
植物SUS基因家族的系统进化及其在玉米中的干旱诱导表达分析
2023-05-27 01:06丁鹏钧陈君如王睿蕾张恺文刘梦真韩兆雪
干旱地区农业研究 2023年3期
吴 越,丁鹏钧,陈君如,王睿蕾,张恺文,刘梦真,韩兆雪
(西北农林科技大学生命科学学院,陕西 杨凌 712100)
蔗糖合酶(Sucrose synthase,SUS)是植物中以基因家族形式广泛存在的一种糖基转移酶。在植物体内SUS可以催化光合作用产物蔗糖的水解,促进蔗糖和尿苷二磷酸(UDP)转化为果糖和尿苷二磷酸葡萄糖(uridine diphosphate glucose,UDP-G)[1],从而进一步参与淀粉和纤维素的合成代谢等过程,因此SUS基因在植物碳源分配及糖信号调控途径中扮演重要角色[2]。根据已有报道,SUS基因参与了植物生长发育和非生物胁迫响应过程[3]。拟南芥内的6个SUS基因存在转录表达模式差异,敲除突变体分析发现它们之间存在功能冗余[4];水稻在成熟期受到高温胁迫时SUS3基因表达增加,提高水稻高温抗性[5];水稻中OsSUS3的大量表达可以增强其纤维素和胼胝质的积累,进而增加生物乙醇产量并提高抗逆性[6];当存在氧胁迫时,小麦和杨树中SUS活性提高进而增加细胞壁厚度、促进纤维素积累以及提高抗逆性[7];黄瓜CsSUS3基因反义表达植株的低氧胁迫抗性显著降低,表明CsSUS3参与黄瓜低氧胁迫响应[8];在高镉浓度下,玉米枝条中的蔗糖合酶活性提高,根系中蔗糖合酶活性降低[9];另外,蔗糖合酶还具有潜在的抗旱胁迫功能,例如在干旱胁迫下番茄SUS5在叶片和根中的表达显著下调[10],黄瓜CsSUS3基因在幼根和幼叶中表达上调,蔗糖合酶活性增加[11],大豆GmSusy1和GmSusy2在叶片和种子中表达量增加。
目前,SUS基因家族的系统发育研究已经在拟南芥、水稻、番茄、葡萄、苹果和芥菜等多个单物种中广泛展开[12-15],但对多物种中SUS基因家族的系统进化研究仅有1篇文献报道,该文献主要对19个物种的100个SUS基因进行了系统进化分析,初步分析了该基因家族的进化特征[16]。已有研究表明,多物种的系统发育进化分析更有利于探究基因家族的起源,有助于全面呈现物种演化过程中基因家族成员的保留、丢失和功能分化等过程。因此,为全面准确地揭示植物SUS基因家族的系统进化规律及功能特征,本研究拟对包括绿藻、苔藓、蕨类、裸子植物和被子植物在内的50个代表物种中的SUS基因家族进行大规模系统的生物信息学分析,推断植物SUS家族的系统发育轨迹,进一步结合RNA-seq基因表达数据,以玉米SUS家族为例,分析干旱胁迫诱导SUS基因的表达模式,这将有助于深入理解高等植物SUS基因表达和功能分化与物种进化的关系,进一步为SUS基因响应非生物胁迫的功能研究提供参考。
1 材料和方法
1.1 50个物种中SUS基因家族的鉴定
