
大豆品种齐黄34 广适性的遗传分析
2023-06-19 12:04刘薇王玉斌李伟张礼凤王彩洁徐冉戴海英张彦威
山东农业科学 2023年5期
刘薇,王玉斌,李伟,张礼凤,王彩洁,徐冉,戴海英,张彦威
(山东省农业科学院作物研究所/山东省特色作物工程实验室,山东济南 250100)
大豆对光周期敏感[1-3],这一特性直接反映在开花期、成熟期等生态适应性相关性状上。 具体表现为大豆品种在不同地区间引种会导致开花期和生育期改变,进而影响产量,严重时导致植株不能正常开花结实。 因此,大豆品种的适应范围非常狭窄,极大限制了品种的推广及产量突破。
开花期是大豆适应性的一个重要指标,对开花期调控基因的研究是大豆适应性改良的重要切入点。 目前,包括E 系列基因(E1~E11)、J基因以及Tof11/12等在内的与开花期相关的关键位点相继被发现[4]。 在这些基因中,E1、E2、E3和E4基因的相互组合与大豆的开花期、生育期性状密切相关[5-7],其中E1对开花期及生育期的影响最大[8-11]。E1基因为豆科作物所特有,在长日下特异表达,其过量表达极显著抑制大豆开花,是重要的开花抑制基因[12]。
齐黄34 是近年培育出的高产广适大豆品种[13],先后通过国家黄淮海北片和中片、山东省、江苏省淮北和淮南区、贵州省审定,通过安徽省、河南省引种审批,是农业农村部主导品种和主推的四大核心品种之一[14]。 该品种适应范围广,适种范围跨越了20 个纬度。 但是目前关于该品种的广适性遗传机理尚缺乏研究。
前期,我们利用齐黄34 和另一个黄淮海推广品种冀豆17[15]为杂交亲本衍生得到重组自交系群体,并基于此群体材料构建了高密度遗传图谱[16]。 本研究利用该群体针对开花期进行QTL 定位,以期阐明使齐黄34 适应范围广的QTL 位点和关键基因,同时为齐黄34 的遗传改良提供分子依据。
1 材料与方法
1.1 试验材料
试验材料为齐黄34、冀豆17 及其杂交衍生的由256 个家系成员组成的重组自交系(RIL)群体。
1.2 试验方法
1.2.1 表型统计 重组自交系群体分别在2018和2019 年6 月种植于山东省农业科学院济阳试验基地,行长为3 m,株距和行距分别为10 cm 和50 cm。 试验设3 次重复。 每行近一半的大豆植株开花时记为初花期。
齐黄34 和冀豆17 于2019 年种植于人工光照培养箱(温度为26℃,12 h 光照/12 h 黑暗)进行短日培养,分别统计两个品种植株的开花时间。
1.2.2 QTL 分析 使用软件IBM SPSS 对表型数据进行统计和分析。 使用QTL IciMaping V4.2 软件[17],利用区间作图法(IM-ADD)进行QTL 定位。 似然函数比值对数值(logarithm of odds,LOD)的阈值设为2.5。
1.2.3E1基因片段的克隆及序列分析 分别取齐黄34 和冀豆17 的叶片,提取RNA,并利用TAKARA 反转录试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser(RR047A,TAKARA)反转录。 以 E1 - F ( 5'-ATGAGCAACCCTTCAGATGAAAG-3') 和 E1 - R ( 5'-CTCCCTTATTGTTCATCTCCTC-3')为引物,利用KOD-FX 高保真酶(KFX-101,东洋纺生物科技有限公司)进行E1部分序列的克隆。 扩增体系:2×buffer 20 μL,2 mmol/L dNTP 8 μL,E1-F 1.5 μL,E1-R 1.5 μL,模板2 μL,KOD-FX 0.8 μL,ddH2O 4.2 μL。 扩增程序:94℃2 min;98℃30 s,53℃30 s,68℃20 s,30 个循环;68℃5 min。 扩增产物直接交由擎科生物技术有限公司进行测序。
1.2.4 开花促进基因GmFT2a实时荧光定量PCR检测 将齐黄34 和冀豆17 种植于人工气候室,从出苗第4 天开始,每隔4 天取最上部叶片,迅速置于液氮中,带回并保存在-80℃冰箱中备用。提取所有样品的RNA 并反转录成cDNA,利用实时荧光定量PCR 试剂盒TB GreenPremix Ex Taq(RR820A,TAKARA) 进行实时荧光定量PCR(qRT-PCR)检测。GmFT2a的检测引物为qGm-FT2a-F(5'-TGGGGGAGTAATTGGGGATG-3') 和qGmFT2a - R (5'-ACCTCATGGCCGAAACTAGC-3');内参基因的检测引物为qGmActin-F(5'-CGGTGGTTCTATCTTGGCATC-3') 和 qGmActin - R( 5'-GTCTTTCGCTTCAATAACCCTA-3')。 qRT -PCR 的反应体系:2×TB GreenPremix Ex Taq10 μL,上、下游引物各0.4 μL,模板2 μL,ddH2O 7.2 μL。 反应程序:95℃30 s;95℃10 s,60℃30 s,40个循环。 利用2-ΔΔCt方法[18]进行相对表达量的计算。
2 结果与分析
2.1 开花表型分析
表1 结果显示,RIL 群体开花期存在较大差异,存在超高亲和超低亲现象。 2018 年RIL 群体的开花期范围是28.0 ~57.0 d,标准误差为7.21,峰度和偏度分别为-0.71 和0.50;2019 年RIL 群体的开花期为30.0 ~58.0 d,标准误差为6.81,峰度和偏度分别为-0.58 和0.59。 RIL 群体两年的开花期分布近似正态分布(图1),且峰度和偏度的绝对值均小于1,表明该群体可用于QTL 定位。
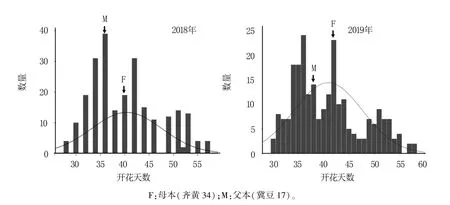
图1 RIL 群体开花期表型数据频率分布

