
干旱胁迫对疏叶骆驼刺幼苗生长和生理的影响
2023-03-16 01:55徐梦琦高艳菊张志浩黄彩变曾凡江
干旱区研究 2023年2期
徐梦琦, 高艳菊, 张志浩, 黄彩变, 曾凡江
(1.中国科学院新疆生态与地理研究所,新疆荒漠植物根系生态与植被修复重点实验室,新疆 乌鲁木齐 830011;2.中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室,新疆 乌鲁木齐 830011;3.新疆策勒荒漠草地生态系统国家野外科学观测研究站,新疆 策勒 848300;4.中国科学院大学,北京 100049)
干旱是限制植物生长发育和影响生态系统功能的重要环境胁迫之一,不仅影响植物各器官的生物量累积和分配,也能调节其形态、生理等性状,继而影响到种群结构和生态系统功能[1-3]。荒漠植物在长期进化过程中对干旱胁迫已表现出具有较强的可塑性和生态适应性。叶片和根系是快速感知干旱胁迫的敏感器官,其功能性状变化被认为是反映植物生存与环境变化间的纽带[4-5]。植物生长发育依赖于叶片光合的同化作用和根系吸收作用,因此叶片和根系也是揭示植物对干旱适应策略的关键器官[6]。因此,探讨水分胁迫对极端干旱区荒漠植物叶片和根系性状的影响和协同变化规律,对阐明荒漠植物应对水分胁迫的适应策略有重要意义[7-8]。
干旱胁迫会导致荒漠植物叶面积减小,叶片气孔关闭、光合速率降低来减少水分散失[9],同时增大叶厚度与根长,积累脯氨酸、可溶性糖、丙二醛等功能性状来增强其抗旱性[4,8-10]。而干旱会造成植物体内活性氧的累积,影响其生理代谢功能[11]。干旱条件下植物地上与地下的生理生态过程不是独立存在的,而是密切关联的。在内蒙古草原对植物性状间关系的研究发现,地上与地下器官能够协同变化以适应外界环境改变[12]。但是叶片和根系的功能性状以及二者之间的耦合关系也与不同程度的环境胁迫程度及生长时期有关[13]。目前,极端干旱区植物水分适应策略的研究主要集中在叶片[14-16],而对荒漠植物根系性状及其与叶片的协同变化方面的关注较少。
骆驼刺是多年生豆科草本植物,也是典型的荒漠深根植物,广泛分布于塔克拉玛干沙漠南缘(简称“塔南”)[17],在该区域的风沙治理和生态修复中发挥着重要作用[18]。众多学者已围绕多年生骆驼刺的形态结构、生理特征、根系分布、抗旱抗盐碱能力等方面开展了系统研究[19-20],但是对于骆驼刺幼苗在干旱胁迫下地上和地下器官功能性状及其关联性研究还相对较少。本研究通过人工控制的盆栽试验来模拟干旱胁迫,设置充分灌溉、轻度胁迫、重度胁迫3 个水分处理,在不同生长时期测定骆驼刺幼苗叶片和根系的干物质含量、形态特征、渗透调节物质和过氧化产物含量,分析不同水分条件下叶片和根系功能性状的变化与生理响应,以期探讨骆驼刺幼苗的水分适应策略,旨在为策勒绿洲-沙漠过渡带的植被恢复与重建提供科学依据。
1 材料与方法
1.1 研究区概况
