
巢湖浮游甲壳动物功能群时空变化及其影响因子分析
2023-07-14 01:16刘振宇郭匿春梁阳阳
安徽农业大学学报 2023年3期
张 佳,刘振宇,李 静,郭匿春*,梁阳阳,吴 利
巢湖浮游甲壳动物功能群时空变化及其影响因子分析
张 佳1,刘振宇1,李 静2,郭匿春1*,梁阳阳2,吴 利3
(1. 安徽农业大学资源与环境学院,合肥 230036;2. 安徽省农业科学院水产研究所,水产增养殖安徽省重点实验室,合肥 230001;3. 合肥师范学院生命科学学院,合肥 230061)
为研究巢湖中浮游甲壳动物功能群的时空变化特征及其主要影响因子,在巢湖的主要河口区以及敞水区共设置了20个采样点,对浮游甲壳动物群落结构和水环境因子进行了调查和分析。结果表明:(1)巢湖河口区的8个采样点中共鉴定出枝角类10属21种,桡足类11属16种;敞水区12个点中鉴定出枝角类9属18种,桡足类12属13种;共22属39种。(2)巢湖河口区和敞水区浮游甲壳动物平均密度为530.8 ind·L-1和506.3 ind·L-1,平均生物量分别为14.7 ind·L-1和21.5 ind·L-1;浮游甲壳动物Shannon-Wiener多样性指数显示巢湖秋季河口区的污染程度高于敞水区。(3)巢湖浮游甲壳动物按照体型大小和食性可划分5个功能群,其中,河口区有SCF、LCF共2个优势功能群,敞水区有SCF、LCF、MCF共3个优势功能群,SCF在功能群中占绝对的优势地位。(4)水温是影响河口区和敞水区浮游甲壳动物功能群主要的环境理化因子;巢湖敞水区中浮游甲壳动物LCF功能群能显著影响SCF、MCC、LCC等功能群。巢湖河口区污染程度高于敞水区,巢湖的富营养化治理和藻类水华控制应侧重于河口区的外源污染治理;敞水区可以通过调整浮游甲壳动物功能群结构重建水生生态系统。
巢湖;河口区和敞水区;浮游甲壳动物功能群;环境因子;RDA分析
浮游甲壳动物是水生生态系统的重要组成部分,作为水体中的初级消费者,在水生生态系统的信息、物质和能量传递中发挥重要的作用[1-3]。由于浮游甲壳动物具有能对环境变化迅速做出响应,群落的动态变化能够反应水体的营养状态等独特的生理和生态学特性,因此常被作为环境监测的指示生物[4]。浮游甲壳动物不仅受水温、光照、pH、营养盐、溶解氧、浊度等多种水环境因子的影响[5],同时受浮游植物上行作用、高等水生植物庇护、滤食性鱼类捕食和浮游甲壳动物种内竞争等生物因素的影响,传统浮游甲壳动物的种类分类研究方法难以确切反映其在水生态系统中独特的生态功能[6]。浮游动物功能群研究可较好的明确环境因子对浮游动物群落结构的影响,有助于揭示水生生态系统的结构及食物网关系。霍元子[7]最早研究黄海流域的浮游动物功能群,根据浮游动物的大小、食性、营养级水平以及与高等生物之间的摄食关系等,将黄海浮游动物划分为6个功能群,并建立了基于浮游动物功能群的黄海食物产出的关键过程模型。安睿等根据浮游甲壳动物的大小、摄食习性以及浮游甲壳动物之间的相互作用将黑龙江三江平原的三环泡湿地中的浮游甲壳动物划分为7个功能群,确定了对浮游植物起到主要滤食作用的浮游甲壳动物功能群,同时发现夏季和秋季浮游动物RC优势功能群可以抑制了浮游动物功能群RF对浮游植物的滤食作用[8]。[10, 11]{Ichinokawa, 2006 #188;Le Quere, 2005 #189}{, 2010 #178;Ichinokawa, 2006 #176;Araujo, 2006 #177}[10-12][10-12][10-12]{, 2010 #178;Ichinokawa, 2006 #176;Araujo, 2006 #177}[10-12][10-12]然而,前期国内对于水生生物功能群的研究多集中在浮游植物、底栖动物和鱼类上,而对于淡水浮游甲壳动物功能群的研究相对较少,少数的研究也集中在海洋浮游甲壳动物功能群上[9]。富营养化的淡水水体浮游甲壳动物功能群的研究由于影响因子较多,且生态系统结构较复杂而缺乏全面的研究。富营养化淡水水体浮游甲壳动物功能群的研究对明确水环境中的主导因子,分析水生生态系统的信息、物质和能量变化有积极的意义。
