
高原土壤菌株的分离及抗菌活性初步评价
2023-03-30 03:13郭欢欢林文星胡运琪马晓莉南泽东江志波吴秀丽
农业与技术 2023年6期
郭欢欢林文星胡运琪马晓莉南泽东江志波吴秀丽
(1.北方民族大学化学与化学工程学院餐厨废油转化清洁能源与高附加值化学品创新团队,宁夏 银川 750021;2.宁夏医科大学药学院,宁夏 银川 750004)
病原微生物(真菌、细菌和病毒等)引起的严重感染是造成临床死亡的重要因素之一。2019年世界卫生组织(WHO)公布的全球十大健康威胁就有埃博拉、登革热和艾滋病毒等病原体感染[1]。2019-新型冠状病毒(Novel coronavirus pneumonia,NCP,COVID-19)更因其严重的人传人性质,迅速引发肺炎、严重呼吸道综合征、肾衰竭甚至死亡等特征,成为传播速度最快,发病到死亡时间最短的恶性传染性病原体。2020年1月17日WHO的一份报告指出,在研的药物中针对革兰氏阴性菌的很少。因此加快抗革兰氏阴性菌药物的研发刻不容缓[2-4]。
微生物在长期进化过程中,通过与环境和各种生命体相互作用及斗争中获得大量的次级代谢产物生物合成基因簇(Biosynthetic gene clusters,BGCs),并通过编码低分子化合物、多肽和蛋白质等形式保障生存、繁殖和进化。微生物次级代谢产物及其衍生物因其形式多样,结构复杂且具有良好的生物活性,一直是临床药物的重要来源,在保障人类生命健康方面作出了重要的贡献。20世纪40—70年代为抗生素发展的黄金时期,一大批抗生素的临床使用,有效遏制了感染性疾病的发生和传播。但随着抗生素的广泛使用和局部地区的滥用,越来越多的病原菌对已有抗生素产生了耐药性。而更不利的是,自20世纪80年代以来,其研发进度严重减缓,目前也只有一些中小企业在坚持。2017年WHO报告全球12株致命超级细菌亟需开发新型抗生素,也给全世界药企和相关科研单位敲响了警钟。
抗生素发展速度减慢因素有很多,其中最主要的就是人们对陆地和淡水生境的实验室可培养的微生物次级代谢产物进行大量研究,已经很难从中获得具有新结构的活性化合物来作为新抗生素开发的先导化合物。而基因组挖掘技术(通过分子生物学激活潜在的隐性生物合成基因簇)获得新结构化合物方法,不仅受到实验方法的限制,还需要付出高昂的代价。特殊生境微生物(指生存在火山口、深海、高原盐湖和盐碱地等生态环境下的菌群)通常对热、辐射、高压或高盐具有耐受性,次级代谢产物中通常含结构新型、活性多样的化合物,成为当前研发新抗生素的重要方向。
本实验对青海湖土壤样品和一份大型真菌进行微生物分离,共得到49株菌株。借助于平板法检测部分菌株的抗菌活性,发现1株放线菌和5株真菌发酵液对普通变形菌(Proteus vulgaris)具有明显的生长抑制作用。本实验为进一步探究其次级代谢产物的多样性奠定基础,为理解青藏高原生物多样性和以此为基础的微生物药物研发奠定基础。
1 材料与方法
1.1 仪器试剂
超净工作台(YT-CJ-1ND),北京亚泰科隆仪器有限公司;高压蒸汽灭菌锅(SQ810C),雅马拓科技贸易(上海)有限公司;摇床(ZHWY-211C),上海智城分析仪器制造公司;真空冷冻干燥机(FD-1A-50),北京博医康实验仪器有限公司;甲醇、氯仿、丙醇、石油醚,酵母浸膏、马铃薯重铬酸钾培养基,国药集团化学试剂有限公司;乙醚、青霉素,天津市北联精细化学品开发有限公司;正己烷、链霉素,天津市光复精细化工研究所;葡萄糖、放线菌培养基,多点化学;磷酸氢二钾(AR),天津科密欧化学试剂有限公司。