表1 亲本及RIL 群体开花期数据统计(d)
2.2 大豆开花期性状QTL 定位及分析
利用IM-ADD 法对开花期性状进行QTL 定位。 2018 年和2019 年均只检测到1 个位于6 号染色体上的QTL。 两个环境下该QTL 的LOD 值分别为26.81 和21.91,表型贡献率分别为40.04%和33.93%。 该QTL 在两个环境下的加性效应均为负值,表现为负遗传效应(表2、图2)。

图2 不同环境下开花期相关QTL 位点

表2 大豆开花期QTL 定位结果
该QTL 区间内存在36 个基因,其中13 个基因CDS 区在双亲间存在变异(表3)。 与父本冀豆17 相比,Glyma.06G206500 在齐黄34 中发生了移码突变,其它12 个基因均发生了非同义突变。 Uniprot 数据库注释信息以及基因注释分析发现该区间内有两个开花期调控相关基因,其中Glyma.06G205800(GmMDE06)是拟南芥AGL8的同源基因,Glyma.06G207800 是大豆生育期组主效基因E1。 Glyma.06G207800 在齐黄34 中是显性E1基因型,而在冀豆17 中第44 位碱基是“C”,与参考基因组Williams 82 相同,为丧失了部分功能的e1-as基因型。

表3 QTL 位点中CDS 区突变基因
2.3 亲本E1 基因的序列分析和开花促进基因GmFT2a 表达量的检测
对亲本齐黄34 和冀豆17 的E1部分序列进行克隆并测序发现(图3),齐黄34 中的E1基因编码区第44 个核苷酸为G,与显性E1序列完全相同;而冀豆17 的序列在第44 位碱基发生了改变,由G 变成了C,导致编码的氨基酸发生了替换。 证实齐黄34 中E1确实是显性基因型,而冀豆17 中为e1-as基因型。