巢湖位于安徽省中部,流域面积约为1.3×104km2,是长江中下游典型的大型浅水湖泊,也是中国五大淡水湖泊之一[10]。近30年来,随着人类活动加剧,巢湖水体营养盐和有机质浓度增加,造成了巢湖湖泊富营养化进程加快,水质恶化和蓝藻水华频繁暴发[11],因此引发了越来越多地关注[12]。对于巢湖水体中浮游生态系统的研究由来已久,颜京松等[13]最早研究了浮游甲壳动物群落结构,共发现枝角类10种,桡足类9种,作为重要的鱼类饵料生物,浮游甲壳动物在巢湖水生生态系统能量流转和食物链网变化中发挥着重要作用。邓道贵[5, 6]研究了巢湖水体中的浮游甲壳动物空间分布、季节演替以及历史变化,发现巢湖春季优势种为大型枝角类,冬季为桡足类中的中华窄腹剑水蚤,夏秋季为小型枝角类,小型枝角类密度与优势蓝藻生物量呈正相关,浮游甲壳动物群落结构主要受上行效应(高营养盐水平)以及透明度、水温等物理因素影响[14]。王凤娟等曾用浮游动物污生指数法评价巢湖东湖区的水质状况,用浮游动物数量(ind‧L-1)作为湖泊水体营养程度的生物量指标,研究发现巢湖东湖区水体在2005年6—9月和2006年3—6月期间的营养状况为富营养状态[15]。陈倩颖等系统分析了巢湖浮游动物功能群和浮游植物的组成特征的关系,发现浮游植物密度可以影响浮游动物功能群,且微囊藻是与巢湖浮游动物功能群相关性最强的环境因子[16]。由此可见,浮游动物功能群的研究对了解巢湖水生生态系统的能量流转、食物链网变化、营养状态以及藻类控制上均有积极意义。近年来,巢湖的营养负荷处于高位波动状态,水体富营养化程度也进一步加深,然而对此过程中的浮游甲壳动物功能群时空变化及其影响因子进行研究,可以揭示富营养化过程中浮游甲壳动物的作用并为富营养化的控制提供理论支持。
本研究通过对巢湖河口及敞水区20个采样点、4个季节的浮游甲壳动物群落结构和功能群进行调查和分析,以期了解巢湖浮游甲壳动物功能群的时空变化特征,探究水环境因子对浮游甲壳动物群落结构和功能群的影响。
1 材料与方法
1.1 研究地点与采样点的设置
按照《全国湖泊富营养化调查规范(第二 版)》[17],根据巢湖的自然环境特点,共设置了20个采样点(图1)。其中巢湖主要河口设置了8个采样点E1—E8,敞水区设置了12个采样点P1—P12。河口区采样点E1至E8依次对应十五里河、派河、南淝河、杭埠河、白石天河、马尾河、柘皋河、双桥河8条入湖河流。本研究在2020年8月到2021年4月开展为期一年的湖泊生态监测和调查。分别在2020年8月(夏季)、2020年11月(秋季)、2021年2月(冬季)和2021年4月(春季)对全湖进行4次样品采集与分析。
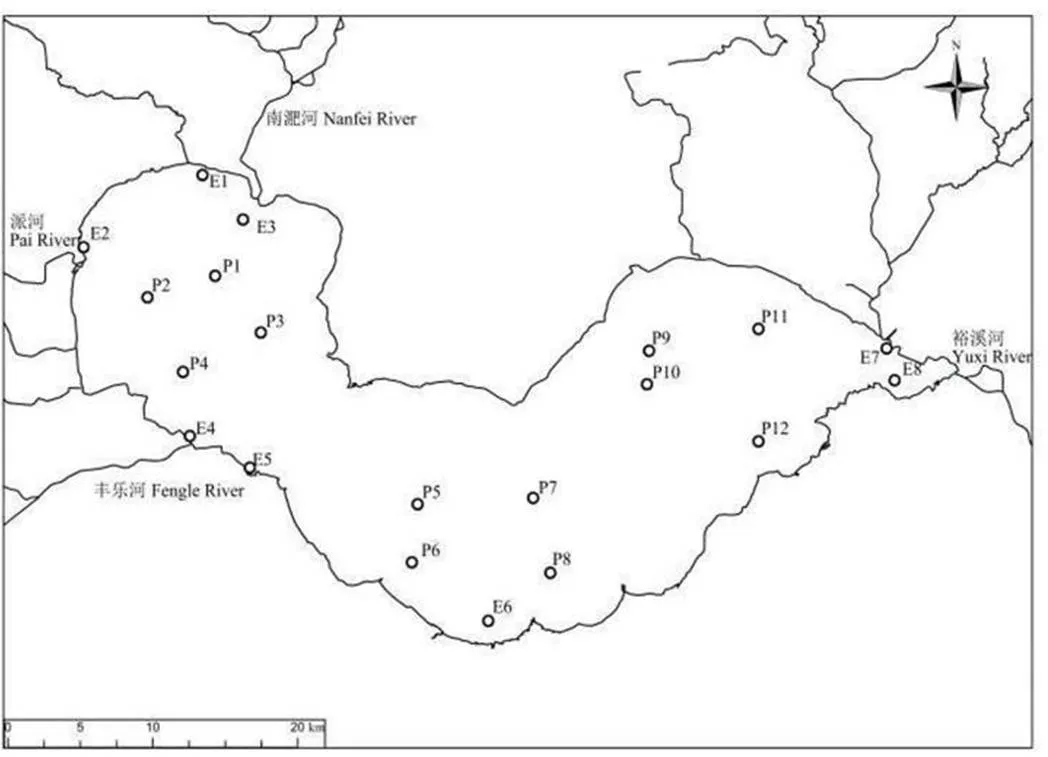
图1 巢湖采样点设置图
Figure 1 Distribution of sampling sites in Chaohu Lake
1.2 水质理化指标分析方法
水体透明度采用Secchi盘现场测定,WT、pH、DO等使用YSI6600V2型多参数水质监测仪现场测定。水体理化指标总氮(TN)、总磷(TP)、铵态氮(NH+4-N)、硝态氮(NO3--N)等浓度等参照《水和废水监测分析方法》在实验室内测定[18]。
1.3 浮游生物采样方法和样品处理
浮游甲壳动物定性样品采集时用13号浮游生物网在河水中反复做“∞”字型划水,将其放入50 mL的塑料小方瓶中,加入4%的甲醛溶液固定保存。浮游甲壳动物定量样品采集时用5 L的采水器在采样点取10 L水样,之后用25号浮游生物网过滤,将其放入50 mL塑料小方瓶中,用4%的甲醛溶液固定保存。每个季度在20 个采样点各采集一次,将样品带回实验室处理。定性及定量样品均在10×4倍的显微镜下用5 mL浮游生物计数板进行观察和计数。其中,浮游甲壳动物枝角类和桡足类按照《中国动物志(淡水枝角类)》[19]和《中国动物志(淡水桡足类)》[20]鉴定.浮游甲壳动物生物量的估算参照陈雪梅[21]、范正年[22]等学者的方法。
1.4 数据处理与分析
浮游甲壳动物的多样性研究采用多样性指数Shannon-Wiener指数、丰富度指数Margalef指数和优势度指数:
优势度指数():
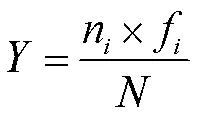
Shannon-Wiener多样性指数:

Margalef丰富度指数:
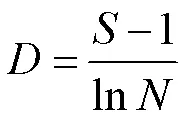
式中,为物种数目,值越大,多样性越高,n为物种的密度,f为物种出现的频率,为所有物种的总密度,当>0.02时,该物种为群落的优势种[23]。水质判断标准为:′ >3:清洁;1~3:中污(其中1~2为-中污,2~3为-中污);0~1:重污。>3:清洁;2~3:轻污;1~2:中污;0~1:重污[24-25]。实验数据经Microsoft Excel 2010、SPSS 20.0、Canoco 5.0(环境因子数据进行lg(+1)转化)软件分析。
2 结果与分析
2.1 巢湖水环境特征