1.2 培养基
1.2.1 高氏l号培养基
1.2.2 马铃薯培养基
马铃薯200g,加水1L,熬制泥状,纱布过滤,定容至1L;上述溶液中依次加入葡萄糖20g,TRYPTONE 3g,琼脂15g,MgSO41g,K2HPO40.5g,酵母浸膏1g;115℃灭菌15min。使用前,加入链霉素(120μg·mL-1)和青霉素(120g·mL-1),用于真菌筛选。
1.2.3 马丁培养基
葡萄糖1g,蛋白胨0.5g,KH2PO4·3H2O 0.1g,MgSO4·7H2O 0.05g,孟加拉红溶液(0.1g·mL-1)0.33mL,琼脂1.5~2g,蒸馏水100mL,pH6.4;115℃灭菌15min。使用前,加入2mL的2%去氧胆酸钠溶液及0.33mL的链霉素溶液(10000μmol·mL-1),迅速摇匀,作为筛选真菌的生长及保存培养基。
1.2.4 MH肉汤培养基
如果通信电缆沟中布设的人孔井数量较少,在实际作业期间,可以在原观测点处,完成相应的埋设,也可以通过现浇的方式,制作独立水泥观测墩,通过对其进行应用,完成相应的观测工作[6]。
牛肉粉2.0g,可溶性淀粉1.5g,酸水解酪蛋白17.5g,琼脂12~20g,蒸馏水1L,pH7.4;121℃灭菌20min。用于检定菌株生长和抗菌活性评价。
1.2.5 Luria-Bertani(LB)培养基
酵母提取物5g,胰化蛋白胨10g,NaCl 10g,蒸馏水1L,NaOH调pH7.0,于121℃高压灭菌20min。用于稀释土壤样品和植物样品。
1.3 样品来源
样品于2019年07月28—31日在青海省青海湖边和柴达木盆地采集。新鲜样品用无菌器具采集,无菌袋密封,-20℃保藏。
1.4 实验方法
菌株分离和样品前处理、活性测试等方法见郭欢欢等[5]文章。
1.4.1 样品采集与处理
采用5点采集法,取地面下40~60cm深度的土壤。将土壤样品上下均匀混合后装入无菌信封,标注采样信息,所有样品低温保存。
1.4.1.1 土壤样品处理
超净台中,取4份土壤样品各10g,加入90mL无菌LB液体培养基,混合均匀。依次稀释至0.01g·mL-1、0.001g·mL-1。
1.4.1.2 菌株生长
取2种浓度悬浊液200μL涂布在固体平板上。静置0.5h后,倒置于恒温培养箱中,28℃进行培养。马铃薯培养基15~20d,高氏一号培养基5~7d,期间观察菌落生长情况。
1.4.2 菌株分离
用无菌竹签挑取单菌落,固体培养基“Z”划线培养。放线菌使用高氏一号培养基;真菌使用马丁培养基,重复3次,直至菌落在显微镜下观察无异性菌。确定为单一菌落的菌株利用无菌竹签接种至斜面培养基,正常生长至生长期,4℃冰箱保存;对于产孢放线菌使用20%甘油洗涤孢子,-70~-80℃低温冰箱中保存;产孢真菌使用50%甘油洗涤孢子,保存于-70~-80℃低温冰箱中。
1.4.3 菌株鉴定
1.4.3.1 光学显微镜镜检
超净台中,将灭菌的盖玻片45°斜插入各个培养基中,每个培养基插入3~5片,放入恒温培养箱中培养1~2d,根据菌株生长情况取盖玻片进行显微镜观察,保存菌丝体清晰的图片,并记录。