从Pfam数据库中下载SUS家族的两个保守蛋白质结构域Sucrose synth(PF00862)和Glycostransf 1 (PF00534)的隐马尔可夫模型(HMM,Hidden Markov Model),使用软件HMMER中的hmmsearch对包含34种双子叶植物、11种单子叶植物、1种基部被子植物、1种裸子植物、1种蕨类植物、1种苔藓植物和1种绿藻植物在内的50个具有代表性的物种的候选SUS蛋白进行鉴定[17-18],阈值设置为E=0.001;进一步利用拟南芥内已鉴定的6个SUS蛋白进行Blastp检验[19]。最后使用Conserved Domain Database (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)在线数据库将所得候选蛋白序列进行分析,去除结构域不完整的序列[20]。50个物种基因组序列和注释文件从GitHub(https://github.com/zhaotao1987)内获取[21-22]。
1.2 SUS基因家族的序列比对、系统进化及基因共线性分析
使用MAFFT对鉴定出的SUS蛋白质序列进行多序列比对[23]。以来自胶球藻(Coccomyxasubellipsoidea)的SUS蛋白作外类群[24],使用FastTree构建系统发育树。使用DIAMOND进行物种间和种内的基因组序列比对,MCScanX对共线性区域进行检测[25],Rstudio对结果进行调取,Gephi将共线性结果可视化。
1.3 SUS蛋白的理化性质和亚细胞定位预测
使用ExPASy Compute pI/Mw、GRAVY CALCULATOR和TMHMM - 2.0等在线数据库分析SUS候选蛋白序列的分子量、等电点、亲疏水性及跨膜结构域等理化性质。使用YLoc对SUS蛋白进行亚细胞定位预测[26-27]。
1.4 干旱胁迫下玉米SUS基因的表达分析
在GEO数据库[28]获取玉米正常生长条件与干旱条件下SUS基因的转录组表达数据(GSE71723)[29],探究干旱胁迫下玉米SUS基因的响应表达情况。使用FPKM(Fragments per kilobase of transcript per million fragments mapped)表示SUS基因的表达水平,当基因的|log2(fold change)|≥1且错误发现率(FDR,False discovery rate)≤0.05时被认定为差异表达基因。
2 结果与分析
2.1 植物SUS基因家族的鉴定及系统发育分析
50个植物物种包括大豆、苹果、拟南芥和棉花在内的34种双子叶植物、包括水稻、玉米和小麦在内的11种单子叶植物、1种基部被子植物、1种裸子植物、1种蕨类、1种苔藓和1种绿藻。