
基于实验种群两性生命表的伊氏叶螨在番茄和龙葵上的适应性评价
2023-09-25 11:54陈凌娟范青海尤民生蔡立君
生物安全学报 2023年3期
钟 隽 , 陈凌娟, 高 菲 , 范青海 , 尤民生* , 蔡立君*
1福建农林大学闽台作物有害生物生态防控国家重点实验室,福建 福州 350002; 2福建农林大学应用生态研究所,福建 福州 350002; 3福建农林大学教育部害虫生态防控国际合作联合实验室,福建 福州 350002; 4福建农林大学海峡两岸特色作物安全生产省部共建协同创新中心,福建 福州 350002; 5新西兰初级产业部植物健康与环境实验室,新西兰 奥克兰 1142
伊氏叶螨TetranychusevansiBaker and Pritchard隶属于蜱螨亚纲Acari叶螨科Tetranychidae,俗称番茄红叶螨(tomato red spider mite)(Navajasetal.,2013)。该螨寄主范围广,涉及37科138种植物,偏好取食茄科Solanaceae植物,如番茄SolanumlycopersicumL.、茄子S.melongenaL.、马铃薯S.tuberosumL.、龙葵S.nigrumL.以及刺茄S.touvumSwartz等(Migeon &Dorkeld,2010; Silva,1954),也危害豆类、棉花、蓖麻、柑橘类作物以及一些绿化植物(Fanetal.,2021)。伊氏叶螨取食危害会造成植物叶片黄化、枯萎,果实退色、脱落,严重时整株死亡(Wakiletal.,2018)。该螨曾严重危害非洲东南部与西部茄科作物,导致减产90% (Azandémè-Hounmalonetal.,2015; Sibandaetal.,2000)。