1.4.3.2 电子显微镜成像
将导电胶于超净台中紫外灯照射灭菌,用无菌牙签挑取单菌落,将单菌落均匀涂抹在导电胶上。蔡司扫描电子显微镜(EVO MA 10/LS 10型)观察视野中完整菌株,分别获取×50、×200、×2000倍图像。以图片编辑工具给出菌株的直径(或长、宽),记录菌株的形态。
1.4.4 活性评价方法
1.4.4.1 样品制备
固体发酵琼脂块捣碎,乙酸乙酯超声萃取3次,每次20mL。合并有机溶剂,减压蒸馏回收有机溶剂。残留物以2mL甲醇溶解,作为待测液。
液体摇瓶发酵液10mL,上样至固相萃取色谱柱中,以10mL蒸馏水洗涤后,依次使用2mL的50%甲醇水溶液、95%甲醇水溶液洗涤,减压干燥,样品存放于玻璃瓶中,于4℃下保存。
1.4.4.2 活性测试方法
直径6mm的加厚滤纸上滴加200μL样品,自然风干(2h),并平铺于检定菌(金黄色葡萄球菌)的平板上。金黄色葡萄球菌检定板于28℃恒温培养箱中培养12~24h。观察样品的抑菌情况。
2 结果
2.1 青海湖边土壤样品中菌株分离结果
利用特异性菌株筛选培养基从青海湖边土壤样品中获得23株菌株,包括17株真菌,见图1(QHH-1~17),6株放线菌,见图1(QHH-18~23)。真菌在马铃薯培养基生长状态显示菌落较大,形态各异。除QHH-6、QHH-7、QHH-11、QHH-16外,其它真菌皆有明显的同心环。所有真菌均有绒状气生菌丝。
放线菌的菌落直径约为真菌的1/10。QHH-18和QHH-19 2株放线菌无气生菌丝,菌落光滑,有颜色较浅的色素分泌,其中QHH-18分泌色素为淡黄色。QHH-20菌体显示为红色,可能为放线紫红素。QHH-18与QHH-23外观非常相似,也同样具有淡黄色的色素代谢物,比较明显的差别在于QHH-23具有白色气生菌丝。
2.2 柴达木盆地土壤样品中菌株分离结果
柴达木盆地土壤样品比较干燥,从中分离得到了24株菌株,包括16株真菌和8株放线,见图1。从菌落形态来看,DLHX-1、DLHX-3、DLHX-4形态相似,都存在同心环,差别在于菌丝体颜色和代谢产物色素的颜色差别较大。DLHX-1中心圈显色为白色,而DLHX-3和DLHX-4分别为黑色和紫色。DLHX-10和DLHX-13可能都是霉菌,差别在于DLHX-10菌丝为青色,DLHX-13为绿色。放线菌中DLHX-18、DLHX-19、DLHX-20无气生菌丝,菌体表面光滑,有微微凸起。DLHX-20菌株可分泌亮黄色色素与其它菌株差别较明显。DLHX-22菌株中心为绿色菌丝体,周边有白色绒毛状气生菌丝,在所有放线菌中少见。DLHX-21~24具有气生菌丝,可能为放线菌属的链霉菌。具体菌株分离还将通过其它理化性质和16S或18S rRNA测序鉴别。
2.3 青藏高原大型真菌样品中菌株分离结果
大型真菌采集于西宁至青海湖途中日月乡。从大型真菌外壁培养得到1株真菌(MGWC-1),而将其组织破碎后,通过培养得到1株放线菌(MGWC-2)。真菌MGWC-1在培养时间10d之前为气生菌丝,白色;10d以后中心部位出现绿色菌丝体。放线菌MGWC-2培养到第5天时出现气生菌丝白色;第3天开始产生黄色素,且其扩散性强,提示可能为水溶性物质。
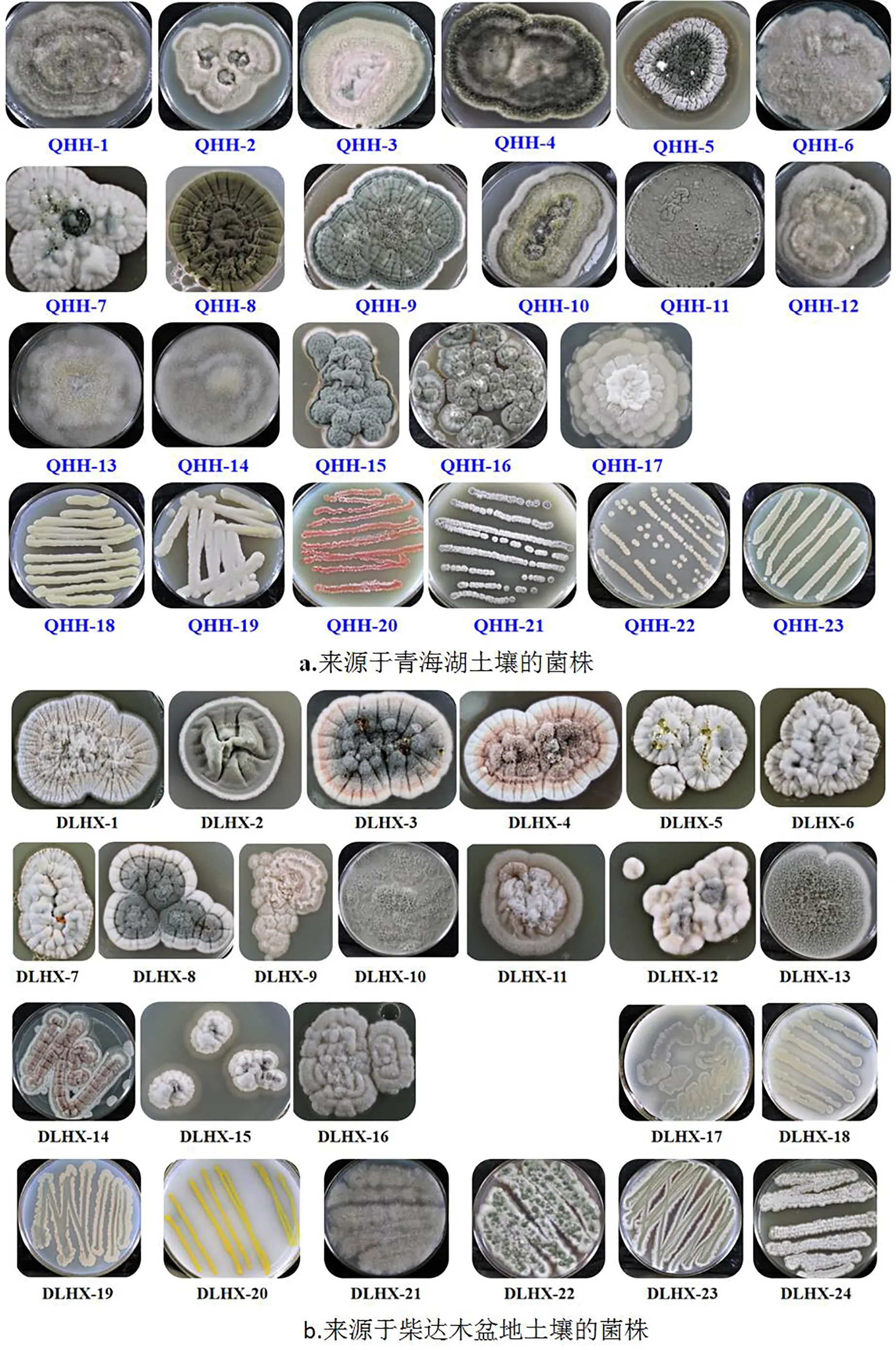
