
内蒙古绒山羊毛长的遗传规律研究
2023-08-02 03:33李学武王瑞军李晓凯刘志红李金泉
中国饲料 2023年14期
李学武,王瑞军,李晓凯,张 弋,兰 玛,苏 蕊,刘志红,李金泉
(1.攀枝花学院 康养学院,四川攀枝花 617000;2.内蒙古农业大学 动物科学学院,内蒙古呼和浩特 010018)
动物毛纤维主要有两种类型,分别是单层毛被类型和双层毛被类型,单层毛被类型其毛囊所产的毛纤维均为细毛,如安哥拉山羊、美利奴羊,双层毛纤维类型是由生产粗毛的初级毛囊和生产细毛的次级毛囊组成,如绒山羊、细毛羊(Sumner和Bigham,1993)。内蒙古绒山羊属于双层毛被类型,其毛纤维主要是由初级毛囊产生的羊毛和次级毛囊产生的羊绒组成,羊绒生长具有明显的季节性,但羊毛没有明显的季节性生长和脱落(Paula 等,1996),因此,按其毛长将内蒙古绒山羊分为3 个类型,即短毛型(≤13 cm)、中间型(>13 cm 且≤22 cm)和长毛型(>22 cm),并且经过显著性检验得出每个类型的毛长对其他抓绒性状均存在显著影响(李学武,2019)。
Mcgregor 等(2009)对安哥拉山羊的毛长进行REML(restricted maximum likelihood)分析,发现髋部、背中部两个位点可以作为群体毛长遗传选择位点(Mcgregor 和Butler,2009)。由于羊绒的经济价值高于羊毛,目前的研究中主要集中在羊绒的研究(Shamsalddini 等,2016),关于绒山羊羊毛的研究甚少,因此,本文对内蒙古绒山羊不同毛被类型进行分析,揭示绒山羊羊毛的遗传规律,为绒山羊的遗传育种奠定一定的理论基础。
1 材料与方法
1.1 数据来源 本试验所用数据来源于内蒙古白绒山羊种羊场,该羊场采用分群饲养管理,利用人工授精方法实施统一配种,生产性能数据和系谱数据记录完整。本研究收集了该羊场2001—2010 年间37086 只绒山羊羊毛的数据记录进行遗传参数估计。
1.2 统计分析
1.2.1 毛长的基本统计分析 将1990 ~2014 年毛长的重复记录数据利用Excel 进行初步整理并用于统计分析。ASREML 是由A.R.Gilmour 开发适用于大型数据混合线性模型的拟合分析软件,以REML 算法为主,结合平均信息算法(AI)和稀疏矩阵(Sparse matrix)可以快速准确地完成大型复杂模型的分析(Gilmour 等,2009)。