图3 齐黄34 和冀豆17 E1 部分序列比对
短日处理下,齐黄34 开花期极显著晚于冀豆17(图4A)。 齐黄34 和冀豆17 不同时期叶片中开花促进基因GmFT2a表达量的检测结果(图4B)显示,冀豆17 中GmFT2a的表达量在前期高于齐黄34,且在出苗18 天达到高峰;齐黄34 中GmFT2a的积累滞后于冀豆17,在出苗22 天达到高峰,之后表达量高于冀豆17。

图4 冀豆17 和齐黄34 开花期比较(A)和GmFT2a 表达量(B)比较
3 讨论
齐黄34 是高产广适大豆品种,在黄淮海地区、长江中下游地区以及贵州、广西等低纬度地区均有较好的产量表现。 为挖掘齐黄34 开花期相关QTL,阐明其广适性的遗传机制,本研究以齐黄34 为母本,与大豆品种冀豆17 杂交构建RIL 群体,并进行了开花期性状的QTL 定位。 由于冀豆17 开花早于齐黄34,RIL 群体在开花期表型上存在明显的性状分离;在2018、2019 年两个环境中,均只检测到了1 个QTL(qFT6),位于6 号染色体,表型贡献率最高达到40.04%。 值得注意的是,大豆生育期主效基因E1位于该QTL 区间内。E1基因是大豆光周期响应的核心调控因子,与开花期密切相关。 显性E1基因过量表达将导致大豆开花明显延迟,是十分重要的开花抑制基因。E1的变异类型主要包括e1-fs、e1-nl、e1-as和e1-b3a[11,12],其中,e1-as基因型是E1基因的第44个碱基发生改变,使编码蛋白的定位发生变化[12]。e1-as型植株开花及成熟早于E1基因型植株,也早于功能丧失型(e1-fs及e1-nl)植株。本研究证实E1在齐黄34 中为显性,在冀豆17 中为e1-as基因型。 因此,E1基因可能是解释齐黄34 和冀豆17 衍生的RIL 群体开花表型变异的关键基因,该基因的存在可能是使齐黄34 也适宜在除黄淮海地区以外的日照较短的低纬度地区种植的重要原因。
同时,在该区间内还存在一个开花相关基因GmMDE06(Glyma.06G205800)。 有研究表明Gm-MDE06过量表达能促进大豆开花[19]。 但目前GmMDE06基因单倍型与开花期的相关性尚未得到阐明。 本研究结果显示,该基因在齐黄34 和冀豆17 中有一个氨基酸的差异,因此推测其可能也是造成RIL 群体表型变异的候选基因。
GmFT2a基因在大豆开花调控中起重要作用,是成花素候选基因[20-22]。 在E1过表达植株中,GmFT2a的表达量显著下降[12]。 近期研究表明,E1直接调控GmMDE06进而影响GmFT2a的表达[19]。 本研究中,齐黄34 前期的GmFT2a表达量低于冀豆17,且积累峰值滞后于冀豆17。 可能是由于齐黄34 中显性E1基因对GmFT2a的抑制作用更强,造成了开花促进基因表达量的降低进而显著缩短了其开花期。
本研究在齐黄34 和冀豆17 杂交产生的RIL群体中发现qFT6 位点,也解释了齐黄34 不适宜在高纬度地区种植的原因:主要是因为E1为显性,在日照偏长的高纬度地区对开花促进基因GmFT2a的抑制作用更为强烈,导致齐黄34 开花过晚,甚至不能正常开花。 随着基因编辑技术的快速发展,今后可对齐黄34 中的E1进行定向敲除,使其转变为隐性基因型,进而创制出以齐黄34 为遗传基础但适宜在北方高纬度地区种植的优异大豆种质。
猜你喜欢
中国农学通报(2022年29期)2022-11-25
支部建设(2019年24期)2019-09-28
金桥(2019年12期)2019-08-13
文苑(2019年14期)2019-08-09
现代园艺(2017年21期)2018-01-03
甘肃林业科技(2016年2期)2016-11-16
河南农业大学学报(2016年2期)2016-09-26
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
现代检验医学杂志(2015年5期)2015-02-06
