
鸭源鸡杆菌的药物敏感性及其对氟喹诺酮类药物的耐药机制
2023-09-25 11:34皇甫和平贾含笑孙彦婷薛通通彭志锋王宏魁曹素芳
中国兽医学报 2023年8期
皇甫和平,贾含笑,2,孙彦婷,薛通通,彭志锋,王宏魁,董 青,曹素芳*
(1.河南牧业经济学院,河南 郑州 450046;2.华中农业大学 动物医学院,湖北 武汉 430070)
鸭源鸡杆菌(Gallibacteriumanatis,G.anatis)是一种存在于鸡上呼吸道和下生殖道的条件致病菌,与产蛋鸡输卵管炎和腹膜炎关系密切,可造成产蛋量下降和感染禽的高致死率,被认为是引起家禽呼吸道和生殖道病变的主要原因之一[1-2]。近年来,关于G.anatis多重耐药现象的报道逐渐增多[3-6],同时由于该菌具有丰富的抗原多样性[1,7-8],从而使得采用药物和疫苗防治G.anatis感染效果不佳。现阶段,采取严格的生物安全措施,对该菌进行耐药监测,了解其药物敏感性和耐药机制,指导临床合理用药,仍然是控制这一疾病的可靠办法。
为了解近年来国内G.anatis的药物敏感性,本研究以实验室保存的204株G.anatis为对象,检测该菌的耐药情况。鉴于研究中发现这些G.anatis有高水平的氟喹诺酮类耐药性,有必要开展相应耐药机制的研究。氟喹诺酮类药物是人兽共用抗菌药物,近些年被广泛应用于畜牧生产。因此,细菌进化出了多种耐药机制来适应这种药物选择压力,相关研究主要集中在2个方面:一是染色体基因组上的DNA旋转酶和拓扑异构酶Ⅳ基因的喹诺酮耐药决定区(quinolone resistance-determining region,QRDR)突变导致耐药[9-11];二是质粒介导的喹诺酮耐药[12-15]。QRDR突变是细菌对氟喹诺酮类药物产生耐药的最主要原因[9-10],因此本试验拟通过检测实验室保存的G.anatis中QRDR存在的突变位点,分析不同突变位点在氟喹诺酮类耐药中的作用,以揭示G.anatis分离株对氟喹诺酮类药物高度耐药的分子机制。
1 材料与方法
1.1 仪器与试剂ETC-811 PCR仪购自北京东胜创新生物科技有限公司;3-30K高速冷冻离心机购自德国Sigma公司;DW-HL398 -80℃超低温冰箱购自中科美菱低温科技股份有限公司。4S Green核酸染料、DNA Marker购自生工生物工程(上海)股份有限公司;绵羊血琼脂平板购自郑州福博赛生物技术有限责任公司;药敏纸片氨苄西林(AMP)、环丙沙星(CIP)、庆大霉素(CN)、妥布霉素(TOB)、四环素(TE)、左氧氟沙星(LEV)、头孢曲松(CRO)、阿米卡星(AK)、头孢噻肟(CTX)、复方新诺明(SXT)和氧氟沙星(OFX)购自英国Oxoid Limited公司;麦氏比浊管购自北京天安联合科技有限公司;DNA提取试剂盒、PCR扩增试剂购自宝生物工程(大连)有限公司。
1.2 菌株204株G.anatis由本实验室保存,其中包括从河南、山西、山东、湖北、陕西和上海市等地的养禽场或动物医院分离的192株,以及由河南农业大学传染病实验室惠赠的12株。基本菌株信息见表1。
1.3 引物参照文献[16]的方法,委托生工生物工程(上海)股份有限公司合成gyrA、parC基因PCR引物,引物序列见表2。
1.4 药物敏感性试验根据美国临床和实验室标准协会(CLSI)抗菌药物敏感性试验标准(M45),按照常规K-B纸片法,采用11种药物对204株G.anatis进行药物敏感性试验。
1.5gyrA、parC基因的扩增及测序取细菌LB肉汤培养物1 mL加入1.5 mL离心管中,10 000 r/min离心5 min,弃上清,加入600 μL的TE 缓冲液吹打混匀,100℃沸水浴10 min,立即冰浴2 min,12 000 r/min离心10 min,取上清立即使用,或-20℃ 保存备用。
选取其中30株G.anatis进行gyrA、parC基因的PCR扩增。反应体系(25 μL):TaKaRa Ex Taq 0.125 μL,dNTP 2.0 μL,Ex Taq buffer 2.5 μL,上、下游引物各1 μL,模板DNA 1 μL,灭菌超纯水17.375 μL。反应条件:95℃预变性5 min;94℃ 1 min,52℃ 50 s,74℃ 50 s,30个循环;72℃ 10 min。PCR产物由生工生物工程(上海)股份有限公司进行测序,测序成功后上传至GenBank数据库。