本试验在中国科学院策勒荒漠草地生态系统国家野外科学观测研究站(80°03′E~82°10′E,35°17′N~39°30′N)进行。研究区地处塔克拉玛干沙漠南缘策勒绿洲-沙漠过渡带,属典型的大陆性干旱气候,平均海拔1360 m,多年平均降雨量仅35.1 mm,平均蒸发量高达2600 mm[17]。该地区风沙较大,年均8级以上大风天数达40 d,沙尘暴天气年均30 d[21]。土壤以风沙土为主,持水力差。绿洲内植物主要依赖于夏季洪水和地下水得以生存。由于风沙活动频繁,夏季高温,该区域环境条件恶劣,植被稀疏,覆盖度仅有15%。该区域的优势植物以多年生草本植物骆驼刺(Alhagi sparsifolia)为主,并伴生有花花柴(Karelinia caspica),还有少量的多枝柽柳(Tamarix ramosissima)和胡杨(Populus euphratica)等。
1.2 试验设计
2021 年4 月18 日进行盆栽种植。试验选用聚乙烯螺纹管(直径30 cm,高度60 cm)作为盆栽容器,底部封闭。所用土壤取自策勒绿洲外围荒漠区0~30 cm 土层的混合土壤。土壤pH 为8.47,有机质含量为3.31 g·kg-1,全氮、全磷和全钾含量分别为0.22 g·kg-1、0.62 g·kg-1和16.27 g·kg-1。每桶装土50 kg,充分浇水后播种。播前种子用温水浸泡12 h,每盆播种6 粒种子,播种深度为3 cm。在幼苗生长稳定前进行统一管理,保持土壤湿润。待幼苗长至3~4 片叶时按照长势一致的原则进行剔苗,每桶留苗1株。
前期研究发现,骆驼刺幼苗存活的土壤含水量阈值为6%[22-23],研究区域土壤最大田间持水量为18%。据此共设定3 个水分梯度:(1)充分灌溉(CK):土壤含水量为最大田间持水量的70%~75%;(2)轻度干旱胁迫(W1):土壤含水量为最大田间持水量的50%~55%;(3)重度干旱胁迫(W2):土壤含水量为最大田间持水量的30%~35%。于2021 年6 月18日开始按照3个水分梯度进行水分处理,每个处理6个重复。整个生长季内每间隔3 d 采用TDR300 土壤水分测定仪(soil moisture equipment,Santa Barbara,CA,USA)测量各处理的土壤含水量,计算水分损耗并进行补灌,补灌后使各处理盆栽土壤含水量分别达到各自设定值范围,补水时间固定为北京时间19:00。
1.3 样品采集与测定
1.3.1 样品采集 分别于2021年7月18日(处理30 d,生长前期)和9月18日(处理90 d,生长后期)进行样品采集,每次各处理采集3 个重复。每个重复随机采集10片完整成熟叶片带回实验室称重后,用于叶片形态指标测定;另采集5 g 新鲜成熟叶片于-20 ℃冷冻保存,用于测定叶片生理指标,将地上部分剩余样品的叶、茎和刺分离,称重后于75 ℃烘干称重。根系用0.15 mm 筛网冲洗取出,用扫描仪进行根系扫描后,截取5 g新鲜根系于-20 ℃冷冻保存用于测定根系生理性状,剩余根系烘干称重。