采样期间,全湖水温变化范围为7.4~30.9 ℃,透明度变化范围为15.0~60.0 cm,溶解氧变化范围为1.49~12.70 mg·L-1,pH全年的平均值为7.88,整体呈现为弱碱性水体。不同季节以及不同采样点水环境理化指标有所差异,河口区TN最高为5.48 mg·L-1,最低为1.92 mg·L-1;NH4+-N最高为1.04 mg·L-1,最低为0.32 mg·L-1;NO3--N最高为2.79 mg·L-1,最低为1.07 mg·L-1;NO2--N最高为0.13 mg·L-1,最低为0.03 mg·L-1;TP最高为0.45 mg·L-1,最低为0.15 mg·L-1;CODMn最高为65.00 mg·L-1,最低为24.00 mg·L-1,敞水区TN最高为3.64 mg·L-1,最低为0.12 mg·L-1;NH4+-N最高为3.99 mg·L-1,最低为0.12 mg·L-1;NO3--N最高为2.29 mg·L-1,最低为mg·L-1;NO2--N最高为0.07 mg·L-1,最低为0.00 mg·L-1;TP最高为0.30 mg·L-1,最低为0.02 mg·L-1;CODMn最高为44.00 mg·L-1,最低为1.00 mg·L-1。从平均值大小来看,除NH4+-N以外,其余5项营养盐指标均呈现出河口区大于敞水区的现象。

表1 巢湖河口区与敞水区水环境理化因子
2.2 浮游甲壳动物种类、密度、生物量和生物多样性指数分析
在巢湖的20个采样点中,共鉴定出枝角类10属22种,桡足类12属17种,总计22属39种,枝角类种类居多,占总种类数的56.41%。其中河口区8个采样点中枝角类共计10属21种,桡足类11属16种;敞水区12个点中鉴定出枝角类9属18种,桡足类12属13种。整体来看,巢湖河口区的浮游甲壳动物种类组成与敞水区相比较为丰富,但是数量差异不明显。巢湖河口区和敞水区浮游甲壳动物平均密度为530.8 ind·L-1和506.2 ind·L-1,平均生物量分别为14.7 mg·L-1和21.5 mg·L-1。在4个季度的20个采样点中,河口区E4夏季密度最高,为334.8 mg·L-1;敞水区P12夏季密度最高,为334.8 mg·L-1;敞水区P12夏季密度最高,为410.9 ind·L-1;河口区E6夏季生物量最大,为4.8 mg·L-1;敞水区P11夏季生物量最大,为3.0 mg·L-1。根据多样性指数评价标准及巢湖浮游甲壳动物多样性指数变化,从Shannon-Wiener指数整体上看河口区和敞水区均处于中度污染状态,其中河口区夏季和秋季为α-中污,春季和冬季为β-中污,敞水区仅夏季为α-中污,其余季节都为β-中污;从丰富度Margalef指数看,无论是河口区还是敞水区只有夏季为重污染状态,其余季节均为中污染程度。
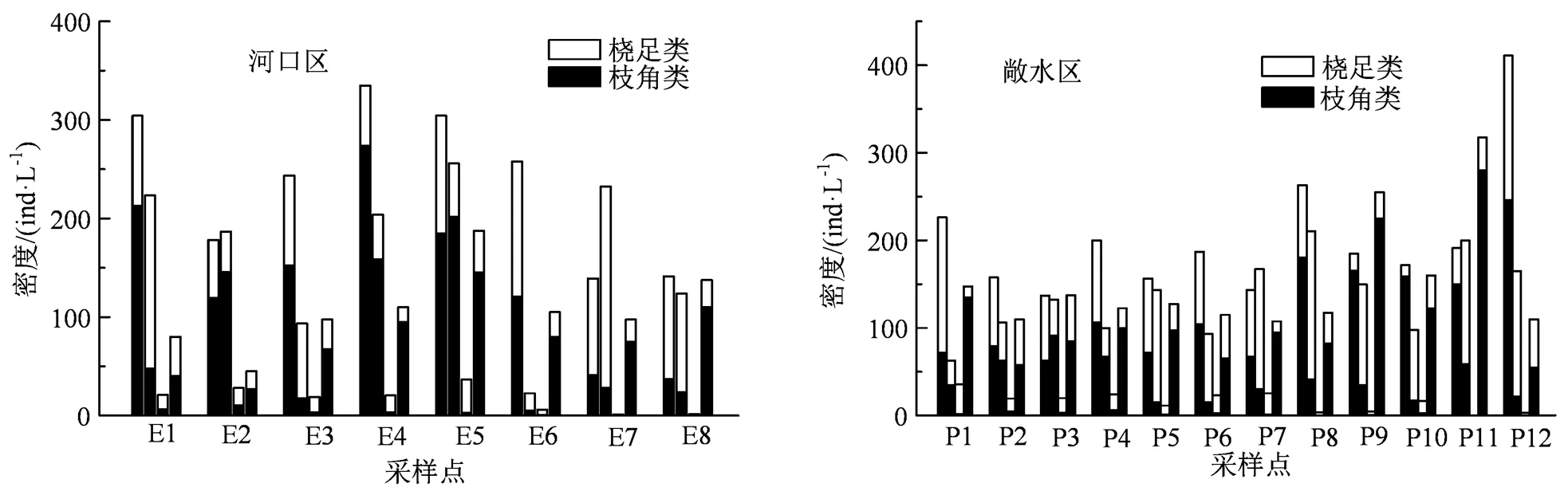
各采样点从左至右依次为夏季、秋季、冬季、春季,下同。
Figure 2 Density of planktonic crustaceans in Chaohu estuary and pelagic area
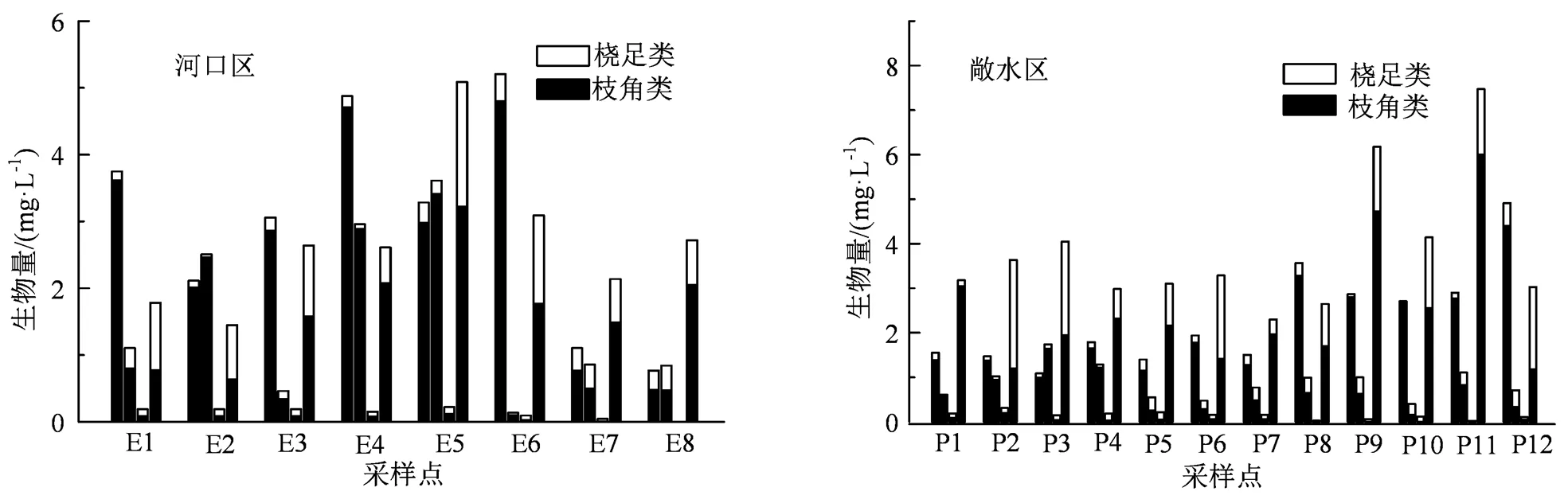
图3 巢湖河口区与敞水区浮游甲壳动物的生物量
Figure 3 Biomass of planktonic crustaceans in Chaohu estuary and pelagic area
2.3 浮游甲壳动物功能群划分和时空变化分析
根据体型大小和食性将巢湖浮游甲壳动物共划分出5个功能群(表3),分别为小型浮游甲壳动物滤食者SCF、中型浮游甲壳动物滤食者MCF、中型浮游甲壳动物捕食者MCC、大型浮游甲壳动物滤食者LCF和大型浮游甲壳动物捕食者LCC,LCC功能群仅在冬季没有出现。河口区浮游甲壳动物优势功能群主要为SCF、LCF功能群,敞水区游甲壳动物优势功能群主要为SCF、LCF、MCF功能群,其中SCF功能群优势度最大。此外,河口区和敞水区各功能群的生物量存在明显的季节变化,不同季节各功能群所占比例不同。河口区夏季的优势功能群是SCF功能群,超过70%;秋季、冬季、春季的优势功能群均是SCF+LCF功能群,其中尽管SCF功能群仍然占主体地位,但LCF功能群的比例逐渐增大。敞水区夏季的优势功能群为SCF功能群,且高于河口区,约占85%;秋季的优势功能群为SCF+MCC;冬季为SCF+LCF功能群;春季与夏季功能群相同,都是SCF+MCF功能群,其中夏季、秋季和春季SCF功能群都超过总数的一半,秋季SCF功能群和LCF功能群所占的比例为30.61%和29.64%,较为接近。

表3 巢湖浮游甲壳动物功能群划分
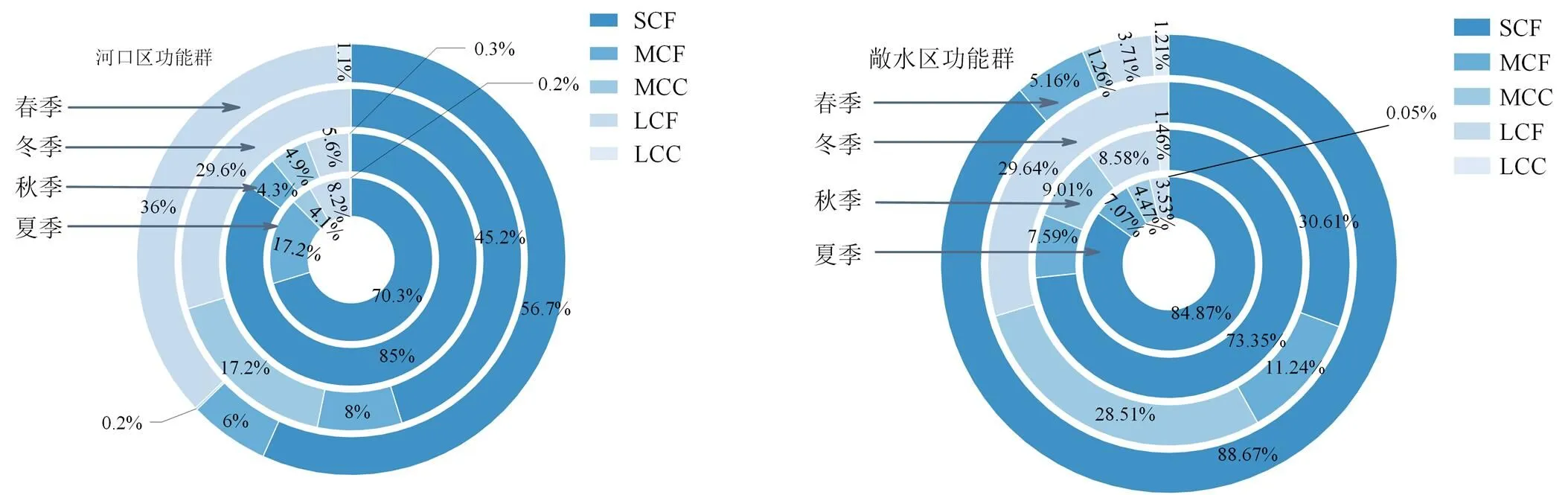