在50种植物内共鉴定出283个SUS基因,对基因数目分析发现,三个早期物种卷柏(Selaginellamoellendorffii)、小立碗藓(Physcomitrellapaten)和胶球藻(Coccomyxasubellipsoidea)中共含有6个SUS基因,裸子植物云杉(Piceaabies)含有2个,其余275个SUS基因都存在于被子植物中,其中基部被子植物无油樟(Amborellatrichopoda)中鉴定出2个SUS基因,大部分双子叶植物和单子叶植物的SUS基因数目为5~8个,双子叶植物中基因数目大于10个的有四个物种,如桉树(Eucalyptusgrandis)和大豆(Glycinemax)内分别有11个和 12个,蔷薇科植物苹果(Malusxdomestica)和梨(Pyrusxbretschneideri)内分别有15个和14个;单子叶植物内SUS基因数目均不超过10个,例如玉米(Zeamays)内有6个,高粱(Sorghumbicolor)内有5个。结合物种进化树上显示的基因组复制事件,我们发现4个SUS基因数目大于10的双子叶植物物种均发生过全基因组二倍化或三倍化复制事件,推测其基因组经历全基因组多倍化扩增出的SUS基因被大量保留下来,而尽管单子叶植物中也发生了多次全基因组二倍化事件,但在物种进化过程中很多扩增基因发生了丢失;另外,双子叶植物猕猴桃(Actinidiachinensis)和甜菜(Betavulgaris)内仅有2个SUS基因,蓖麻(Ricinuscommunis)中只有1个SUS基因,这可能也与全基因组多倍化事件后的基因丢失有关[30](图1)。
以胶球藻的SUS基因(38455)作外类群,对SUS基因的推断氨基酸序列进行系统发育分析,根据系统发育树的拓扑结构(图2,见76页),发现3个早期物种包括胶球藻 (1个)、小立碗藓(4个)和卷柏(1个)的6个SUS基因聚为一个原始进化枝,其余SUS基因聚类在3个进化枝内,分别命名为SUS Ⅰ、SUS Ⅱ和SUS Ⅲ 亚家族。SUS Ⅰ是最大的亚家族,包含来自基部被子植物(1个)、单子叶植物(30个)和双子叶植物(79个)的110个SUS基因;SUS Ⅱ是最小的亚家族,其包含来自单子叶植物(8个)与双子叶植物(57个)的65个SUS基因;SUS Ⅲ由来自裸子植物(3个)、基部被子植物(1个)、单子叶植物(21个)和双子叶植物(78个)的102个基因组成。3个亚家族中的单子叶和双子叶植物来源的成员分别聚类于不同的分枝内,其中双子叶植物成员的进化枝均按照真双子叶植物-茄科-蔷薇分支种类依次分布。这表明在早期物种中,SUS基因并未分化,3个亚家族是在物种长期进化过程中产生的。
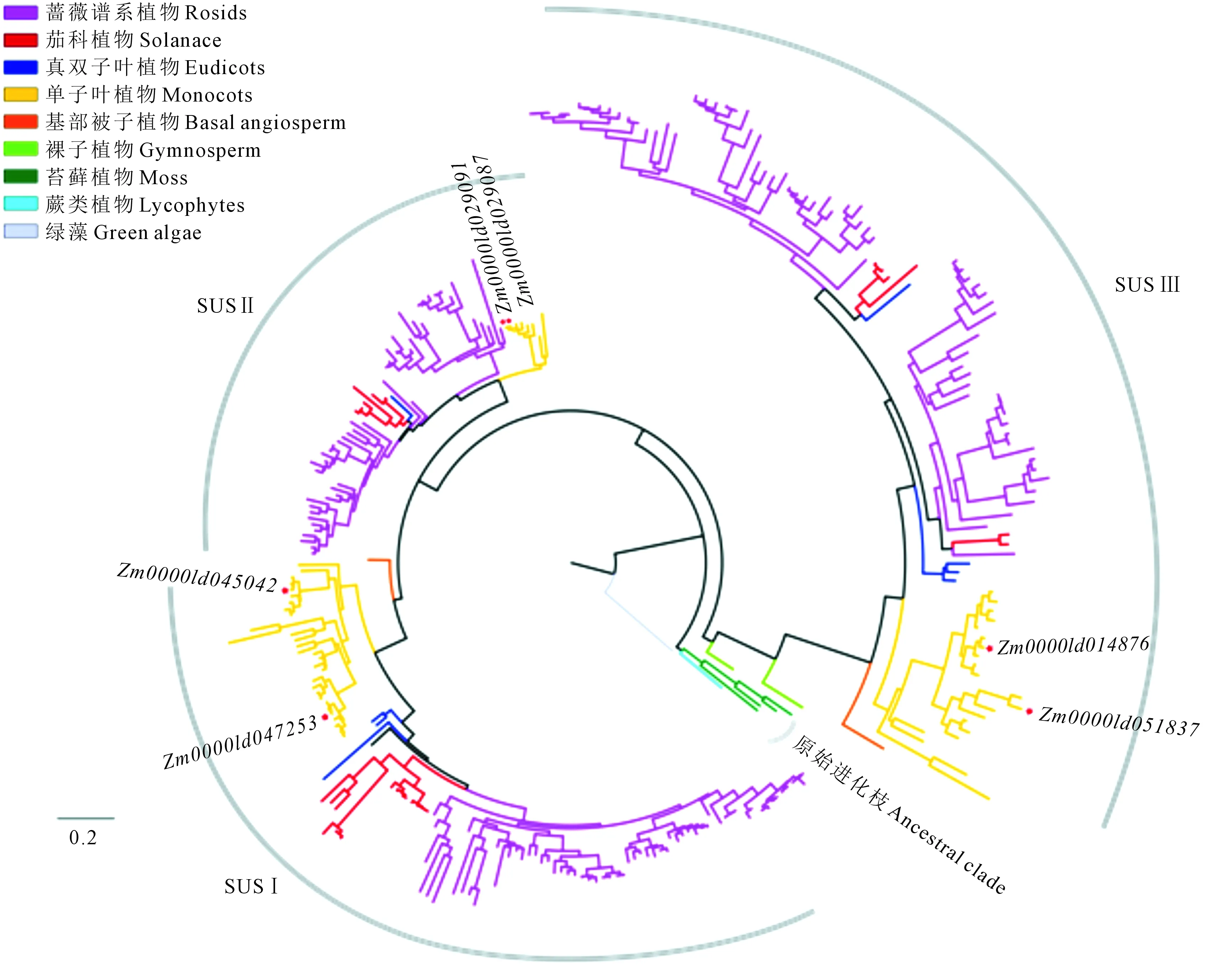
注:图中五角星代表玉米SUS基因。Note:The maize SUS genes are marked byasterisks.图2 SUS基因家族的系统发育树Fig.2 Phylogenetic tree of SUS gene family
2.2 SUS基因家族的理化特征分析