1.3.2 形态指标及生物量测定 采集的10 片叶用精度为0.01 g 的天平称重获得叶鲜重。叶片叠加,用精度为0.01 mm 的电子游标卡尺测量叶前端、中端、尾端3个点的叶厚度(LT),取平均值,测量时避开主脉位置。用扫描仪对叶片进行扫描,利用Image J软件分析图像获得叶面积(LA),将扫描后的叶片放入75 ℃烘箱烘至恒重获得叶干重。根据公式得到:比叶面积(SLA)=叶面积/叶干重、叶组织密度(LTD)=叶干重/(叶面积×叶厚度)、叶干物质含量(LDMC)=叶干重/叶鲜重。
根系采用扫描仪进行扫描后,用WinRHIZO Pro根系分析系统对图像进行分析获得根长(RL)、根表面积(RSA)、根体积(RV)。分离5 g新鲜根系后,剩余根系烘干至恒重,用电子天平(精度为0.01 g)称得干重。根据公式得到:比根长(SRL)=根长/根干重、根组织密度(RTD)=根干重/根体积、根冠比(R/S)=地下生物量/地上生物量。
1.3.3 生理指标测定 骆驼刺叶片和根系的脯氨酸、可溶性糖、丙二醛的测定参照李合生[24]的方法:脯氨酸(Pro)含量测定采用酸性茚三酮比色法,可溶性糖(SS)含量采用蒽酮比色法测定,丙二醛(MDA)含量采用硫代巴比妥酸法测定。
1.4 数据处理
数据统计分析用SPSS 16.0 进行。用单因素方差分析(one-way ANOVA)分别对不同水分处理下骆驼刺叶片和根系的生物量、形态性状和生理参数进行分析(95%置信区间),多重比较采用Duncan检验法;利用Perason 相关系数分析各性状间的相关性;利用Origin 2018 和GraphPad Prism 8.0.1 绘图。数值均为平均值±标准差。
2 结果与分析
2.1 干旱胁迫对骆驼刺幼苗生物量的影响
干旱胁迫对骆驼刺幼苗的生物量累积有显著影响(P<0.05),各部分生物量在2个时期均呈现出:CK>W1>W2的变化趋势,且CK处理显著高于其他处理(表1)。在生长前期,骆驼刺的地上与地下生物量在W1下较CK 减少了48.90%、27.46%;在W2下较CK 减少了74.08%、45.91%;地上生物量>地下生物量。而生长后期,尽管各部分生物量也随水分减少显著降低,但地下生物量>地上生物量。根冠比在前期和后期均随水分减少显著增加(P<0.05)。表明干旱胁迫虽抑制骆驼刺幼苗整体的生长,但其地下生长能力在干旱胁迫下有所增强。

表1 干旱胁迫下骆驼刺的生物量累积Tab.1 Biomass allocation of A.sparsifolia under drought stress
2.2 干旱胁迫对骆驼刺幼苗叶和根形态性状的影响
干旱胁迫对骆驼刺幼苗的叶面积、叶组织密度、叶片干物质含量的影响因生长时期而异(P<0.05,图1)。在生长前期,与CK处理相比,叶面积在W1和W2处理下显著减小(P<0.05),而比叶面积、叶组织密度和叶干物质含量则受干旱胁迫影响不明显。在生长后期,叶面积在W2处理下显著下降(P<0.05),叶组织密度和叶干物质含量随干旱胁迫程度的增强而增加,在W2处理下显著高于其他处理,比叶面积在各处理间无明显差异。
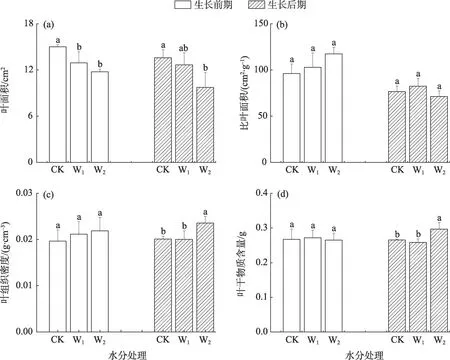