表1 G.anatis菌株基本信息

表2 gyrA、parC基因引物
1.6gyrA、parC基因QRDR编码氨基酸突变分析从30株试验菌株中选取对环丙沙星耐药和敏感的G.anatis菌株,与GenBank收录的G.anatis、E.coli一并进行分析。使用DNAStar、Clustalx 1.81软件,分别对上述菌株的gyrA、parC基因序列进行比对,分析其QRDR编码氨基酸的突变情况。30株G.anatis和其他用于比较菌株的信息见表3。
2 结果
2.1 204株G.anatis的药物敏感性204株G.anatis对11种药物的敏感性结果见表4和图1。结果显示,11种药物中有9种的耐药率达40%以上,其中环丙沙星、左氧氟沙星、氧氟沙星、复方新诺明、四环素和氨苄西林的耐药率达84.80%以上;庆大霉素、阿米卡星、头孢噻肟和头孢曲松的敏感率较高,分别为57.35%,48.53%,39.22%,37.25%。
多重耐药情况见表5和图2。结果显示,耐3种以上药物的菌株有202株,占比99.02%;耐6种以上药物的菌株有188株,占比92.16%,其中有6株对所选的11种药物全部耐药。
2.2 30株G.anatisgyrA和parC基因的PCR扩增和测序通过gyrA基因PCR扩增,30株G.anatis中有22株扩增出约433 bp预期DNA片段,部分菌株的电泳结果见图3;通过parC基因PCR扩增,30株G.anatis均扩增出约418 bp预期DNA片段,部分菌株的电泳结果见图4。PCR产物送至生工生物工程(上海)股份有限公司进行DNA测序,测序结果上传至GenBank,登录号见表6。