伊氏叶螨最早在1952年发现于巴西,1960年方命名(Baker &Pritchard,1960; Silva,1954),之后逐步入侵北美、非洲、欧洲等地区,现已广泛分布于全球45个国家和地区(Boubouetal.,2012; Danieletal.,2010; Guaniloetal.,2010; Meynardetal.,2013)。2005年首次在日本和我国台湾地区发现,目前在我国广东、广西和四川的茄子、番茄和黄瓜CucumissativusL.上也发现该害螨,表明该螨至少入侵了中国以上3省。笔者于2019—2021年在福建、江西的番茄与龙葵上采集到伊氏叶螨(玻片标本经由叶螨分类学专家进行形态学鉴定并保存于福建农林大学)。
伊氏叶螨具有耐高温、种群增长速度快、世代时间短等特征,表现出极强的入侵性(Ghazyetal.,2019)。如伊氏叶螨入侵瓦伦西亚后导致当地本土叶螨种群数量大幅度下降(Ferragutetal.,2012)。当伊氏叶螨与二斑叶螨T.urticaeKoch同时危害番茄时,前者因具有更大的环境最大容纳量与更快的种群增长速度等特点在种间竞争中更占优势(Djossouetal.,2020; Savietal.,2019a,2019b)。
年龄-阶段两性生命表是研究生物种群生长发育与繁殖的常用方法。两性生命表中用于评价昆虫种群特征的常用参数包括发育历期、性比(Nf/N、Nm/N)、繁殖力F、存活率sxj、内禀增长率r、周限增长率λ、净生殖率R0、平均世代周期T等(Chi &Liu,1985),在一定程度上反映昆虫种群在存活率、龄期分化、发育历期、繁殖等方面的情况,综合评价了害虫的危害潜力(齐心等,2019; Yang &Chi,2006)。目前,两性生命表已被广泛应用于害虫治理、杀虫剂抗性研究、天敌-害虫相互关系及捕食研究等方面(Chietal.,2020)。Zrikietal.(2013)通过对比伊氏叶螨在叙利亚的番茄和龙葵上的生命表,发现伊氏叶螨入侵叙利亚后对其寄主植物番茄和龙葵都表现出高适应性。Murungi (2010)对比伊氏叶螨取食5种非洲龙葵属植物(毛龙葵S.sarrachoides、少花龙葵S.americanum、红果龙葵S.villosum、S.tarderemotum、木龙葵S.scabrum)的生命表参数,发现其在取食毛龙葵后存活率低,雌成螨寿命短,产卵量少,且种群动态参数(r、λ、R0、T)弱于其余4种茄科寄主植物,表现出极低的适应性。