注:QHH-1~17为真菌;QHH-18~23为放线菌;DLHX-1~16为真菌;DLHX-17~24为放线菌。
2.4 抗菌活性测定结果
普通变形杆菌属于变形杆菌属(Proteus),广泛存在于水、土壤、腐败有机物以及人和动物的肠道中,最常见的是普通变形杆菌(P.vulgaris),此外还有奇异变形杆菌(P.mirabilis)、摩根氏变形杆菌(P.morganii)及瑞特杰尔变形杆菌(P.rettgeri),性状皆大同小异。普通变形杆菌有明显的多形性,可呈球形和丝状等多形态,无芽孢、无荚膜,有周鞭毛,运动活泼。本实验以普通变形杆菌(P.vulgaris)为检定菌,利用平板扩散法(液体发酵样品)和纸片扩散法(含有机溶剂样品),对23株菌株样品进行了抑菌活性测试。测定结果显示,柴达木盆地土壤来源真菌DLHX-5和放线菌DLHX-1~5具有对P.vulgaris生长抑制作用。
3 讨论
土壤菌株多样性是全球生物多样性的重要组成部分,不仅是通过病原或共生与植物形成非常密切的联系,而且还是凋落物和植物残体降解的“主力军”。研究表明,从局部领域尺度到全球领域尺度,植物多样性与土壤菌株多样性的耦合关系并非一致,特别是在高寒生态系统的植物生产力、植物多样性和菌株多样性的关系还不是很明确。
青藏高原作为全球海拔居首位的高原,被称为“第三极”。该地区气候低、养分贫乏以及生态系统也是十分的脆弱,全球气候变化极为敏感。尽管青藏高原自然环境差,但是该区域依旧栖息着大量的微生物,促使整个生态系统的元素生物有序的循环。对整个高原生态系统的组成和维持具有不可忽视的作用,同时具有重要的研究意义。
中国科学院南京土壤研究所褚海燕教授课题组研究了青藏高原高寒草原土壤菌株的多样性以及菌落组成。着重讨论了地上植物与土壤菌株在空间分布上的耦合关系。研究结果表明,地上植物的β多样性和α多样性与土壤菌株的β多样性和α多样性呈现出强烈的耦合关系。偏最小二乘回归模型、多元线性回归以及方差分解,分析共同揭示了植物多样性对土壤菌株多样性的驱动作用,但植物生产对菌株多样性的作用并不明显,该研究指出,制约土壤菌株多样性的首要因素是地上植物多样性。此外,溶解性有机碳、土壤碳氮比和土壤总磷对土壤菌株多样性也有显著影响,结果对我国高寒草原土壤菌株多样性的资源开发利用以及保护具有一定的意义。
4 结论
本论文通过系统菌株分离纯化,从青海湖和柴达木盆地2份土壤和1株大型青藏高原菌株样品中分离得到49株菌,有真菌34株、放线菌15株。所有菌株进行斜面或甘油保存,为进一步探究其次级代谢产物多样性奠定基础。对青藏高原微生物多样性研究国内起步比较晚,文献报道,与其它生境生长的微生物相比,青藏高原土壤微生物具有耐寒和耐紫外线活性,本论文由于时间关系无法考究所有菌株的这些性质。
猜你喜欢
陕西农业科学(2022年2期)2022-03-28
复旦学报(医学版)(2021年4期)2021-08-05
浙江农业学报(2021年5期)2021-05-31
国际呼吸杂志(2019年22期)2019-12-09
上海农业学报(2017年4期)2017-04-10
广东饲料(2016年5期)2016-12-01
现代食品(2016年24期)2016-04-28
食品工程(2015年3期)2015-12-07
电测与仪表(2015年20期)2015-04-09
应用海洋学学报(2014年4期)2014-11-22