表3 用于氟喹诺酮类药物耐药基因分析的有关菌株信息
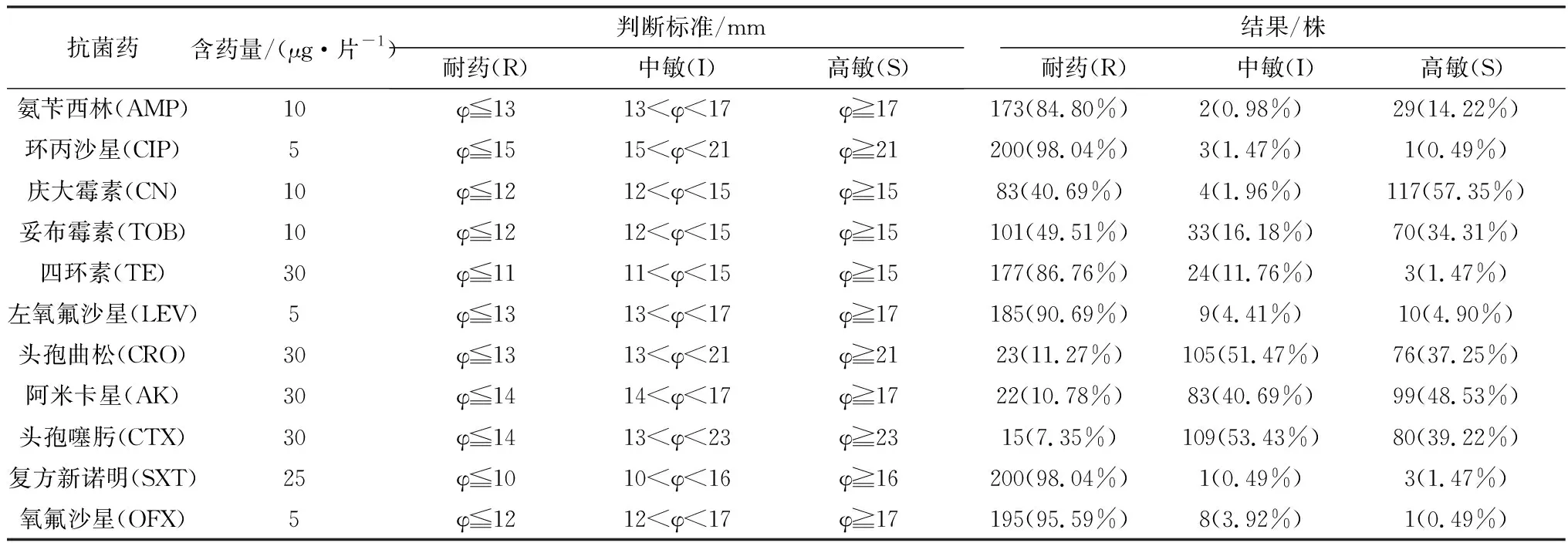
表4 药物敏感性试验结果

图1 204株G.anatis对11种药物的耐药比例
2.3gyrA、parC基因QRDR序列比对结果表6中F149T为环丙沙星敏感菌株,其余实验室收集的G.anatis为环丙沙星耐药菌株。将同时检测出gyrA和parC基因的22株菌,与表6中GenBank收录的其他G.anatis和E.coli参考菌株,进行GyrA和ParC亚基氨基酸序列对比发现,G.anatis耐药菌株GyrA亚基存在Ser83→Phe、Asp87→Ala/Tyr/Asn和Asp139→Tyr位点氨基酸的改变(图5)。G.anatis耐药菌株ParC亚基存在Thr84→Ile、Glu88→Gly、Ala94→Val和Gly179→Val位点氨基酸突变(图6)。

表5 204株G.anatis的多重耐药情况

图2 G.anatis多重耐药柱状图

M.DL2000 DNA Marker;1~22.G. anatis 菌株;23.阳性对照;24.空白对照

M.DL2000 DNA Marker;1.空白对照;2.阳性对照;3~24.G.anatis 菌株

图5 GyrA亚基的QRDR序列比对

图6 ParC亚基的QRDR序列比对
2.4G.anatis对环丙沙星的敏感性与其QRDR突变的关系以同时检出gyrA和parC基因的22株G.anatis为对象,分析其对环丙沙星的敏感性与QRDR突变的关系,上述菌株对环丙沙星的耐药表型与QRDR突变情况见表6。由表6可知,对环丙沙星敏感的菌株F149T没有发生氨基酸的替换;中度敏感菌株GAC003、GAC006发生了3处氨基酸的替换;19株耐药菌株中,除1株存在3处氨基酸替换外,其余18株菌存在4~5处氨基酸的替换;环丙沙星耐药菌株均发生了GyrA亚基83,87位和ParC亚基84位氨基酸的替换。