图4 巢湖河口区和敞水区不同季节浮游甲壳动物功能群对比
Figure 4 Comparison of functional groups of planktonic crustaceans in different seasons in Chaohu estuary and pelagic area
2.4 浮游甲壳动物功能群的作用关系及其与水环境因子的关系
2.4.1 不同浮游甲壳动物功能群同其他浮游甲壳动物功能群及水环境因子的Pearson相关分析 研究发现,功能群与环境因子有密切联系。河口区SCF功能群与WT(=0.361,<0.05)、CODMn(=0.389,<0.05)呈显著正相关;MCF功能群与WT(=0.650,<0.01)、pH(=0.473,<0.01)呈极显著正相关;MCC功能群和WT(=0.577,<0.01)呈极显著正相关,与TP(=0.414,<0.05)呈显著正相关。敞水区SCF功能群与WT(0.471,<0.01)、CODMn(=0.468,<0.01)呈极显著正相关,与NO3--N(=-0.471,<0.01)呈极显著负相关,与pH(=0.325,<0.05)呈显著正相关;MCF功能群与WT(=0.524,<0.01)呈极显著正相关,与SD(=-0.303,<0.05)呈显著负相关,pH(=0.339,<0.05)、CODMn(=0.296,<0.05)呈显著正相关;MCC功能群与WT(=0.414,<0.01)、TP(=0.373,<0.01)呈极显著正相关,与NO3--N(=-0.368,<0.05)呈显著负相关;LCF功能群与SD(=-0.367,<0.05)、TN(=-0.315,<0.05)呈显著负相关,与CODMn(=0.478,<0.01)呈极显著正相关。不同功能群之间也存在影响,在河口区SCF功能群和MCC功能群(=0.417,<0.05)呈显著正相关,MCC功能群与LCF功能群(=-0.416,<0.05)呈显著负相关。敞水区SCF功能群与LCF功能群(=0.394,<0.01)呈极显著正相关,与MCF功能群(=0.292,<0.05)呈显著正相关;LCF功能群与MCC功能群(=-0.378,<0.01)呈极显著负相关,与LCC功能群(=0.379,<0.01)呈极显著正相关。

表4 不同浮游甲壳动物功能群同水环境因子的Pearson相关性分析
注:*在0.05水平显著相关;**在0.01水平上显著相关。下同。
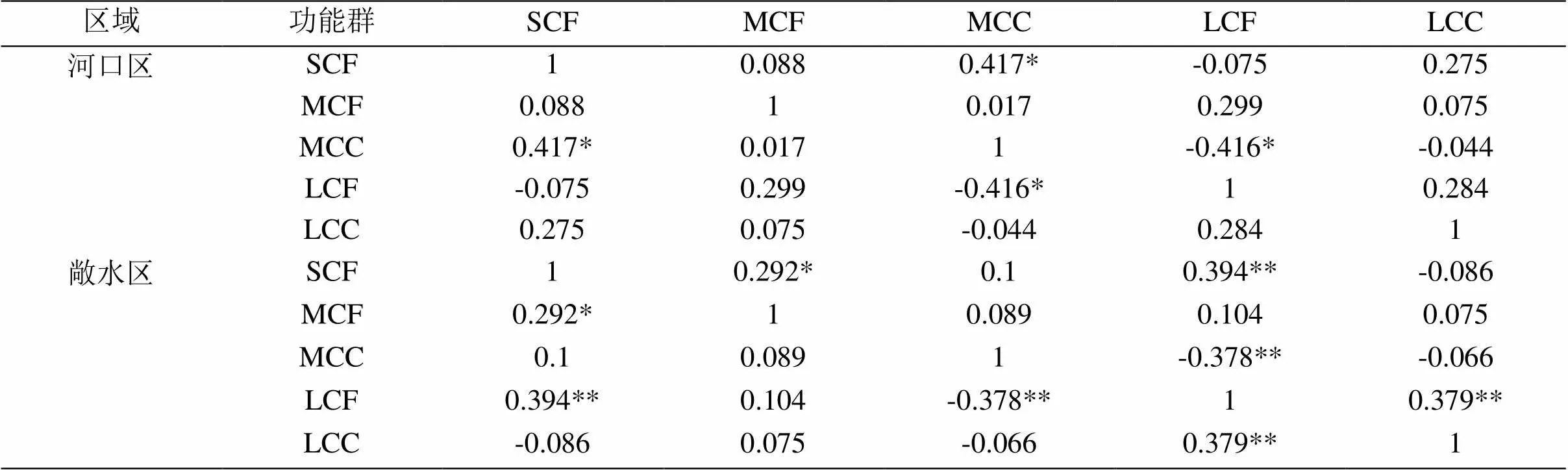
表5 不同浮游甲壳动物功能群同其他浮游甲壳动物功能群的Pearson相关分析
2.4.2 浮游甲壳动物功能群同水环境因子的 RDA 分析 对数据进行去趋势对应分析(DCA),结果显示排序轴长度<3,故对浮游动物功能群与环境因子间的关系采用冗余分析(RDA)。河口区第一轴与第二轴的特征值分别为0.202 9和0.138 7,第一轴解释了物种信息的20.29%,第二轴进一步解释了物种信息的13.87%,前两轴共解释了34.16%的物种信息与82.44%的物种-环境信息。表明在河口区环境因子可较好的解释与浮游甲壳动物功能群之间的关系。敞水区第一轴与第二轴的特征值分别为0.196 5和0.147 7,第一轴解释了物种信息的19.65%,第二轴进一步解释了物种信息的14.77%,前两轴共解释了34.42%的物种信息与77.95%的物种-环境信息,同样表明在敞水区浮游甲壳动物功能群与环境因子之间存在明显的相关性。冗余分析(RDA)发现,前两轴对河口区和敞水区物种-环境信息解释率相似,表明在富营养化水体中水质理化因子对浮游甲壳动物功能群的解释度较为接近,不能完全区别河口区和敞水区的差异。
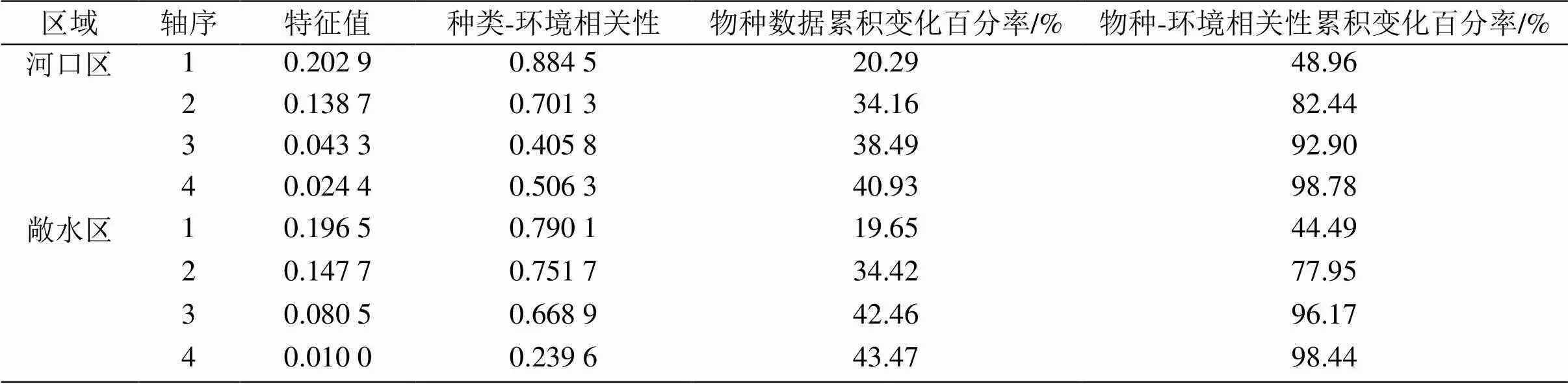
表6 浮游甲壳动物功能群与水环境因子之间的RDA分析结果
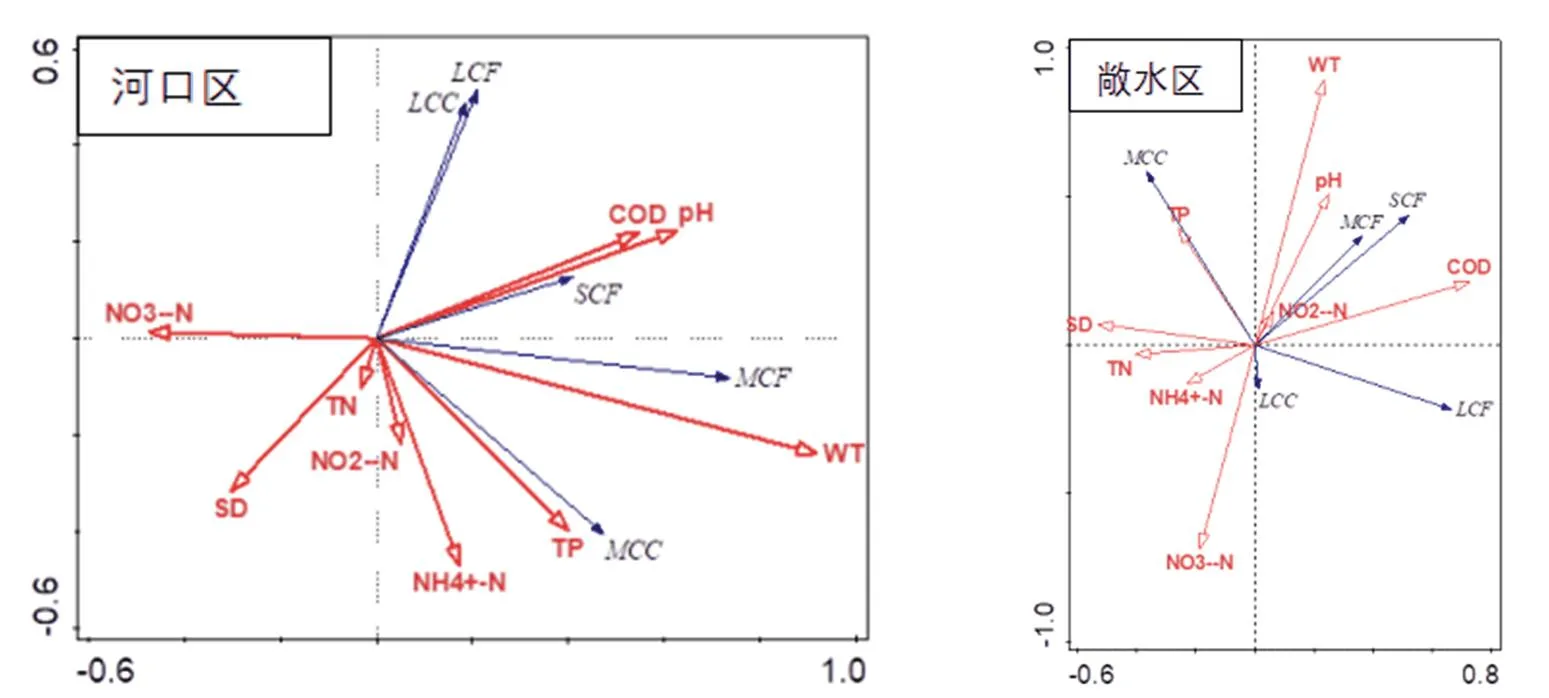
图5 浮游甲壳动物与环境因子的RDA分析
Figure 5 RDA of zooplankton and environmental factors
3 讨论
3.1 巢湖河口区和敞水区的环境特征比较