对SUS基因家族推断蛋白的理化性质分析发现,不同亚家族SUS蛋白相对分子质量大部分分布在90 000~100 000 kD,SUS Ⅰ和SUS Ⅱ亚家族内的成员平均分子量较为接近,约为93000 kD,SUS Ⅲ成员平均分子量约为101 000 kD(图3a);SUS Ⅰ和SUS Ⅱ蛋白的等电点较接近,均为pI<7,为酸性蛋白,而SUS Ⅲ 中则有39.2%(40/102)的蛋白等电点pI>7,为碱性蛋白(图3b)。多肽序列疏水性分析表明所有SUS蛋白均为亲水性蛋白(GRAVY<1);此外,我们共鉴定出23个含有跨膜结构域的SUS蛋白,其中SUS Ⅲ含有20个,SUS Ⅰ、和SUS Ⅱ分别含有2个和1个(图3c)。这一结果表明,SUS Ⅰ和SUS Ⅱ亚家族内的成员序列较保守,进化变异较少,而SUS Ⅲ亚家族成员蕴含着较高的序列变异性,该亚家族成员理化性质变异较大。

图3 SUS蛋白理化性质分析Fig.3 Physicochemical properties of SUS protein
亚细胞定位预测结果显示,SUS蛋白家族定位于细胞质、细胞核、线粒体、过氧物酶体和质膜等,其中细胞质内含有245个SUS蛋白,包括早期进化物种如卷柏(1个)和小立碗藓(4个)的蛋白;细胞核内预测出31个SUS蛋白,除了1个来源于无油樟,其余30个均来源于双子叶植物;质膜内预测出3个SUS蛋白,1个来源于双子叶植物甘蓝,另外2个分别来源于单子叶植物玉米和二穗短柄草;过氧物酶体内预测出2个SUS蛋白,均来源于猕猴桃;线粒体内预测出1个来源于二穗短柄草的SUS蛋白(图3d)。这一结果表明不同物种谱系内的SUS蛋白的亚细胞定位已产生分化,可能各自执行不同的功能。
2.3 SUS基因家族的共线性分析
共线性分析在多物种基因家族分析中是了解种间和种内基因及基因组进化轨迹的有力方法[31]。通过对283个SUS基因进行共线性分析发现,其中有197(197/283,69.86%)个基因为共线性同源基因,这些共线性同源基因间存在1969对共线性关系,仅存在于被子植物中。结合系统发育进化关系,我们发现SUS Ⅰ、SUS Ⅱ和SUS Ⅲ亚家族内分别存在962对、587对和411对共线性基因关系,大部分共线性基因对的两个基因均位于同一亚家族内,而有9对共线性关系是由不同亚家族之间的基因产生的(图4)。进一步对这9对基因分析发现,有8对共线性关系都是由SUS Ⅲ的菜豆(Phaseolusvulgaris)基因(Phvul.006G087300)与SUS Ⅱ的 8个基因之间产生的,该基因还与SUS Ⅲ的11个基因有亚家族内部的共线性关系(图4b);另一对则来源于SUSⅠ的紫色假雀麦(Brachypodium_distachyon)基因(Bradi1g46670)和SUS Ⅱ的香蕉(Musaacuminata)基因(GSMUA_Achr8G22470),Bradi1g46670在SUS Ⅱ内部仅有1个共线性基因,而GSMUA_Achr8G22470在SUSⅠ内部有21对共线性基因(图4b)。