图1 干旱胁迫下骆驼刺叶片形态性状的变化特征Fig.1 Changes of leaves morphological traits of A.sparsifolia under drought stress
不同水分处理下骆驼刺幼苗根系形态特征也发生了显著变化(P<0.05,图2),且在不同时段呈现出不同的变化规律。在生长前期,根长和表面积在W1和W2处理下显著下降(P<0.05);比根长的变化趋势为:W2>CK>W1,仅W2与W1处理间存在显著差异(P<0.05);根组织密度的变化趋势与比根长相反,表现为:W1>CK>W2,W1处理显著高于CK 和W2处理(P<0.05)。在生长后期,骆驼刺幼苗根长在W1处理下值最小,且显著低于CK处理(P<0.05)。干旱胁迫使骆驼刺根表面积显著减小(P<0.05),比根长在W1处理下无明显变化,W2处理下显著增加(P<0.05),根组织密度则不受水分变化的影响。

图2 干旱胁迫下骆驼刺幼苗根系形态性状的变化特征Fig.2 Changes of roots morphological traits of A.sparsifolia seedlings under drought stress
2.3 干旱胁迫下骆驼刺幼苗叶和根的渗透调节物质及丙二醛(MDA)的变化
干旱胁迫显著增加了骆驼刺幼苗叶片的脯氨酸和丙二醛含量(P<0.05,图3)。在生长前期,W2处理下骆驼刺幼苗叶片的脯氨酸含量显著大于CK与W1处理(P<0.05),比CK 与W1分别高出341.72%与228.57%;丙二醛也在W2处理下显著高于CK 处理(P<0.05),较CK增加了30.77%;不同处理下可溶性糖含量无显著变化。在生长后期,骆驼刺幼苗叶片的可溶性糖在W1和W2处理下显著降低(P<0.05),与CK 处理相比分别减少了30.63%和23.20%;脯氨酸在W2条件下显著增加(P<0.05),比CK和W1处理分别高出135.29%和100.67%。分别对比各指标在不同时期间的变化发现,W1处理下骆驼刺叶片的丙二醛含量在生长后期有所降低;W2处理下脯氨酸和丙二醛含量均在生长后期有所降低。

图3 干旱胁迫下骆驼刺叶片生理性状的变化Fig.3 Changes of leaf physiological traits of A.sparsifolia under drought stress
干旱胁迫后骆驼刺幼苗根系渗透调节物质和丙二醛含量在不同生长时期变化也不同(图4)。在生长前期,根系可溶性糖和脯氨酸含量的变化规律与骆驼刺叶片相同,不同处理间的可溶性糖含量无显著差异;脯氨酸随胁迫程度加剧而显著增加,但CK 与W1处理差异不明显。丙二醛在W1处理下值最小,且显著低于CK处理。在生长后期,不同处理间骆驼刺幼苗根系脯氨酸含量无显著差异;丙二醛含量随干旱胁迫增强而逐渐增大,在W2处理下显著高于CK 与W1处理。可溶性糖含量的变化趋势为CK>W2>W1,各处理间差异显著(P<0.05)。
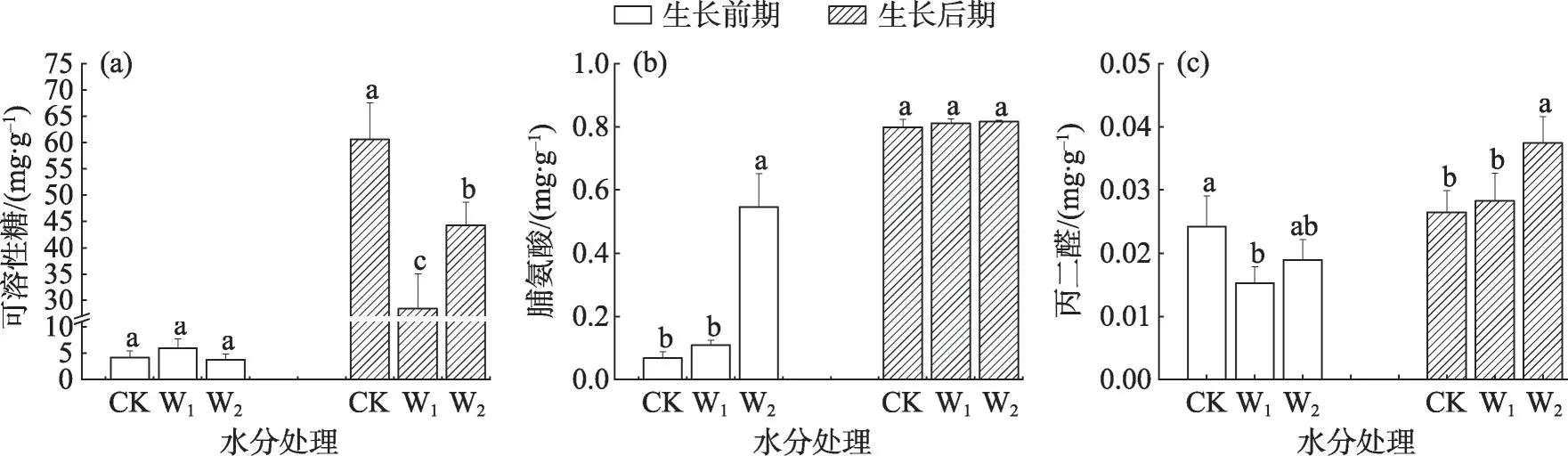