本研究发现巢湖河口区和敞水区TN>1 mg·L-1、TP>0.1 mg·L-1,均已达到水体富营养化水平,巢湖河口区和敞水区的WT、SD和pH均在正常范围内波动且相差不大。同时,巢湖河口区的TN、TP、NO3--N、NO2--N和CODMn这5项指标均比敞水区高,只有NH4+-N敞水区略高于河口区。其中,巢湖派河河口春季的E2采样点TN含量高达5.48 mg·L-1,E8(双桥河)秋季的TP含量最高为0.45 mg·L-1,表明河口区污染程度高于敞水区,巢湖TP浓度、TN浓度与2018年张民等[26]的研究结果接近,河口区TP浓度较2018年增加20%,敞水区下降了12%,TP浓度是影响巢湖优势水华蓝藻鱼腥藻和微囊藻两种藻时空分布的主要因素之一[27],这也是巢湖水华暴发的重要原因;TN与2018年相比,均呈下降趋势,河口区下降12%,敞水区下降41%,氮含量的降低会使水体富营养化的风险减少[28]。与此同时,河口区的N/P也高于敞水区,N/P作为水体营养结构的主要指标,当N/P在10~23之间时易造成水体水华暴发[29],河口区(12.80)和敞水区(11.55)均处于这个阶段。研究结果显示,巢湖的富营养化水平维持在较高水平波动,西部入湖河流污染是造成的巢湖富营养化的主要原因,湖泊外源污染负荷主要来源于流域西北部。入湖河流作为氮和磷转移到湖泊中的主要路径,集中了所在流域的各种点源污染(工业污染、生活污水等)和面源污染(农业生产污水)[30],其中农田对全流域面源TN和TP污染输出的贡献分别为81%和52%,化肥施用强度高依然是流域面源污染的核心问题[31],同时入湖河流在很大程度上也受到了底泥向水体释放的氮、磷营养盐的影响;敞水区各项指标较河口区低可能是与湖泊对氮磷的沉积与吸附有关[32]。鉴于巢湖河口区污染程度高于敞水区,河口区是外源污染的主要来源,因此巢湖的外源污染是造成巢湖富营养化程度高,水质恶化的主要原因。通过消减外源污染负荷,切断其污染途径是巢湖富营养化治理的重要措施。近年来,巢湖周边实施的大量工程如引江济巢工程通过消减外源入湖污染、增加江湖水量交换、缩短巢湖水体置换时间可以有效降低巢湖河口区营养负荷和减弱外源污染,进而达到改善湖泊水环境质量和抑制蓝藻大规模暴发的目的[33]。
3.2 巢湖河口区和敞水区浮游甲壳动物群落结构比较
研究发现巢湖浮游甲壳动物22属39种,河口区21属38种,敞水区21属31种。河口区的浮游甲壳动物种类数与敞水区相比较为丰富;从浮游甲壳动物优势种类及其占比上看,巢湖水体中优势种类为枝角类中个体较小的长额象鼻溞, 桡足类主要以无节幼体形式存在,优势种组成均与李静等[34]2015年的研究结果相似,表明巢湖近几年浮游动物群落结构未发生明显变化。河口区和敞水区浮游甲壳动物平均密度为530.8 ind·L-1和506.2 ind·L-1,平均生物量分别为14.7 mg·L-1和21.5 mg·L-1,巢湖河口区浮游甲壳动物密度高于敞水区但生物量低于敞水区,表明河口区浮游甲壳动物的体型小于敞水区,河口区作为外源污染物的主要来源,反映出了污染程度高时,浮游甲壳动物小型化程度也越高。本研究由于未对原生动物以及轮虫进行检测与分析,无法使用王凤娟等[15]2006年提出的利用浮游动物数量(ind·L-1)作为湖泊水体营养程度的评价方法,有待在后续的研究中进行补充。Margalef多样性指数显示巢湖夏季的污染程度更高,这可能是由于温度较高时,一些小粒径藻类大量生长,为浮游甲壳动物提供了充足的食物来源;夏季水华暴发,蓝藻浓度高但由于一些小型浮游动物长期与蓝藻共存时,产生了一定适应性[35],因此小型浮游甲壳动物种类数在总种数中所占比例较高, 大型浮游甲壳动物种类数数占比例较低,这与邓道贵等[14]的研究结果一致,体型较大的个体随藻类丰度的上升而下降,小型种类则呈现上升趋势。Shannon-Wiener多样性指数表明河口区秋季的污染程度高于敞水区。进入秋季,由于降水等的减少导致河流域内水流量减少、流速缓慢,水体扰动性减弱,使得整个水体环境相对稳定,加上河岸带植物开始凋零,随着枯枝落叶的腐败、降解,增加了水体中的营养物质,适合的水体温度加上丰富的营养物质影响了群落多样性[36],因而河口区多样性指数高于敞水区。
3.3 巢湖河口区和敞水区的浮游甲壳动物功能群的时空变化分析
与巢湖整体的富营养化状态一致,巢湖浮游甲壳动物优势功能群主要为SCF功能群,其中小型浮游甲壳动物滤食者SCF功能群占比超过50%,这也造成了巢湖水体中浮游动物滤食效率低下[37],下行作用较弱的现状。同时,敞水区夏秋季节出现了较多的MCC和MCF功能群;这可能与敞水区富营养程度较弱,浮游甲壳动物的生态功能更强有关。本研究中巢湖浮游甲壳动物各功能群存在明显的时空变化特征,不同季节和不同区域各功能群所占比例不同。巢湖夏季优势功能群为SCF功能群,主要包括枝角类的尖额溞、象鼻溞和盘肠溞等;随着水温的升高,枝角类和桡足类会逐渐增多,出现这种情况的原因有可能是一些小型种类在夏季的生长速率比较高,繁殖加快所致;夏季是蓝藻暴发的主要时期,水体的富营养化则会导致浮游动物小型化,因此会引起小型种类的增加[38]。