进一步对共线性结构进行可视化分析发现,SUS Ⅲ的共线性网络又可细分为两个亚群,两个亚群内部各自有213对和166对共线性基因,而亚群之间仅有23对共线性基因,我们推测在SUS家族分化为三个亚家族后,SUS Ⅲ在后面的进化过程中进一步分化为SUS Ⅲ-A和SUS Ⅲ-B两个亚群,表明两个亚群的成员可能发生了潜在的功能分化;结合系统发育树分析发现,SUS Ⅲ亚家族内的单子叶植物所在的进化枝对应于B亚群,双子叶植物进化枝对应于A、B两个亚群(图4b),这表明双子叶植物来源的SUS Ⅲ亚家族基因可能比同家族的单子叶植物来源的基因具有功能多样性。
综合系统发育分析和共线性分析可推测出植物SUS基因家族的进化轨迹(图5,见77页)[32],结果表明,SUS基因家族在蕨类植物到种子植物的进化过程中由一次复制事件分化为两个亚家族,分别是SUS Ⅲ和SUSⅠ&Ⅱ的共同祖先,SUSⅠ&Ⅱ的共同祖先在挪威云杉(Piceaabies)中可能已丢失;在被子植物分化以前,SUSⅠ&Ⅱ的共同祖先由一次复制事件扩增为SUSⅠ和SUS Ⅱ亚家族,SUS Ⅱ在基部被子植物无油樟中可能已丢失;并且SUS Ⅲ在双子叶植物中可能具有继续分化为SUS Ⅲ-A和SUS Ⅲ-B的趋势。
2.4 玉米SUS基因家族的干旱胁迫响应表达模式分析
为探究SUS基因在植物应对非生物胁迫中的生物学功能,以玉米为例,分析了正常生长和干旱胁迫下玉米三个组织(叶、雄穗和雌穗)在四个发育时期(V12、V14、V18和R1)内SUS基因家族成员的表达情况。
玉米SUS基因家族包含SUSⅠ-Ⅲ 3个亚家族,共有6个成员,每个亚家族各有2个成员。同正常生长条件(WW)相比,干旱胁迫(DS)处理下玉米中4个SUS基因表达水平呈现差异,亚家族之间的基因表达具有明显不同的表达模式,同一亚家族内的两个基因呈现出相似的表达模式,尤其SUSⅡ内的两个基因在不同组织的不同发育时期均具有极为相似的表达趋势和表达水平。SUS Ⅰ有一个差异表达基因(Zm00001d045042),干旱胁迫下,Zm00001d045042在叶片的R1时期表达上调(|log2(Fold change)|=1.82143,FDR=1.88×10-4);SUSⅡ中两个基因均呈现差异表达,其中Zm00001d029087在雌穗的R1时期受干旱诱导表达上调(|log2(Fold change)|=1.47467,1.88×10-4),在Tassel_V12表达上调(1.80696,9.37×10-4),Zm00001d029091在Ear_R1表达上调(1.7783,4.55×10-3),在Ear_V12表达上调(1.93171,3.65×10-4);SUSⅢ有一个基因差异表达,Zm00001d014876在Ear_R1表达下调(-1.85212,3.56×10-4),在Leaf_R1表达上调(1.03997,1.88×10-4)(图6,见78页),玉米SUS基因在干旱胁迫下的表达水平变化表明其响应干旱胁迫调节,有潜在的抗旱功能。另外,SUSⅢ的两个基因的表达模式不同,Zm00001d014876在R1和V18两个时期有表达高峰,而Zm00001d051837在各个时期都只有低水平的表达,这暗示了SUSⅢ中可能发生的功能分化或冗余。