番茄在我国的种植面积约125万 hm2,年产量5000 t,品种10000余种,是我国蔬菜产业的重要组成部分(赵洪等,2020)。目前伊氏叶螨在我国番茄上的危害尚缺乏研究。龙葵常见于路边、荒地、山坡等地,广泛分布于我国各地(程海涛等,2014),但作为伊氏叶螨的野外寄主之一,其对伊氏叶螨在我国的传播潜力尚无报道。本研究选用我国2种常见茄科植物番茄和龙葵作为寄主植物,对比伊氏叶螨在这2种植物上的生长发育及繁殖参数,为防治国内伊氏叶螨种群提供理论基础。
1 材料与方法
1.1 供试虫源
伊氏叶螨由南京农业大学提供,在人工气候室内分别用番茄和龙葵植株饲养5代以上。
饲养小室由3层亚克力板构成,上层为长方形(3.5 cm×2.5 cm×0.17 cm),中、下2层为正方形(中层为3.5 cm×3.5 cm×0.27 cm,下层为3.5 cm×3.5 cm×0.17 cm)。中层板中心打一个直径为1.5 cm的圆孔。在中层和下层板之间夹入一片直径大于1.5 cm的寄主植物叶片,使圆孔位于叶片中间,将叶螨放寄主植物叶片上后盖上上层板,将三层板叠放后用燕尾夹固定板四角,置于浸满水的海绵垫上饲养、观察。
所有试验均在温度(25±2)℃、相对湿度(70±5)%、光暗比L∶D=16 h∶8 h实验室条件下进行。
1.2 供试植物
供试番茄品种粉冠和龙葵种子购买自山东寿光欣欣然园艺有限公司,经浸种后种于花盆中,待植株叶片直径大于1.5 cm时用于试验。
1.3 伊氏叶螨生命表试验方法
分别从番茄和龙葵上的伊氏叶螨种群中随机挑取100对雌雄成螨,来自同种寄主植物的两性进行配对,等待雌螨产卵。从2个种群中分别随机挑取100粒4 h内新产的卵,单头放入盛有对应寄主植物叶片的饲养小室内,每12 h观察、记录各螨态的存活数以及发育状态;待发育至成螨后,将雌雄成螨配对,挑入同一小室饲养,并记录雌雄成螨存活数与雌成螨产卵量,直至所有个体死亡。若雄成螨死亡,及时补入备用雄成螨;如雄螨和雌螨不够配对,从备用虫源中挑相应性别的成螨用以配对。试验过程中及时更换饲养小室及寄主植物叶片。