表6 22株G.anatis QRDR的突变分析
3 讨论
药感试验结果显示,试验菌株对氟喹诺酮类药物、复方新诺明、四环素和氨苄西林的耐药率达到84.80%以上,其中对环丙沙星、氧氟沙星和复方新诺明耐药的菌株占比甚至达到了95.59%以上,只有头孢曲松与头孢噻肟的耐药率较低,分别为11.27%和7.35%,与研究报道[5,17-18]的结果相似,反映出G.anatis分离株中耐药性极为普遍。204株G.anatis中,99.02%的菌株耐受3种以上药物,92.16%的菌株耐受6种及以上药物,甚至有6株菌对11种药物全部产生了耐药性,该结果进一步验证了VAN DRIESSCHE等[4]、唐诗等[8]、彭志锋等[19]的研究结果,说明该菌作为一种条件致病菌,目前存在严重的多重耐药性。上述研究结果一方面为防治G.anatis感染提供了一定的参考;另一方面也提醒人们要高度重视该菌日益严重的耐药问题,积极探索绿色健康养殖模式,尽量减少养殖过程中抗菌药物的应用。
G.anatis在国内外的家禽中广泛存在[20-22],2011年BOJESEN等[23]报道墨西哥和丹麦两个国家23个鸡群中分离的46株G.anatis对环丙沙星耐药率高达100%;2018年ELBESTAWY等[24]对埃及的102个鸡群进行了调查,发现从2013-2015年分离的菌株对诺氟沙星和环丙沙星的耐药性有所增强。国内G.anatis分离株对氟喹诺酮类药物的耐药性发展也很快,2016年彭志锋[19]报道了61株G.anatis的药敏试验,结果显示分离株对环丙沙星的耐药率达到83.61%。本研究中,G.anatis对环丙沙星、氧氟沙星和左氧氟沙星的耐药率分别达到98.04%,95.59%,90.69%,上述研究说明国内G.anatis对氟喹诺酮类药物的耐药性有愈发严重之势,亟需开展相应的耐药机制研究。
本研究通过对22株G.anatis的耐药表型与基因型的分析发现,耐药菌株的GyrA亚基上检测到3个氨基酸位点(Ser83、Asp87和Asp139)的突变,ParC亚基上检测到4个氨基酸位点(Thr84、Glu88、Ala94和Gly179)的突变,所有的耐药菌株均发生了GyrA亚基83,87位和ParC亚基80位氨基酸的突变,推测这3个位点突变对其氟喹诺酮类药物耐药性的产生十分重要,很可能起到前提和基础的作用,而GyrA亚基的139位和ParC亚基88,94,179位氨基酸的突变只存在于部分菌株,非耐药所必需的,推测有增强耐药程度的作用。敏感菌株F149T不存在这些位点的突变,中度敏感菌株发生了3处氨基酸的替换,19株耐药菌有1株发生3处氨基酸位点替换,其余18株(94.7%)均发生4处及以上氨基酸位点的替换,说明QRDR编码氨基酸发生突变的数量与耐药程度呈正相关。
G.anatis中QRDR突变导致其对氟喹诺酮类药物耐药的机制与其他细菌类似[10,25-26],但也存在一些差别,在1株耐药菌株的GyrA亚基上同时存在Asp139→Tyr位点氨基酸的置换,在5株耐药菌株的ParC亚基上同时存在Gly179→Val位点氨基酸的改变,从而导致相应菌株在2个亚基上氨基酸置换达到5处,表明G.anatis分离株QRDR突变正在变得更为复杂;G.anatisGyrA亚基发生氨基酸替换的位点(83,87)与大肠杆菌、沙门菌一致,而G.anatisParC亚基发生替换的位点(84,88,94)与上述其他菌株不一致,对应于这些菌株的80,84,90位点[27-29],提醒进行G.anatisQRDR突变分析时应该注意。
G.anatis分离株产生了严重的多重耐药性,对氟喹诺酮类、四环素和磺胺类等药物高度耐药。该菌对氟喹诺酮类药物的高度耐药性与QRDR编码氨基酸位点突变关系密切,突变位点数量与耐药情况呈正相关;在其GyrA、ParC亚基QRDR区域存在7个突变位点,推测GyrA83、GyrA87和ParC84位氨基酸的突变对氟喹诺酮类药物的耐药起着决定性作用,其余4个位点突变起协同增强作用。
猜你喜欢
湘潭大学自然科学学报(2022年2期)2022-07-28
世界最新医学信息文摘(2021年12期)2021-06-09
世界最新医学信息文摘(2020年68期)2020-12-25
现代检验医学杂志(2016年2期)2016-11-14
农业环境科学学报(2016年10期)2016-11-11
西安理工大学学报(2016年3期)2016-11-10
国外医药(抗生素分册)(2016年5期)2016-07-12
国外医药(抗生素分册)(2016年2期)2016-07-12
中国卫生标准管理(2015年25期)2016-01-14
吉林大学学报(医学版)(2015年4期)2015-12-17