3 讨 论
随着植物全基因组序列的公开,越来越多SUS基因家族从不同单物种中得到鉴定,然而基于多物种进行系统分析SUS基因家族的研究还鲜见报道。本研究整合物种进化、氨基酸序列、基因共线性关系以及玉米SUS家族的基因结构和干旱诱导转录表达谱分析,对50个物种内鉴定出的283个SUS基因进行了综合分析。

注:绿色节点代表SUSⅠ基因,橙色节点代表SUSⅡ基因,黄色节点代表SUSⅢ-A基因,深黄色节点代表SUSⅢ-B基因,棕色节点代表SUSⅢ的其它基因,蓝色节点代表原始进化枝的基因。Note:Each node represents a gene of the SUS family.Green nodes represent genes of SUSⅠ,orange nodes represent genes of SUSⅡ,yellow nodes represent genes of SUSⅢ-A subgroup,dark yellow nodes represent genes of SUSⅢ-B subgroup,brown nodes represent the other SUSⅢ genes,and blue nodes represent genes of the ancestor clade.图4 SUS基因家族的共线性关系Fig.4 Synteny analysis of SUS gene family
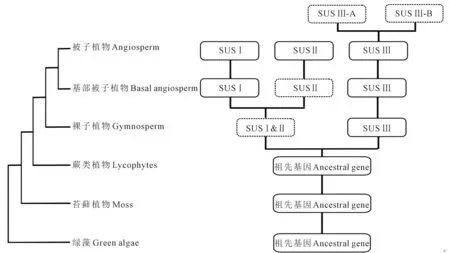
注:虚线框代表推测存在的亚家族共同祖先和可能出现的进一步分化结果。Note:The dashed box represents the potential common ancestor of the subfamily and possible further differentiation results.图5 植物SUS基因家族进化轨迹Fig.5 The evolutionary track of the plant SUS gene family
系统发育分析结果表明,植物SUS基因家族在三个早期物种胶球藻、小立碗藓和卷柏中未出现明显的亚家族结构分化,从裸子植物挪威云衫开始出现结构的分化,SUS祖先基因最先进化出SUS Ⅲ亚家族,到基部被子植物无油樟中出现SUS Ⅰ亚家族的分化,而单子叶和双子叶植物中又进化出SUS Ⅱ亚家族,这可能表明SUS基因家族的结构分化是从裸子植物和被子植物的共同祖先中开始出现的,每一次新的结构分化可能伴随着物种的分化而产生。