1.4 数据统计与分析
根据Chi &Liu (1985)的研究方法构建伊氏叶螨实验室种群的年龄-龄期两性生命表,利用年龄-龄期两性生命表软件TWOSEX-MSChart进行数据处理和分析(单位时间为0.5 d),得到特定年龄-龄期存活率Sxj(Sxj表示个体存活到年龄x、发育阶段j的概率,x为按龄期划分的单位间距,j为特定阶段);
其他相关参数的计算采用公式如下:

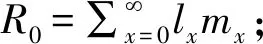

周限增长率λ=er。
采用Bootstrap方法计算种群动态参数的平均值和标准误,其中Bootstrap重复次数为100000次。采用paired bootstrap test法在TWOSEX-MSChart软件中进行差异显著性检验(Huangetal.,2017)。使用SigmaPlot 14.0软件绘图。
2 结果与分析
2.1 伊氏叶螨在番茄和龙葵上的发育与繁殖参数
取食番茄和龙葵的伊氏叶螨的发育历期与繁殖参数如表1所示,除后若螨期外其他未成熟期均存在显著差异,说明伊氏叶螨在未成熟期取食龙葵比取食番茄发育得更快。在2种寄主植物上伊氏叶螨雌雄成螨均可存活20 d以上,其中取食番茄的雄成螨存活时间最长,为38.95 d;取食番茄的雌成螨较取食龙葵的雌成螨存活时间长,为35.36 d。各发育历期中,取食番茄的伊氏叶螨的卵期、幼螨期和前若螨期比取食龙葵的伊氏叶螨显著延长。取食番茄的伊氏叶螨的总寿命与取食龙葵的伊氏叶螨总寿命间存在显著差异。取食番茄的成虫产卵前期(APOP=1.51)与取食龙葵的成虫产卵前期(APOP=1.43)间无显著差异。取食番茄的总产卵前期(TPOP=16.08)显著长于取食龙葵的总产卵前期(TPOP=13.80),表明取食龙葵的雌成螨比取食番茄的雌成螨更早进入产卵期。取食番茄和龙葵的雌成螨平均产卵量分别为61.02和65.19粒,无显著差异。