图4 干旱胁迫下骆驼刺根系生理性状的变化Fig.4 Changes of root physiological traits of A.sparsifolia under drought stress
2.4 骆驼刺幼苗叶和根性状间的关联关系
骆驼刺幼苗叶片和根系形态性状间Pearson 相关性分析表明,根系性状间及叶与根系性状间存在较强的耦合关系,而叶片性状间耦合关系相对较弱(图5)。叶-叶性状间耦合关系主要表现为:叶组织密度与叶脯氨酸含量呈显著正相关(P<0.05),与叶干物质含量呈极显著正相关(P<0.05);比叶面积与叶MDA含量呈极显著正相关(P<0.01),与干物质含量呈显著负相关(P<0.05),与叶可溶性糖含量呈极显著负相关(P<0.01);叶面积与叶脯氨酸呈极显著负相关(P<0.01)。根形态与生理性状间耦合关系较强,渗透调节物质与根长、比根长、根表面积间均呈现出显著相关性。而根形态间耦合关系主要集中在根长、比根长和根表面积之间,根组织密度与各性状不相关。根生理性状与叶形态、叶MDA含量间存在较强的权衡关系,而根生理与叶可溶性糖含量间存在显著协同关系。叶片与根系在渗透调节物质、过氧化产物MDA、比叶面积-比根长4对性状间存在性状平行关系,其余性状在叶和根之间无平行性。
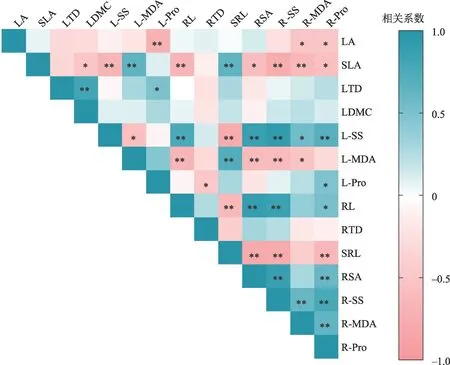
图5 骆驼刺性状间相关关系Fig.5 The correlation between the characters of A.sparsifolia
3 讨论
3.1 干旱胁迫对骆驼刺幼苗生长的影响
生物量的累积能够直接反映植物营养物质的分配关系和生理生态过程,是衡量植物生产力的重要指标之一[25],同时也是植物适应胁迫环境(如干旱)的重要策略之一[26]。生物量的分配表征植物生长发育中将同化的资源分配到各器官的过程,是植物生存权衡的结果[27]。大量研究发现,较低的土壤含水量会显著抑制植物干物质的形成,但可通过增加地下生物量的分配比例来增强自身的抗旱性[28-29]。与前人研究结果一致,在本研究中,骆驼刺幼苗各部位生物量均随干旱程度的增加而显著减少。这可能是因为干旱通过削弱地上的光合作用,从而导致整体干物质累积受限[30-31]。骆驼刺幼苗的地下生物量在生长后期时显著大于地上生物量,根冠比也在各时期均随水分胁迫增强而显著增加。因此,根冠比的增大提升了其对水分和养分获取的能力,同时地上生物量的减少也有效防止过多的水分散失,这种生物量分配策略与大多植物的抗旱策略一致[29,32]。
叶片和根系是植物资源获取的主要器官,具有较强的可塑性,其功能性状直接反映了植物光合固碳、资源获取的能力,与植物的适应策略密切相关[14,33]。干旱胁迫下,植物往往会通过控制叶片气孔的闭合,保持体内水分的平衡[34],并将更多的光合产物用于干物质的积累,维持细胞的正常运转[7]。本研究表明,骆驼刺幼苗在干旱胁迫下主要通过减小叶面积降低蒸腾失水,并在生长后期提高了叶组织密度和叶干物质含量,这可能加强了叶片的韧性和紧实程度,自身的防御力提高。该结果与其他植物适应干旱胁迫的策略一致[3,7,34]。在本研究中,尽管随干旱胁迫增强比叶面积变化并不显著,但生长后期时,比叶面积在一定程度上有所降低,这表明骆驼刺在生长后期地上资源获取能力有所减弱。