敞水区秋季是SCF+MCC功能群,与夏季不同的是优势功能群出现了中型浮游甲壳动物捕食者,主要是桡足类的广布中剑水蚤、透明温剑水蚤和胸饰外剑水蚤,MCC功能群主要由广布中剑水蚤组成,有研究表明广布中剑水蚤在水温15~20 ℃时的繁殖能力最强[39],这可能是因为夏季和秋季的浮游植物密度较大,功能群MCC主要以功能群MCF为食,而功能群MCF主要以藻类为食,抑制了MCF功能群的增长,所以夏季和秋季的MCC较多;河口区冬春季节出现了LCF功能群,主要是汤匙华哲水蚤,湖泊中由于大型滤食者LCF的存在,春季常常出现大型滤食者滤食作用强,藻类生物量受到控制的“清水期”,而夏秋季节则由于SCF功能群占据优势地位,滤食效率下降,而造成藻类水华的暴发。有研究表明,枝角类适于生长在温度不高的水体中[40-41],所以在温度较低的春冬季SCF功能群的生物量仍占优势。因此富营养化水体全年以SCF功能群占优势,这与陈倩颖等的研究结果一致[16]。敞水区夏秋季节出现了MCC和MCF功能群属于中型浮游甲壳动物滤食者和捕食者,与河口区的SCF功能群相比,中型滤食者的摄食效率更高,对浮游植物的控制能力也更强。因此,敞水区浮游甲壳动物的下行作用更强,对藻类的控制能力也高于河口区。浮游甲壳动物功能群研究解释了巢湖的富营养化特征及藻类控制的关键因子。
3.4 巢湖河口区和敞水区的浮游甲壳动物功能群的主要影响因子
本次研究经RDA分析显示,河口区前两轴共解释了34.16%的物种信息与82.44%的物种-环境信息,敞水区前两轴共解释了34.42%的物种信息与77.95%的物种-环境信息,河口区和敞水区解释度差异不大,不能完全解释河口区和敞水区影响因子的差异。环境因子中仅温度与巢湖浮游甲壳动物相关性较高,其他理化因子与浮游甲壳动物功能群的相关性均不高,这与吴利[34]等的研究结果一致,富营养化的淡水水体浮游甲壳动物功能群的研究由于影响因子较多,对功能群影响最大的是WT,说明不仅温度会影响浮游甲壳动物功能群,同时还有其他因素,比如N、P等对浮游甲壳动物功能群的结构与分布也存在着不可忽视的影响[42]。Pearson分析发现,除环境因子的影响外,各功能群之间也具有明显的相关性,这是由于各功能群之间存在食物竞争和捕食关系,浮游甲壳动物之间的竞争能力不仅体现在影响其竞争者利用相同资源的能力,也体现在能忍受有限资源不断降低的情况,竞争能力的强弱大多取决于浮游甲壳动物本身的大小[43],这也是浮游甲壳动物功能群时空存在异质性的一个十分重要的因素。目前国内已有许多对水环境因子与水生态系统中浮游甲壳动物影响的研究[44-46],浮游甲壳动物间的相互作用的研究相对较少。浮游甲壳动物的季节演替主要受温度、营养盐和浮游植物的种类变化而变化,浮游甲壳动物的种类变化以及生物量的大小都会受到种间竞争、鱼类捕食等关系的影响。巢湖敞水区中浮游甲壳动物LCF功能群能显著影响SCF、MCC、LCC等功能群,这可能表明敞水区浮游甲壳动物有较强的种间作用,LCF优势功能群可以影响其他功能群对浮游植物的滤食作用,与安睿[8]等发现夏季和秋季浮游动物RC优势功能群可以抑制了浮游动物功能群RF对浮游植物的滤食作用相似,本研究中LCF功能群也能影响MCC功能群浮游植物的滤食作用。因此,调整巢湖敞水区浮游动物功能群结构和重建水生生态系统可以在治理富营养和控制藻类水华发挥积极作用。
4 结论
巢湖TN、TP浓度均达到富营养化水平,且河口区TN、TP、NO3--N、NO2--N和CODMn这5项指标均高于敞水区。研究表明河口区污染程度高于敞水区,外源污染经河口区入巢湖是目前巢湖污染的主要来源,巢湖的富营养化治理应优先侧重于河口区的外源污染治理。
巢湖浮游甲壳动物群落及功能群能较好表征巢湖的富营养化水平。Shannon-Wiener指数显示夏季巢湖河口区的污染程度较高;巢湖河口区浮游甲壳动物密度高于敞水区,生物量却低于敞水区,浮游甲壳动物趋于小型化,也说明河口区污染程度高于敞水区。巢湖浮游甲壳动物优势功能群主要为与富营养化状态一致的SCF功能群,敞水区夏秋季节出现了较多的MCC和MCF功能群;这可能与敞水区富营养程度较弱,浮游甲壳动物的生态功能更强有关。
环境因子中仅温度与巢湖浮游甲壳动物相关性较高,而巢湖敞水区中浮游甲壳动物LCF功能群能显著影响SCF、MCC、LCC等功能群,这可能表明敞水区浮游甲壳动物有较强的种间作用,LCF优势功能群可以影响其他功能群对浮游植物的滤食作用。因此,调整巢湖敞水区浮游动物功能群结构和重建水生生态系统可以在治理富营养和控制藻类水华发挥积极作用。