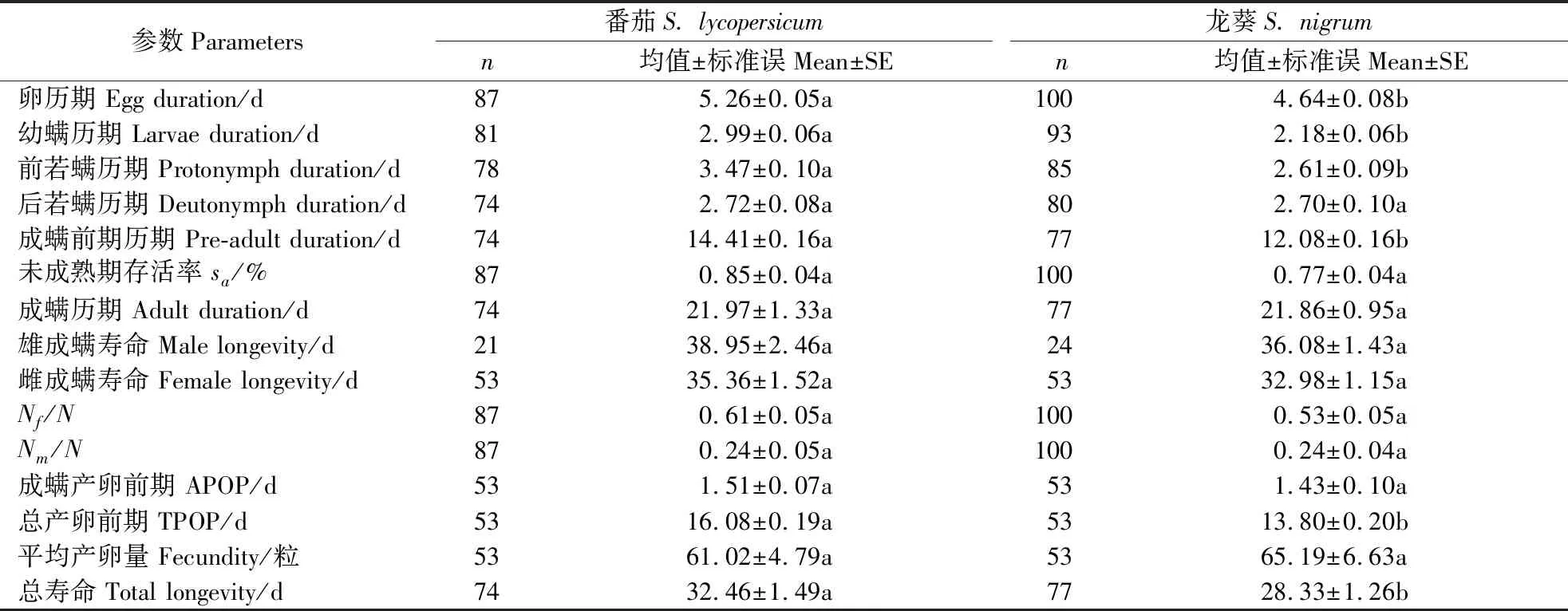
表1 以不同寄主植物为食的伊氏叶螨在各发育阶段历期与繁殖参数
2.2 伊氏叶螨在番茄和龙葵上的存活率和繁殖率
取食番茄和龙葵的伊氏叶螨的存活率曲线(图1)均发生重叠,成螨前期的存活率均高于其成螨,雌成螨存活率高于雄成螨。取食番茄的伊氏叶螨未成熟期存活率(Sa=0.85)与取食龙葵的(Sa=0.77)无显著差异(p=0.16)。成螨存活时间长于其余龄期,且取食番茄的伊氏叶螨成螨存活时间长于取食龙葵的。取食番茄时从卵发育至雌成螨的比率(Nf/N=0.61)和取食龙葵时(Nf/N=0.53)无显著差异;取食番茄的伊氏叶螨从卵发育至雄成螨的比率(Nm/N=0.24)与取食龙葵饲养的(Nm/N=0.24)亦无显著差异(p=0.99)。取食番茄的伊氏叶螨雌成螨出现在第12天,在第13.5天开始产卵;取食龙葵的雌成螨出现在第9天,在第11天开始产卵(图2)。
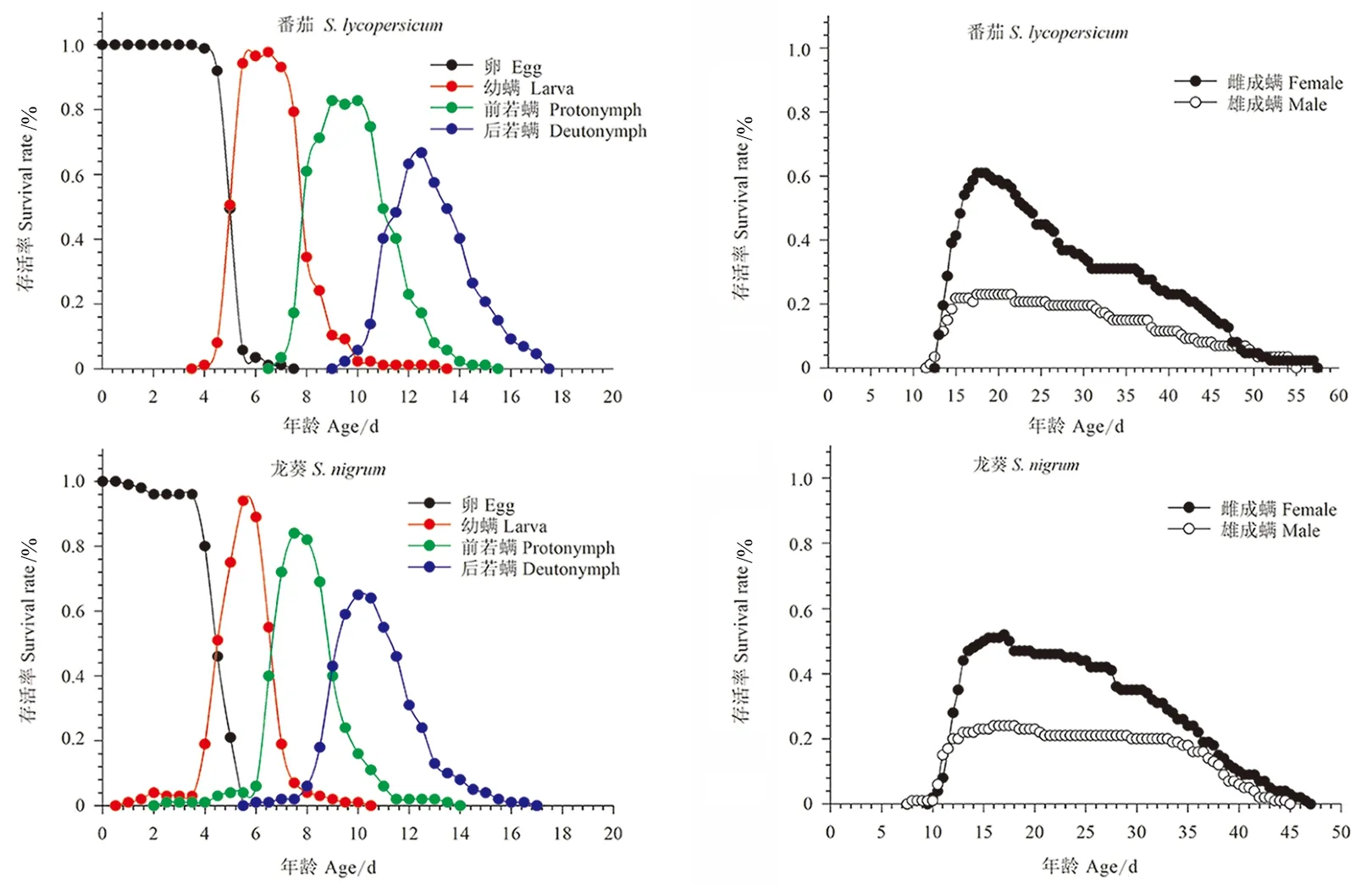
图1 取食番茄和龙葵的伊氏叶螨年龄-龄期存活曲线
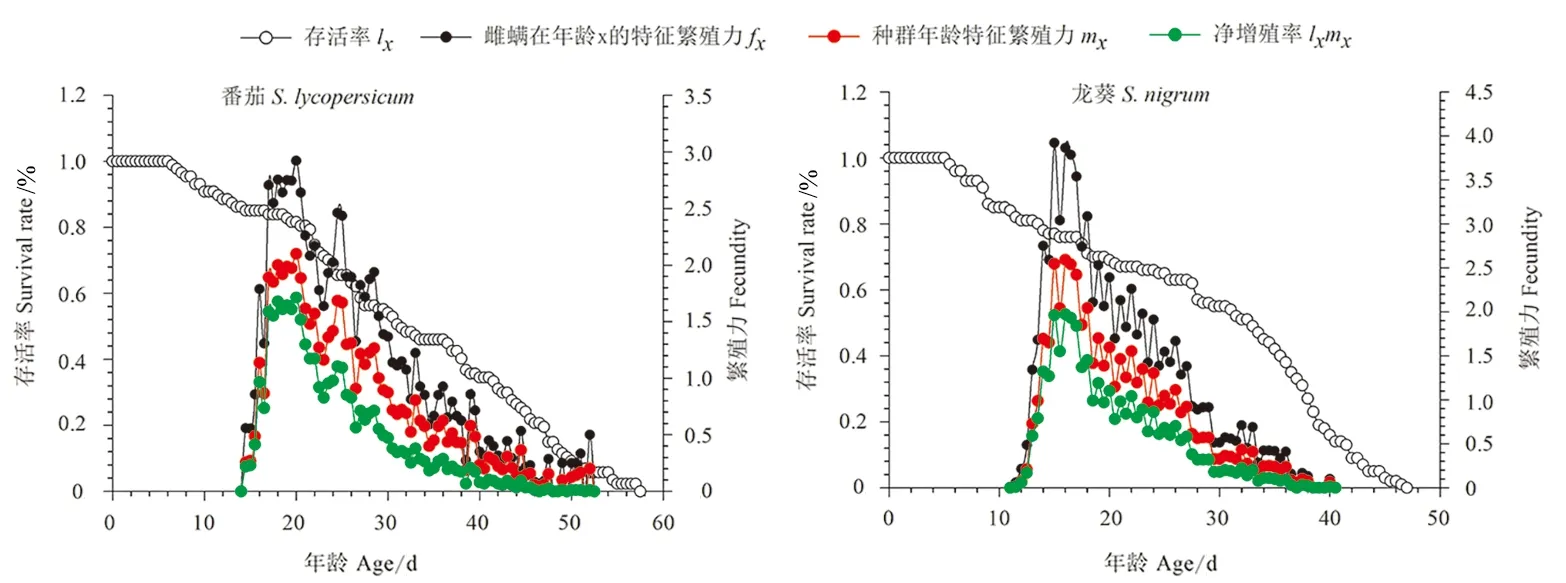
图2 取食番茄和龙葵的伊氏叶螨年龄存活率(lx)、雌螨在年龄x的年龄-龄期繁殖力(fx)、种群年龄特征繁殖力(mx)及净增殖率(lxmx)
取食番茄或龙葵的伊氏叶螨存活率随年龄增长而下降,下降趋势平缓。在0~20 d内,取食番茄的伊氏叶螨存活率大于取食龙葵的伊氏叶螨存活率。取食番茄和龙葵的伊氏叶螨雌成螨的繁殖力fx5和种群年龄-龄期特征繁殖力mx均呈现先上升后下降的趋势。取食番茄的伊氏叶螨的fx5和mx在第20天达到最大值,分别为2.92和2.10;取食龙葵的伊氏叶螨的fx5和mx分别在第15和16天达到最大值,分别为3.92和2.59。取食番茄的伊氏叶螨净增殖率于第21天达到最大值1.61,取食龙葵的伊氏叶螨净增殖率于第16天达到最大值1.97(图2)。
2.3 取食番茄和龙葵的伊氏叶螨期望寿命值