1.2.2 固定效应测定 根据Aylan-Parker 和McGregor(2002)、Li 等(2008)研究得出了影响各抓绒性状的非遗传因素,通过SAS9.0 的GLM程序确定不同毛别类型各性状的固定效应,本文非遗传因素包括场年季(1990—2014)、个体年龄(1 ~7)、母羊年龄(2 ~8)、出生类型(单羔/ 双羔)和性别(公/ 母),而且在随机效应中除个体加性效应和个体永久环境效应外,母体加性效应和母体永久环境效应仅对早期生长性状有影响,对其他抓绒性状和绒毛品质性状没有影响。因此,综合固定效应和随机效应得出如下模型:
式中:y 是性状观察值,b 是固定效应,a 是个体加性效应,p 是永久环境效应。X、Z、W 分别是固定效应、个体加性效应、永远环境效应对应的结构矩阵,e 是随机残差效应。
1.2.3 遗传评估 通过选择全同胞家系模型(亲本模型)、亲本模型计算父系半同胞遗传力估计模型和母系半同胞遗传力估计模型研究亲本毛长对后代毛长的影响。全同胞家系模型(亲本模型):
y 代表性状观察值,β 代表固定效应,b 代表群体随机效应,m 代表父本随机效应,f 代表母本随机效应,mf 公母畜交互效应,e 代表随机残差效应。Z1、Z2、Z3和Z4分别是固定效应,群体随机效应,父本随机效应,母本随机效应,公母畜交互效应的结构矩阵,使用亲本模型计算全同胞遗传力公式:
使用亲本模型计算父系半同胞遗传力公式:
使用亲本模型计算母系半同胞遗传力公式:
2 结果与分析
2.1 毛长的基本统计分析结果 由图1 可知,内蒙古绒山羊的毛长分布符合正态分布,分位数也在一条直线上,所以数据科学合理规范。
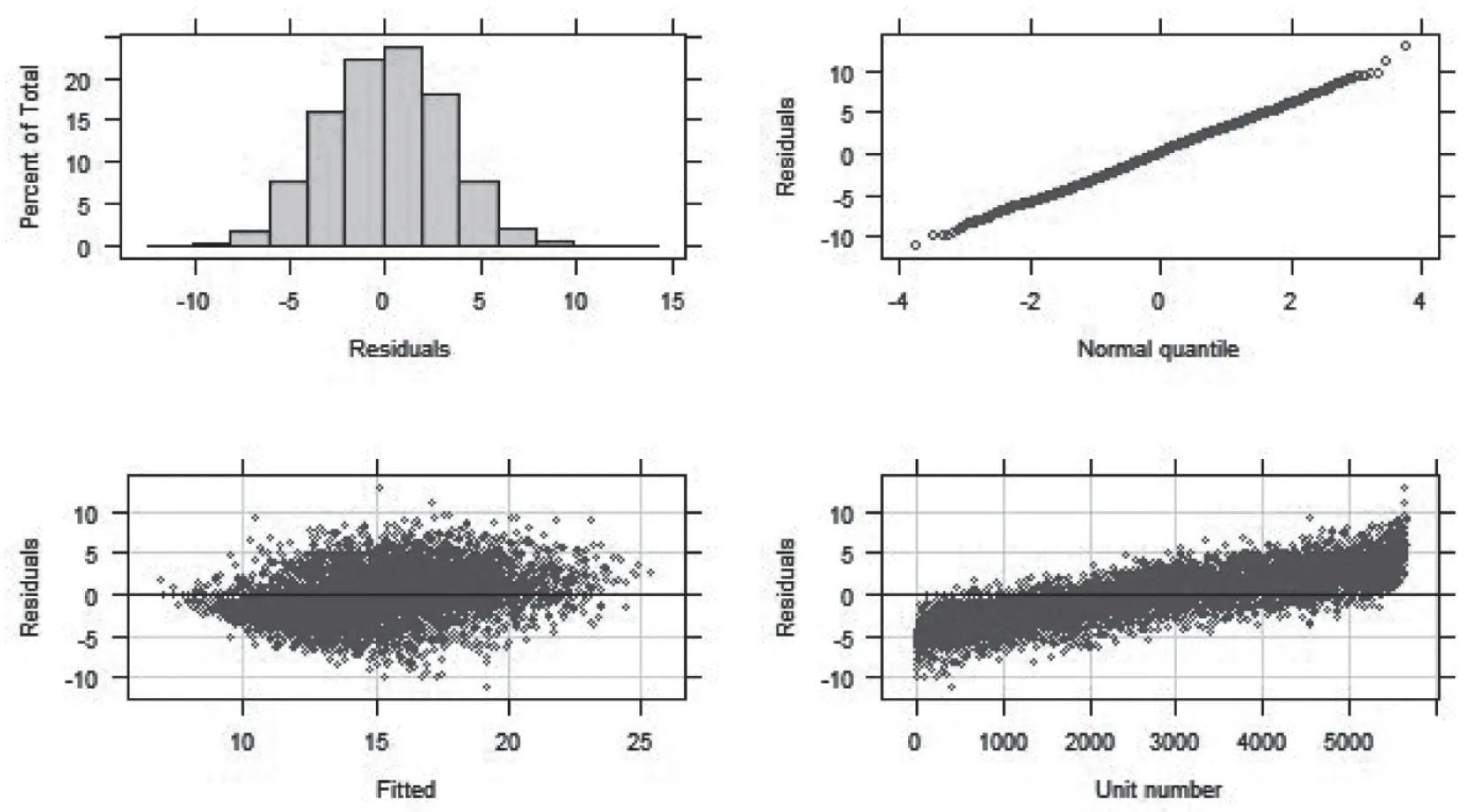
图1 内蒙古绒山羊羊毛表型分布图
2.2 固定效应 由表1 可知,测定年份、群、个体年龄、母羊年龄对毛长影响极显著(P<0.01),性别对内蒙古绒山羊的毛长影响不显著(P>0.05)。

表1 毛长的固定效应
2.3 遗传参数评估 由表2 可知,毛长的全同胞遗传力0.56、父系半同胞遗传力为0.26、母系半同胞遗传力为0.86。可以看出母系半同胞的遗传力显著高于父系遗传力(P<0.05)。
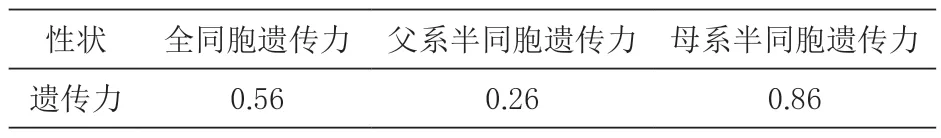
表2 遗传力评估结果
3 讨论
3.1 毛长表型规律分析 经过对内蒙古绒山羊羊毛长度表型统计分析发现,内蒙古绒毛羊毛长度为5 ~34 cm,内蒙古绒山羊羊毛长度分布基本符合正态分布,并且李学武等(2018)研究不同毛被类型发现,内蒙古绒山羊羊毛长度对内蒙古绒山羊其他重要经济性状具有重要影响。由此可以推测,控制内蒙古绒山羊羊毛长度的基因与控制其他重要经济性状的基因存在一因多效或者连锁遗传。因此,通过对不同毛被类型的选择可以对其他重要经济性状进行间接选择,从而提高内蒙古绒山羊的选育效率,缩短世代间隔,加速内蒙古绒山羊重要经济性状的遗传进展。