根系作为与土壤直接接触的资源器官,对土壤水分与养分的变化十分敏感。荒漠植物通常拥有庞大根系,便于在水分亏缺的条件下向下生长获取水分和养分[2,23]。本研究发现,骆驼刺的根长、表面积、组织密度在干旱条件下均显著减小,而比根长显著增加。这表明较低的土壤水分显著限制骆驼刺根系生物量的累积,但可通过比根长和根组织密度提高加强了骆驼刺根系的资源获取能力,减缓干旱胁迫[35],骆驼刺在前期研究中也有类似发现[21]。
3.2 骆驼刺幼苗叶和根对干旱胁迫的渗透调节能力
植物能够通过改变自身渗透调节物质的变化、自由基的产生、细胞质膜功能的变化等多种生理过程形成对干旱的适应性[8,36]。渗透调节是植物细胞通过渗透调节物质的累积降低渗透势,维持细胞膨压,保持吸水能力的一种适应干旱的方式[30,37]。脯氨酸作为游离态的渗透调节物质能够有效防止植物细胞失水,平衡渗透压,可溶性糖与脯氨酸作用相似[38]。土壤水分较低时,植物体内脯氨酸和可溶性糖会大量积累以增强抗旱性[39-40]。本研究中,骆驼刺幼苗叶片和根系的脯氨酸在生长前期时均在重度干旱胁迫时最大,生长后期叶片脯氨酸依旧在重度干旱胁迫下显著高于对照和轻度胁迫,而根系脯氨酸含量在后期趋于平稳,这表明骆驼刺具有较强的抗氧化和防脱水能力[41],能通过累积脯氨酸提高根系吸水能力,以此增强渗透调节力,在叶片水势降低的同时维持细胞膨压。此外,骆驼刺幼苗叶片和根系的可溶性糖在前期并无变化,在后期均明显增加,但干旱胁迫后均低于对照。张美云等[42]研究发现,在一定程度的胁迫条件下,脯氨酸的累积会早于可溶性糖的累积,二者间存在相互补偿关系,这与笔者的研究结果一致。生长后期造成骆驼刺体内可溶性糖变化的原因可能是干旱环境使骆驼刺叶片减小了蒸腾耗水[43],且生长后期骆驼刺根冠比增大,主要以脯氨酸作为渗透调节物质增强抗逆性。
MDA是植物细胞过氧化产物,能够直接反映出细胞损伤程度和植物抗旱性[44],随着胁迫时间的延长,MDA含量也会在一定程度上增加[11]。本研究发现,生长前期骆驼刺幼苗叶片MDA随胁迫程度增加而逐渐提高,根系MDA 则呈下降趋势;后期叶片MDA 无显著变化,根系则显著增加。该结果反映出,在生长前期,干旱条件导致了骆驼刺叶片细胞出现损伤,而根系细胞过氧化水平相对稳定。随着土壤水分胁迫的持续,骆驼刺地上部分逐渐产生适应性;而胁迫引起的过氧化水平升高使根系细胞受到一定的损伤,这说明骆驼刺幼苗在面对不利环境时,在不同生长阶段的适应能力不同。
3.3 水分胁迫下骆驼刺幼苗的适应策略