取食番茄和龙葵的伊氏叶螨期望寿命值总体趋势均随年龄增加而降低,生活史时间长度相似;取食龙葵的后若螨阶段出现先增高后降低再增高再降低的趋势。取食番茄的雄成螨最大期望寿命值为26.84 d,出现在第12天;取食龙葵的雄成螨最大期望寿命值出现在第8天,为28.08 d。取食番茄的雌成螨最大期望寿命值为22.36 d,出现在第13天;取食龙葵的雌成螨最大期望寿命值出现在第10天,为22.93 d(图3)。
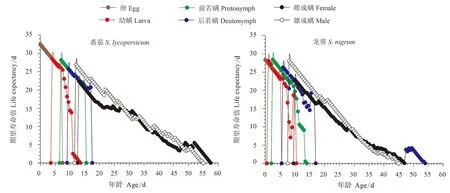
图3 取食番茄和龙葵的伊氏叶螨特定年龄-龄期期望寿命值(exj)
2.4 伊氏叶螨在番茄和龙葵上的繁殖价值
取食番茄和龙葵的伊氏叶螨繁殖价值总体趋势呈先增后减。番茄上伊氏叶螨卵的初产卵生殖力为1.09,龙葵上为1.10。取食番茄和龙葵的伊氏叶螨种群中对繁殖贡献最大的是雌成螨阶段。取食番茄的雌成螨繁殖峰值出现在第17天,达到24.68;取食龙葵的雌成螨繁殖峰值出现在第14天,为27.22。取食番茄的雌成螨繁殖峰值出现的时间较取食龙葵的伊氏叶螨晚,且峰值更小,说明取食番茄的伊氏叶螨种群较取食龙葵的伊氏叶螨种群增长速度慢(图4)。
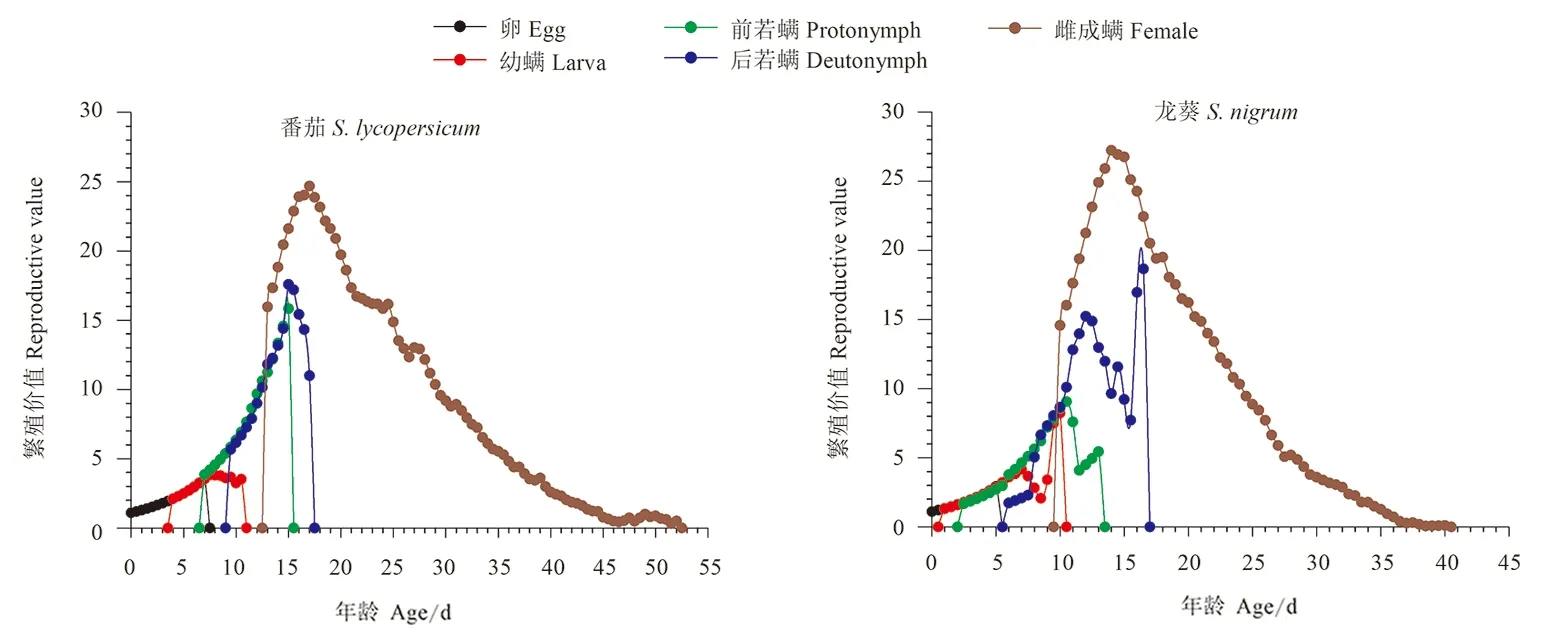
图4 取食番茄和龙葵的伊氏叶螨繁殖价值(vxj)
2.5 取食番茄和龙葵的伊氏叶螨稳定年龄-龄期分布
以番茄和龙葵为食的伊氏叶螨在稳定年龄-龄期分布上趋势相同,卵期稳定年龄结构呈随时间增长而下降趋势外,其他发育阶段都随时间增长呈先上升后下降的趋势,发育至成螨以后雌成螨所占比例均大于雄成螨(图5)。
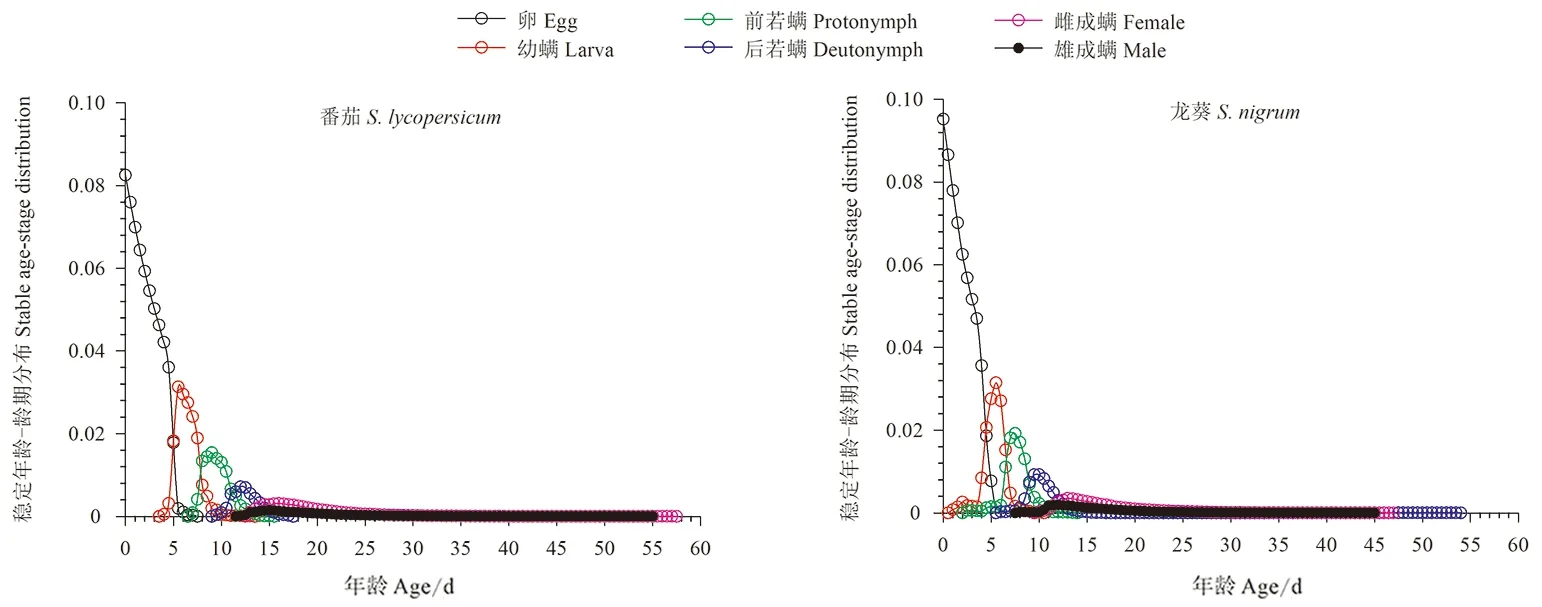
图5 取食番茄和龙葵的伊氏叶螨稳定年龄-龄期分布曲线
2.6 伊氏叶螨在番茄和龙葵上的种群动态参数
由表2可见,除净增值率(R0)外,其他种群动态参数均存在显著差异。取食龙葵的伊氏叶螨内禀增长率(r)、周限增长率(λ)显著高于取食番茄的,分别为0.19和1.21 d-1。取食龙葵的伊氏叶螨世代平均周期明显短于取食番茄的(p=0)。可见取食龙葵的伊氏叶螨种群比取食番茄的具有更快的增长速率及更短的世代时间,表现出更高的适应性。