此外,三个亚家族中的单子叶和双子叶植物来源的成员分别呈现明显的分簇聚类,这与文献[16]报道中10种双子叶植物和5种单子叶植物SUS基因家族进化的分簇聚类模式一致;有趣的是,我们发现SUSⅠ中所有双子叶植物的基因仅聚类在一个进化枝内,而SUSⅡ和SUS Ⅲ内双子叶植物的基因聚类在两个分枝内,这可能暗示了SUS Ⅰ中双子叶植物的基因功能更趋于保守,而SUS Ⅱ和SUS Ⅲ中的双子叶植物的基因产生了潜在的功能分化;然而,三个亚家族内的双子叶植物的基因进化枝存在明显不同,其中SUS Ⅱ内的双子叶植物的两个基因进化分枝的物种类别不同,其中一个分枝内没有鉴定出茄科植物(马铃薯、番茄、辣椒、双峰狸藻、猕猴桃)和真双子叶植物(莲和甜菜)中的SUS基因,这表明这些基因可能在物种进化过程中发生了丢失,而其它双子叶植物基因进化分枝的物种均由真双子叶植物、茄科和蔷薇谱系的物种构成。
SUS基因家族的共线性分析表明,有197个(197/283,69.86%)基因为共线性同源基因,这些具有共线性的基因仅存在于40种被子植物中。基因间的共线性关系主要集中在三个亚家族内部,跨不同亚家族的基因共线性关系仅有9对,我们推测三个亚家族可能具有各自独立的进化模式,这与系统发育关系表明的SUS基因家族呈现出的三个亚家族的进化模式一致;其中SUS Ⅰ内含有最多的共线性基因对(962/1969,48.86%),SUS Ⅱ和SUS Ⅲ含有数目接近的共线性基因对。SUSⅠ的双子叶植物的52个基因之间形成747个共线性基因对,不同双子叶植物的基因之间具有广泛的共线性关系,而且单子叶植物和双子叶植物之间的共线性基因对也较多(163对),暗示SUSⅠ内基因的进化十分保守;然而,SUS Ⅱ内双子叶植物的49个基因之间形成了521个共线性基因对,SUS Ⅲ内双子叶植物的63个基因之间仅形成了362个共线性基因对。有趣的是,我们发现SUS Ⅲ成员的共线性网络中明显呈现出两个独立的共线性亚群SUS Ⅲ-A和SUS Ⅲ-B,而系统发育树(图2)分析表明,SUS Ⅲ亚家族成员分别聚类为单子叶植物和双子叶植物的两个进化枝,其中双子叶植物的基因进化枝又包括两个亚组,而其中的一个双子叶植物的进化枝亚组与单子叶植物的进化枝成员形成SUS Ⅲ-B共线性子网络,另一个双子叶植物的进化枝亚组单独形成SUS Ⅲ-A共线性子网络(图4),这表明两个双子叶植物的进化枝亚组可能在物种进化过程中出现了更多的序列变异,暗示SUS Ⅲ的两个双子叶植物的亚组成员在物种进化过程中具有继续分化的趋势,而且,与SUS Ⅰ和SUS Ⅱ亚家族相比,SUS Ⅲ内各成员也表现出更大差异的理化性质(图3)。
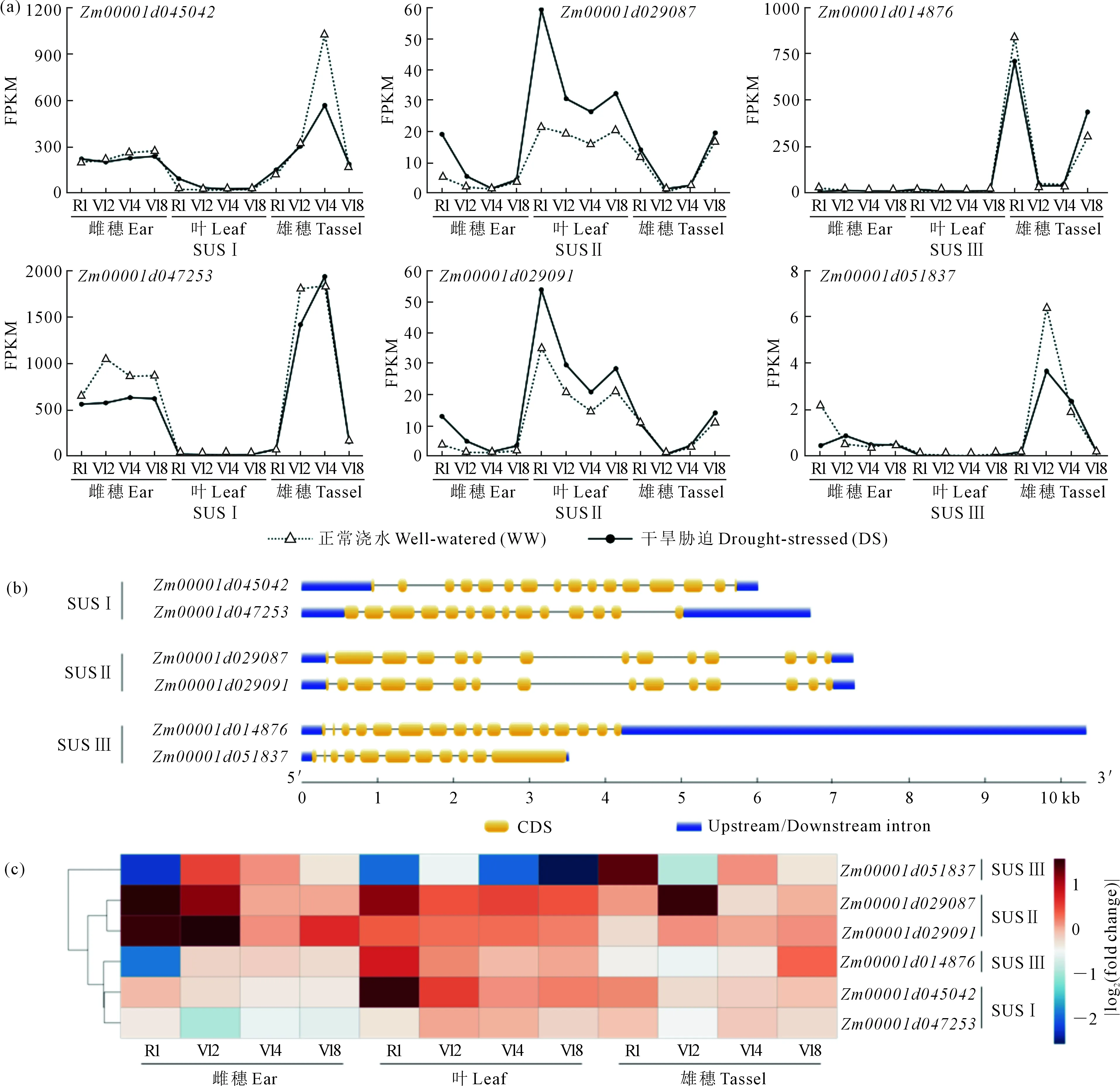