植物地上与地下的生长、生理代谢过程密切相关,功能性状不是彼此独立,而是相互关联发挥作用[4]。植物能结合功能性状共同调节适应环境变化,并以此生成最佳的性状组合[11]。刘金环等[45]和Wilson 等[46]认为,比叶面积与叶干物质含量是植物对资源利用和适应的最佳组合,叶干物质含量的增加和比叶面积的减小使叶片水分散失减弱。而Wright 等[10]发现,植物能调控比叶面积和叶组织密度的权衡以适应环境变化。在本研究中,骆驼刺存在比叶面积与叶干物质含量、叶组织密度间的权衡,表明骆驼刺能有效维持叶片水分散失、干物质积累和防御力间的平衡。同时,骆驼刺叶片和根系分别在其形态与渗透物质间、MDA 间存在耦合关系,这说明叶片和根系感应到胁迫后能调控其生长和生理代谢间的平衡以维持抗旱能力。骆驼刺的叶片和根系在渗透调节物质、过氧化产物以及比叶面积-比根长间存在平行关系,表明骆驼刺体内的生理代谢和资源获取过程具有较明显的整体性。与此同时,随着骆驼刺地下渗透调节能力和抗旱性的增强,地上叶片获取光资源和进行光合作用的能力相对减弱,反映了骆驼刺在叶形态与根生理相关的核心性状间存在权衡,这与赵广帅等[47]对羌塘高原降水梯度下植物地上与地下性状关系的研究结果一致。这也突出了地上与地下间的生长是密不可分的。根系作为首要感应土壤水分变化的器官,当骆驼刺根系感受到干旱胁迫时主动调控根系细胞内渗透物质的产生,并影响根系生长和资源获取情况,从其形态上适应胁迫[48-49];地上部也通过调控叶片减少水分散失和消耗,而影响叶片渗透物质含量的变化,骆驼刺体内碳同化产物在各器官间分配比例也发生变化,同时干旱引起骆驼刺体内MDA的累积破坏生物膜,也能够间接影响骆驼刺各器官生长发育。
干旱胁迫下骆驼刺叶片与根系性状的变化不一致,这也表明地上与地下所面对压力和环境不同,而骆驼刺叶和根性状间具有更强的权衡关系。综合各性状的变化趋势,虽然受干旱的影响骆驼刺整体生长发育受限,但其投入在根系的资源高于叶片,通过调控根系生长增强水分和养分的获取以及防御力,并储存干物质和养分维持地上生长,地上部分也通过适当减弱水分损失,为地下生长提供有利条件,这种适应策略与骆驼刺在极端干旱环境下的自我保护和适应性密切相关。
4 结论
本文研究了干旱胁迫对骆驼刺叶和根系形态、生理以及生物量累积的影响,得出以下结论:
(1)骆驼刺通过减小叶面积,提高叶组织密度和干物质含量来增强叶片紧实度,减弱叶片水分散失。
(2)干旱条件刺激骆驼刺叶片和根系的MDA累积,生物膜受损。
(3)干旱胁迫下骆驼刺的比根长以及根脯氨酸含量增加,提高了根系资源获取和渗透调节能力。
(4)生长后期骆驼刺幼苗根冠比显著增大,重度干旱条件下尤为显著。长期干旱胁迫使骆驼刺将更多干物质集中用于投资根系的资源获取和抗逆性防御构建,在干旱胁迫下整体表现出缓慢投资、保守生长的适应策略。
猜你喜欢
中老年保健(2022年3期)2022-11-21
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
阅读与作文(小学高年级版)(2020年2期)2020-05-21
新疆农业科学(2020年1期)2020-02-14
科学与财富(2019年15期)2019-10-21
发酵科技通讯(2018年2期)2018-07-06
领导文萃(2018年3期)2018-03-12
北方人(2018年3期)2018-03-08
湖南林业科技(2017年1期)2017-02-06
红蜻蜓·低年级(2016年4期)2016-11-19