表2 伊氏叶螨在番茄和龙葵上的种群参数
3 讨论与结论
作为世界性入侵害螨,伊氏叶螨因其极强的入侵能力早在21世纪初就被螨学家预测会入侵我国(Migeonetal.,2009)。本研究运用年龄-龄期两性生命表技术研究了取食番茄及龙葵的伊氏叶螨实验室种群生长发育与繁殖情况,结果显示,伊氏叶螨对番茄和龙葵表现出高适应性,说明伊氏叶螨入侵我国后可保持较高的种群活力,在顺利完成生长发育的同时亦可保持种群的稳定增长。与Zrikietal. (2013)的研究结果相比,本研究中的叶螨种群未成熟期和世代平均周期T更长,平均产卵量、内禀增长率r和净增值率R0更低。这一差异可能与研究中所用的寄主植物的品种不同有关。植食性昆虫的生长发育会受到寄主植物的种类及品种影响(Ode,2006),如苹果全爪螨PanonychusulmiKoch取食养分含量、含水量及叶片品质存在差异的4种桑树MorusalbaL.品种,其在产卵量、发育时间及存活率等方面表现出较大的差异(Dar,2016);叶螨T.bastosiTuttle取食菜豆PhaseolusvulgarisLinn.、木薯ManihotesculentaCrantz和木瓜Caricapapaya时,其发育时间及种群动态参数上表现出一定的差异(de Lima,2017)。另一个原因可能是因不同地理种群特征存在一定差异。Santamaría (2017)发现伊氏叶螨Nice地理种群在繁殖力、后代死亡率、卵存活率及成螨死亡率4个方面与Perpignan种群存在显著差异,前者表现出更高的繁殖力和存活率及更低的死亡率。
昆虫种群动态参数在一定程度上反映昆虫对不同寄主植物的适应性(刘孝纯和吴孔明,1988)。Zrikietal.(2013)发现,在叙利亚取食龙葵的伊氏叶螨未成熟期比取食番茄的短;在同种寄主为食的情况下,雄螨的未成熟期短于雌螨;雌成螨产卵前期无显著差异,但取食龙葵的平均产卵量、净增值率R0与内禀增长率r显著高于取食番茄的,说明寄主植物会影响叶螨的生长发育与繁殖,伊氏叶螨种群对龙葵的适应性高于番茄。