulation of penetration adjustment substances in[J]. J Inner Mongolia Agic Univ (Nat Sci), 2020, 41(3): 10–15. [张林平, 刘艳, 王洋, 等. NaCl胁迫对甘草生长和渗透调节物质积累的影响 [J]. 内蒙古农业大学学报(自然科学版), 2020, 41 (3): 10–15. doi: 10.16853/j.cnki.1009-3575.2020.03.003.]
[37] LIU X, GAO R M, DANG S N, et al. Physiological response of leaves ofunder drought stress [J]. N Hort, 2020(14): 110–117. [刘翔, 高润梅, 党裳霓, 等. 干旱胁迫下穿龙薯蓣叶片的生理响应 [J]. 北方园艺, 2020(14): 110–117. doi: 10.11937/bfyy. 20193640.]
[38] KONG W P, ZHANG X A. Physiological response and drought resi- stance evaluation of four fig varieties to drought stress [J]. Acta Agric Boreali-Sin, 2019, 34(S1): 105–113. [孔维鹏, 张小艾. 四个无花果品种对干旱胁迫的生理响应及抗旱性评价 [J]. 华北农学报, 2019, 34 (S1): 105–113. doi: 10.7668/hbnxb.20190269.]
Physiological Response ofunder Drought Stress
CHEN Yilan1,2, LIU Dongming1,2*, WANG Jun1,2, JIAN Shuguang1,2, WANG Faguo1,2
(1. South China Botanical Garden, Chinese Academy of Sciences, Engineering Laboratory for Vegetation Ecosystem Restoration on Islands and Coastal Zones, Innovation Academy of South China Sea Ecology and Environmental Engineering,Guangzhou 510650, China; 2. South China National Botanical Garden,Guangzhou 510650, China)
Tropical coral islands have adverse natural conditions, and a very fragile vegetation ecosystem. In order to investigate the suitability ofin arid environment of tropical coral islands, the physiological indexes of stress resistance of leaves were studied.The results showed that there was no significant difference in malondialdehyde (MDA) content of leaves among drought degrees at early stress stage.The MDA content rapidly increased at first and then decreased quickly under severe drought stress, which had little change under other drought treatments. The activities of superoxide dismutase (SOD), peroxidase (POD) and catalase (CAT) increased with the increment of drought degree. Along the treatment time, SOD activity increased, POD activity was basically stable, and CAT activity decreased first and then increased. The soluble protein (SP) content under light drought stress was lower than that under control, which increased with increment of drought stress. The change in proline (Pro) content increased first and then decreased with the treatment time, but there was no significant difference among stress treatments at 18th day. Therefore,had strong drought resistance ability and could be used for artificial plant community construction, vegetation restoration and improving environmental conditions of Nansha Islands.
; Tropical coral island; Drought stress;Physiological response
10.11926/jtsb.4553
2021-10-27
2022-01-04
科技基础资源调查专项(2018FY100107);国家重点研发计划项目(2021YFC3100400);广东省科技计划项目(2019B121201005)资助
This work was supported by the Special Project for Science & Technology Basis Resources Investigation of China (Grant No. 2018FY100107), the Project for National Key Research & Development of China (Grant No. 2021YFC3100403), and the Project for Science and Technology Planning in Guangdong (Grant No. 2019B121201005).
陈意兰(1992年生),女,硕士,研究方向为植物资源评价与利用和特殊生境的生态修复。E-mail: chenyilan@scbg.ac.cn
E-mail: liudm@scbg.ac.cn
猜你喜欢
现代临床医学(2021年6期)2021-11-20
女报(2020年7期)2020-08-17
世界科学(2020年1期)2020-02-11
儿童故事画报(2019年8期)2019-08-14
中国生物医学工程学报(2019年5期)2019-07-16
中成药(2017年12期)2018-01-19
Coco薇(2017年5期)2017-06-05
学生天地(2016年6期)2016-04-16
现代电生理学杂志(2015年4期)2015-07-18
小火炬·智漫悦读(2015年4期)2015-06-05