3.2 固定效应 结果表明,内蒙古绒山羊毛长的固定效应有所差别,测定年份、群、个体年龄、母羊年龄对毛长影响极显著,性别对内蒙古绒山羊的毛长影响不显著。由于内蒙古绒山羊的养殖方式为半舍饲半放牧,所以绒山羊的生产性能受环境影响较大。同样,Tao 等(2013)经过研究发现,生产性能测定年份对绒山羊的产绒量、断乳重、绒细,及绒长均有极显著的影响(P<0.01)。同样,Nugroho 等(2018)研究发现,测定年份对羔羊的断奶重具有重要影响。
3.3 遗传参数力评估结果 利用ASREML 软件计算全同胞遗传力、父系半同胞遗传力和母系半同胞遗传力分别为0.56、0.26 和0.86。由此可以发现,父系半同胞的遗传力最低,全同胞遗传力次之,母系半同胞的遗传力最高,由此猜测,控制毛长的主要遗传效应可能来自母本,李学武等(2018)研究发现,内蒙古绒山羊长毛型母羊后代的平均毛长高于长毛型公羊的毛长,而且长毛型内蒙古绒山羊的遗传力高于短毛型,说明毛长的选择不仅对种公羊的选择,母羊的选择同样重要,目前大部分母系遗传的研究主要集中在遗传资源和群体分化方面(Magotra 等,2021),关于性状的研究相对较少。本文也经过研究发现,母系半同胞的遗传力属于高遗传力,做好种母羊的选择可以加速毛长的遗传进展,提高选育效果和经济价值,由于母系半同胞的遗传力高,有可能毛长遗传是母系遗传。研究表明,毛长与绒长的遗传相关属于高度正相关,通过对内蒙古绒山羊毛长的选择也可以加速内蒙古绒山羊绒长的遗传进展,对其他重要经济性状的选择具有重要价值(李学武,2019)。
4 结论
本文通过对毛长的研究发现,毛长性状母系半同胞的遗传力最高,父系半同胞的遗传力最低,通过对母羊毛长的选择可以加速毛长的遗传进展,提高毛长的生产性能。
猜你喜欢
今日农业(2022年14期)2022-09-15
养猪(2022年4期)2022-08-17
中国听力语言康复科学杂志(2021年6期)2021-12-21
猪业科学(2018年5期)2018-07-17
新农业(2016年23期)2016-08-16
新农业(2016年22期)2016-08-16
中国组织化学与细胞化学杂志(2016年4期)2016-02-27
湖北畜牧兽医(2015年11期)2016-01-11
西域历史语言研究集刊(2015年0期)2015-08-21
山东农业科学(2014年1期)2015-03-09