[1] LAMPERT W. Zooplankton research: the contribution of limnology to general ecological paradigms[J]. Aquat Ecol, 1997, 31(1): 19-27.
[2] 余居华,钟继承,张银龙,等.湖泊底泥疏浚对沉积物再悬浮及营养盐负荷影响的模拟[J].湖泊科学, 2012, 24(1): 34-42.
[3] 钟继承,刘国锋,范成新,等.湖泊底泥疏浚环境效应:Ⅰ.内源磷释放控制作用[J].湖泊科学,2009,21(1):84-93.
[4] 王卫成. 胶州湾浮游动物功能群长期变化研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所), 2017.
[5] 陆娇. 湘江长沙综合枢纽上下游浮游生物群落结构动态分布特征及水质评价[D]. 长沙: 湖南师范大学, 2019.
[6] HOOD R R, LAWS E A, ARMSTRONG R A, et al. Pelagic functional group modeling: progress, challenges and prospects[J]. Deep Sea Res Part II, 2006, 53(5/6/7): 459-512.
[7] 霍元子. 黄海浮游动物功能群的研究[D]. 青岛: 中国科学院研究生院(海洋研究所), 2008.
[8] 安睿.三环泡湿地浮游动物功能群对浮游植物功能群的影响[J]. 防护林科技, 2018(8): 56-59.
[9] SUN S, HUO Y Z, YANG B. Zooplankton functional groups on the continental shelf of the Yellow Sea[J]. Deep Sea Res Part II, 2010, 57(11/12): 1006-1016.
[10] 马荣华,杨桂山,段洪涛,等.中国湖泊的数量、面积与空间分布[J].中国科学:地球科学,2011,41(3):394-401.
[11] 张民,孔繁翔.巢湖富营养化的历程、空间分布与治理策略(1984-2013年)[J].湖泊科学,2015,27(5):791-798.
[12] 宁怡, 高峰, 邓建才, 等. 巢湖流域水质生物学评价: 以大型底栖动物为例[J]. 生态学杂志, 2012, 31(4): 916-922.
[13] 颜京松,张玉书,王美珍.巢湖鱼产潜力估算及合理开发利用的一些建议[J].农村生态环境,1987(2):34-41.
[14] 邓道贵. 大型浅水湖泊—巢湖的富营养化对浮游生物影响的生态学研究[D]. 武汉: 中国科学院水生生物研究所, 2004.
[15] 王凤娟, 胡子全, 汤洁,等.用浮游动物评价巢湖东湖区的水质和营养类型[J]. 生态科学, 2006, 25(6):550-553.
[16] 陈倩颖,吴利.巢湖浮游动物功能群和浮游植物的相互关系[J].安徽理工大学学报(自然科学版), 2021, 41(4): 42-50.
[17] 金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 2版. 北京: 中国环境科学出版社, 1990.
[18] 国家环保总局. 水和废水监测分析方法[M]. 北京: 中国环境科学出版社, 2006: 354.
[19] 蒋燮治, 堵南山. 中国动物志,节肢动物门,甲壳纲,淡水枝角类[J]. 北京:科学出版社, 1979.
[20] 中国科学院中国动物志委员会. 中国动物志,节肢动物门甲壳纲,淡水桡足类[M]. 北京:科学出版社, 1999.
[21] 陈雪梅.淡水桡足类生物量的测算[J].水生生物学集刊, 1981, 7(3): 397-408.
[22] 范正年.测定淡水枝角类生物量的两种方法比较[J].动物学杂志,1988,23(5): 32-34.
[23] 高子涵,张健,皮杰,等.湖南省大通湖浮游动物群落结构及其与环境因子关系[J].生态学杂志, 2016, 35(3): 733-740.
[24] 王宏伟,张蕾颖,沈公铭,等.拒马河底栖动物多样性及其水质评价[J].河北大学学报(自然科学版), 2007, 27(5): 530-536.
[25] 张婷,马行厚,王桂苹,等.鄱阳湖国家级自然保护区浮游生物群落结构及空间分布[J].水生生物学报, 2014, 38(1): 158-165.
[26] 张民,史小丽,阳振等.2012—2018年巢湖水质变化趋势分析和蓝藻防控建议[J].湖泊科学,2020,32(1):11-20.