本研究同样发现,伊氏叶螨取食番茄及龙葵后在卵期、幼螨期、前若螨期、内禀增长率r及世代平均周期T上差异显著,在龙葵上比番茄上有更高的适应性,可能说明伊氏叶螨在野外茄科植物龙葵上更容易定殖。Murungi (2010)发现,取食毛龙葵、少花龙葵的伊氏叶螨的未成熟期长于取食红果龙葵、S.tarderemotum、木龙葵的未成熟期;除毛龙葵外,取食其余4种龙葵后未成熟期存活率均达40.00%以上,而取食毛龙葵的未成熟期存活率仅为5.60%;取食毛龙葵后雌成螨平均产卵量极低,取食毛龙葵后伊氏叶螨的内禀增长率r为-0.063 d-1,种群呈负增长,表现出极低的适应性。本研究及以上试验植物均属于茄科植物,由于不同寄主植物间其叶片营养物质和化学物质存在一定的差异,不同茄科植物对伊氏叶螨种群的生长发育及繁殖的影响可能不一致。
本研究仅在实验室条件下进行,而环境因素如温湿度等均影响叶螨的种群变化(Ghazyetal.,2019),因此,还需关注其他非生物因素对伊氏叶螨种群生长的影响。本研究仅评估伊氏叶螨取食番茄和龙葵2种常见茄科寄主植物后其种群参数,取食其他茄科寄主植物或其他科寄主植物适应性尚不明确,因此无法预测取食其他寄主植物的种群增长情况,亟待进一步研究。
致谢:南京农业大学洪晓月教授提供试验虫源;台湾中兴大学齐心教授在两性生命表软件TWOSEX-MSChart的运用和理论知识上给予帮助,特此感谢。
猜你喜欢
植物保护(2023年4期)2023-08-05
中国诗歌(2018年6期)2018-11-14
运动(2017年1期)2017-09-03
江苏农业科学(2017年7期)2017-05-23
国学(2016年8期)2017-03-20
广西农学报(2015年4期)2015-09-11
环球人文地理(2015年11期)2015-09-10
食品工业科技(2014年7期)2014-03-11
湖南中医药大学学报(2011年11期)2011-03-20