注:(a)正常和干旱条件下玉米SUS基因在各组织不同发育时期的表达水平;(b)玉米SUS基因内含子-外显子结构;(c)干旱条件下玉米SUS基因差异表达热图。Note:(a)Expressions profiles of maize SUS gene in three tissues of maize at four developmental stages under well-watered and drought-stressed conditions;(b)The intron-exon structure of SUS genes in maize;(c) Heatmap of expression levels of maize SUS genes under drought stress.图6 玉米SUS基因结构和表达模式Fig.6 Maize SUS gene structure and expression pattern
玉米干旱胁迫转录组数据表达分析进一步揭示了玉米SUS基因在干旱胁迫下的表达模式。共有4个玉米SUS基因受干旱诱导差异表达,这表明SUS基因在玉米干旱应答中起作用,这与关于禾本科作物抗旱胁迫研究类似。干旱胁迫处理后,大麦HvSs3转录表达水平升高[33],高粱蔗糖合酶基因SbSusy1、SbSusy3、SbSusy4和SbSusy5能提高高粱幼苗的抗旱性,SbSusy2在干旱胁迫处理后表达下调[34];而且SUS Ⅰ和SUS Ⅱ亚家族内的2个成员在玉米不同生长发育时期分别表现出相似的表达趋势或模式,而SUSⅢ内的两个基因的表达模式差异较大;基因结构分析表明,SUS Ⅰ和SUS Ⅱ亚家族内的2个成员分别具有相似的外显子-内含子结构特征,而SUS Ⅲ内的两个成员结构表现出较大差异,这充分说明了基因结构决定了基因表达的模式,进一步决定基因的功能。该结果也说明SUS Ⅰ和SUS Ⅱ亚家族成员的结构和表达具有各自的保守性,这两个亚家族可能来自于古老的被子植物祖先经过全基因组复制而产生,进一步产生了某些程度的功能分化;而SUS Ⅲ成员表现出更多的趋异特征,保守性较SUSⅠ、Ⅱ相比更弱,这也与三个亚家族各自的进化特征相一致。
现有的蔗糖合酶基因研究表明,蔗糖合酶基因广泛参与植物生长发育和胁迫响应,在抗低温、低氧、重金属、干旱响应中都发挥作用。本研究初步研究了植物蔗糖合酶家族的进化,并揭示其在玉米中发挥的干旱功能,为进一步研究玉米抗旱功能提供了依据。
猜你喜欢
科学与财富(2021年3期)2021-03-08
小哥白尼(军事科学)(2019年9期)2019-12-21
——以多重共线性内容为例
长沙航空职业技术学院学报(2019年2期)2019-07-13
温州大学学报(自然科学版)(2019年2期)2019-06-04
科学大众(中学)(2019年3期)2019-05-17
电影(2019年3期)2019-04-04
汽车观察(2018年10期)2018-11-06
阅读(低年级)(2018年11期)2018-05-14
少儿科学周刊·少年版(2017年3期)2017-06-29
科技知识动漫(2017年1期)2017-02-06