[27] ZHANG M, ZHANG Y C, YANG Z, et al. Spatial and seasonal shifts in bloom-forming cyanobacteria in Lake Chaohu: patterns and driving factors[J]. Phycol Res, 2016, 64(1): 44-55.
[28] CAMARGO J A, ALONSO Á. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: a global assessment[J]. Environ Int, 2006, 32(6): 831-849.
[29] ZHANG Y, SONG C L, JI L, et al. Cause and effect of N/P ratio decline with eutrophication aggravation in shallow lakes[J]. Sci Total Environ, 2018, 627: 1294-1302.
[30] 朱俊,董辉,王寿兵,等.长江三峡库区干流水体主要污染负荷来源及贡献[J].水科学进展,2006,17(5):709-713.
[31] 高田田, 谢晖, 万能胜, 等. 巢湖典型农村流域面源氮磷污染模拟及来源解析[J]. 农业环境科学学报, 2022, 41(11): 2428-2438.
[32] 张路, 范成新, 王建军, 等. 长江中下游湖泊沉积物氮磷形态与释放风险关系[J]. 湖泊科学, 2008, 20(3): 263-270.
[33] 菅浩然, 韩涛. 引江济巢工程对改善巢湖水质的数值模拟分析[J]. 水电能源科学, 2020, 38(9): 53-55.
[34] 吴利,周明辉,沈章军,等.巢湖及其支流浮游动物群落结构特征及水质评价[J].动物学杂志,2017,52(5):792-811.
[35] 王梦梦, 张玮, 杨丽, 等. 上海市河道水体拟浮丝藻生物量与环境因子的回归分析[J]. 生态学杂志, 2018, 37(1): 187-193.
[36] 杨清,张鹏,安瑞志,等.拉萨河中下游纤毛虫群落时空分布模式及其驱动机制[J].生物多样性, 2022, 30(6): 136-150.
[37] 邱东茹,吴振斌.生物操纵、营养级联反应和下行影响[J].生态学杂志, 1998, 17(5): 27-32.
[38] 杨宇峰, 王庆, 陈菊芳, 等. 河口浮游动物生态学研究进展[J]. 生态学报, 2006, 26(2): 576-585.
[39] 高原, 赖子尼, 曾艳艺, 等. 珠江三角洲河网桡足类群落结构及其与水环境因子的关系[J]. 中国水产科学, 2015, 22(2): 302-310.
[40] 徐梅, 吴芳仪, 刘靓靓, 等. 焦岗湖浮游甲壳动物群落结构的季节动态[J]. 生态学杂志, 2016, 35(5): 1254-1262.
[41] 杨亮杰, 吕光汉, 竺俊全, 等. 横山水库浮游动物群落结构特征及水质评价[J]. 水生生物学报, 2014, 38(4): 720-728.
[42] 刘忱, 黄燕, 刘瑜, 等. 东苕溪夏季浮游动物群落结构及水环境的初步研究[J]. 杭州师范大学学报(自然科学版), 2015, 14(5): 498-506.
[43] BRÖNMARK C, HANSSON L A. The Biology of Lakes and Ponds[M]. Oxford: Oxford University Press, 2017.
[44] 李秋华, 温远志, 杨卫诚, 等. 贵州百花湖麦西河河口后生浮游动物群落结构及与环境因子的关系[J]. 湖泊科学, 2013, 25(4): 531-538.
[45] 苗滕, 高健, 陈炳辉, 等. 惠州西湖生态修复对浮游甲壳动物群落结构的影响[J]. 生态科学, 2013, 32(3): 324-330.
[46] 王博, 廖剑宇, 刘全儒, 等. 东江干流浮游动物群落结构特征及与水质的关系[J]. 北京师范大学学报(自然科学版), 2013, 49(6): 608-612.
Spatial-temporal variations of planktonic crustacean functional groups and analysis for the impact factors in Chaohu Lake
ZHANG Jia1, LIU Zhenyu1,LI Jing2,GUO Nichun1, LIANG Yangyang2,WU Li3
(1. School of Resources and Environment, Anhui Agricultural University, Hefei 230036; 2. Anhui Province Key Laboratory of Aquaculture & Stock Enhancement, Fisheries Research Institute, Anhui Academy of Agricultural Sciences, Hefei 230001; 3. School of Life Sciences, Hefei Normal University, Hefei 230061)
In order to study the temporal and spatial variation characteristics of crustacean zooplankton functional groups and their main influencing factors in Chaohu Lake, 20 sampling points were set up in the main estuary and pelagic areas of Chaohu Lake to investigate and analyze the community structure of crustacean zooplankton and water environmental factors. The results showed that: (1) A total of 21 species of 10 genera of cladocera and 16 species of 11 genera of copepods were identified in 8 sampling sites in Chaohu estuary; 18 species of 9 genera of cladocerans and 13 species of 12 genera of copepods were identified in 12 sites of pelagic areas; a total of 22 genera and 39 species. (2)The average density of crustaceans in estuary and pelagic of Chaohu Lake was 530.8 ind·L-1and 506.3 ind·L-1, and the average biomass was 14.7 ind·L-1and 21.5 ind·L-1, respectively; the Shannon-Wiener diversity index of crustacean zooplankton showed that the pollution degree in the estuary area of Chaohu Lake in autumn was higher than that in the pelagic area. (3)The crustaceans in Chaohu Lake could be divided into 5 functional groups according to their body size and feeding habit. Among them, there were 2 dominant functional groups of SCF and LCF in estuary, and 3 dominant functional groups of SCF, LCF and MCF in pelagic area. (4)Water temperature was the main environmental physical and chemical factor affecting the functional groups of planktonic crustaceans in estuary and pelagic area; the LCF functional group of planktonic crustaceans in the pelagic area of Chaohu Lake can significantly affect SCF, MCC,LCC and other functional groups. The pollution degree in the estuary area of Chaohu Lake was higher than that in the pelagic area. The eutrophication control and algae bloom control of Chaohu Lake should focus on the exogenous pollution control in the estuary area. Pelagic area can reconstruct the aquatic ecosystem by adjusting the structure of planktonic crustacean functional groups.
Chaohu Lake; estuary area and pelagic area; functional group of planktonic crustaceans; environmental factors; RDA analysis
10.13610/j.cnki.1672-352x.20230625.015
2023-06-26 15:55:41
S917.4
A
1672-352X (2023)03-0463-10
2022-07-21
安徽省重点研究与开发计划生态环境专项(202004i07020007)、安徽省水产增养殖安徽省重点实验室开放课题(AHSC20190202)、安徽省现代农业产业技术体系建设专项(皖农科函〔2021〕711号)共同资助。
张 佳,硕士研究生。E-mail:zzhj0304@126.com
通信作者:郭匿春,博士,副教授。E-mail:guonichun@ahau.edu.cn
[URL] https://kns.cnki.net/kcms2/detail/34.1162.S.20230625.1520.030.html
猜你喜欢
中国水运(2022年4期)2022-04-27
潍坊学院学报(2020年2期)2021-01-18
儿童与健康(幼儿教师参考)(2020年9期)2020-09-29
流行色(2019年10期)2019-12-06
城市建筑空间(2018年12期)2018-08-26
现代园艺(2017年23期)2018-01-18
——记山东省东营市河口区河安小学校长刘波
教书育人(2017年23期)2017-09-26
环境科技(2016年2期)2016-11-08
应用海洋学学报(2015年3期)2015-11-22
应用海洋学学报(2014年3期)2014-11-22
