
喀斯特特有耐旱植物地枫皮抗氧化系统的水分响应特征研究
2023-10-08 03:13张秀姣王满莲梁惠凌刘宝玉
安徽农业科学 2023年18期
张秀姣,吴 超*,王满莲,梁惠凌,刘宝玉,李 虹,王 琳
(1.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006;2.广西植物功能物质与资源持续利用重点实验室,广西桂林 541006;3.广西师范大学,广西桂林 541006)
全球气候变化下干旱事件频发,导致土壤干旱程度加剧[1]。喀斯特岩溶地的保水能力极差,气候变化背景下极端干旱事件(土壤含水量<12%)时常发生[2]。前人关于作物适应干旱方面的研究以主要农作物和部分园艺植物为主,但极端干旱下大部分植物难以存活,由于缺乏适应极端干旱研究的优异物种和种质资源,有关作物响应极端干旱的研究较少,作物适应极端干旱的内在机理不够深入[3]。地枫皮(IlliciumdifengpiB.N.Chang et al.)能适应喀斯特山顶极度干旱环境,是研究植物响应和适应极度干旱的优异材料[4]。
地枫皮为五味子科(Schisandraceae)八角属(Illicium)常绿灌木,是《中国药典》收载的广西特产中药材,也是著名壮药,其茎皮和根皮具有祛风除湿、行气止痛等功效[5],是桂龙药膏、桂龙药酒及多种中成药的主要原料。然而,地枫皮野生资源现处于濒危状态。一方面喀斯特地区生态系统脆弱,生境退化不利于地枫皮的繁育和生长;另一方面人为采挖导致其野生资源急剧下降[6]。
研究喀斯特生境植物地枫皮的适应机理将有助于该物种及该地区生物多样性资源的有效保护和可持续利用。前期研究发现,轻度和中度水分胁迫下地枫皮表现较强的耐旱能力[7],干旱复水后其光合能力迅速恢复[8]。但目前关于地枫皮响应极端干旱方面的研究较少,目前尚不清楚极端干旱下复水的植株是否仍具有较强的恢复能力。植物抗氧化物酶系统迅速响应逆境胁迫,是植物响应非生物逆境(干旱、涝渍、高温、冷害等)的初始应激反应[9],但鲜有研究报道地枫皮抗氧化物酶系统在极端干旱及干旱复水后的响应特征。该研究以1年生地枫皮植株为材料,研究极端干旱及干旱后复水处理对地枫皮叶片含水量和抗氧化酶活性的影响,旨在从抗氧活性的角度揭示地枫皮适应极端干旱的生理机理,以期为喀斯特石漠化地区的生态恢复提供理论基础。
1 材料与方法
1.1 试验材料试验材料为大小长势一致的1年生植株,均由采自广西壮族自治区百色市靖西市同一野生种群的种子萌发而来。栽培容器为营养袋(直径23 cm,高18 cm),每盆装棕色石山土1.7 kg,每盆1株。试验在广西植物研究所的防雨棚中开展。该地区属亚季风气候区,年平均气温23.5 ℃,年无霜期300 d,年日照时数1 680 h,年降水量1 949.5 mm,空气湿度82%。
1.2 试验方法
1.2.1试验设计。试验设3个水分处理,即干旱胁迫、干旱复水和正常浇水,每个处理4株。在统一等量浇水后进行干旱胁迫,以正常浇水(每2 d浇水一次)作为对照。干旱胁迫为不浇水自然干旱,干旱持续47 d后测定叶片含水量和抗氧化酶活性指标。干旱复水为不浇水,干旱持续45 d进行复水处理,浇透,直至盆底有水渗出,复水2 d后测定指标同干旱胁迫处理。
1.2.2叶片相对含水量测定和气孔观察。叶片相对含水量是早晨收获的植株上部第2位和第3位功能叶中测定的。采集材料迅速称叶片鲜重,然后将叶片放入装有蒸馏水的标签袋中浸泡12 h后取出,迅速用纸巾小心地吸干叶片,称取获得叶片饱和鲜重,放入纸袋中,然后置于烘箱50 ℃干燥叶片至恒重称叶片干重。叶片相对含水量=(叶片鲜重-叶片干重)/(叶片饱和鲜重-叶片干重)×100%。
采集当年生成熟叶片,放入FAA固定液中固定,依次用70%、85%、90%、95%、100%乙醇各浸泡1 h脱水,100%乙醇浸泡过夜,CO2临界点干燥仪干燥,离子溅射仪(108Auto Ted Pella,USA)镀金。于卡尔·蔡司扫描电镜(EVO18,German)500倍镜下观察并随机选取5个视野照相。
1.2.3酶液制备和活性测定。干旱复水处理2 d后统一进行采样,取叶位一致且完全成熟叶片迅速放入液氮速冻,置于-20 ℃冰箱保存。称1 g叶片洗净后放入预冷的研钵中,加9 mL pH 7.2~7.4(0.05 mol/L)的磷酸缓冲液,冰浴中充分研磨成匀浆,转入离心管中在4 ℃下2 000 r/min离心20 min,所得上清液即粗酶液。使用ELISA试剂盒测定超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性和丙二醛(MDA)含量。
1.2.4土壤含水量的测定。完成以上指标测定后,称烘干前土壤重量,随后放入铝盒进行烘干至恒重称烘干后土壤重量。土壤含水量=(烘干前土壤重量-烘干后土壤重量)/烘干后土壤重量×100%。
1.3 数据分析采用Excel 2007统计和计算试验数据,SPSS 25.0进行方差分析(ANOVA),采用Duncan法在0.05水平进行多重比较,OriginPro 2021进行绘图。
2 结果与分析
2.1 干旱及干旱复水处理对土壤含水量和叶片相对含水量的影响由图1~2可知,对照处理下的土壤含水量约为35.5%,干旱处理下的土壤含水量降低至10.5%,达到极端干旱水平;干旱胁迫下的地枫皮叶片相对含水量(63.4%)显著低于对照(93.0%)。说明干旱造成了地枫皮植物体内水分亏缺严重,对土壤水分的消耗量增加,导致土壤含水量的降低。复水后,2个参数均显著增加(P<0.05),达到对照水平。此外,极端干旱条件下(土壤含水量10.5%)叶片气孔关闭,表明地枫皮具有较强的水分控制和适应能力(图3)。

注:不同小写字母表示处理间差异显著(P <0.05)。Note:Different lowercase letters indicate significant differences between treatments(P <0.05).图1 干旱复水对土壤含水量的影响Fig.1 Effect of drought rehydration on soil moisture content

注:不同小写字母表示处理间差异显著(P <0.05)。Note:Different lowercase letters indicate significant differences between treatments(P <0.05).图2 干旱复水对地枫皮叶片相对含水量的影响Fig.2 Effects of drought rehydration on leaf relative water content of Illicium difengpi
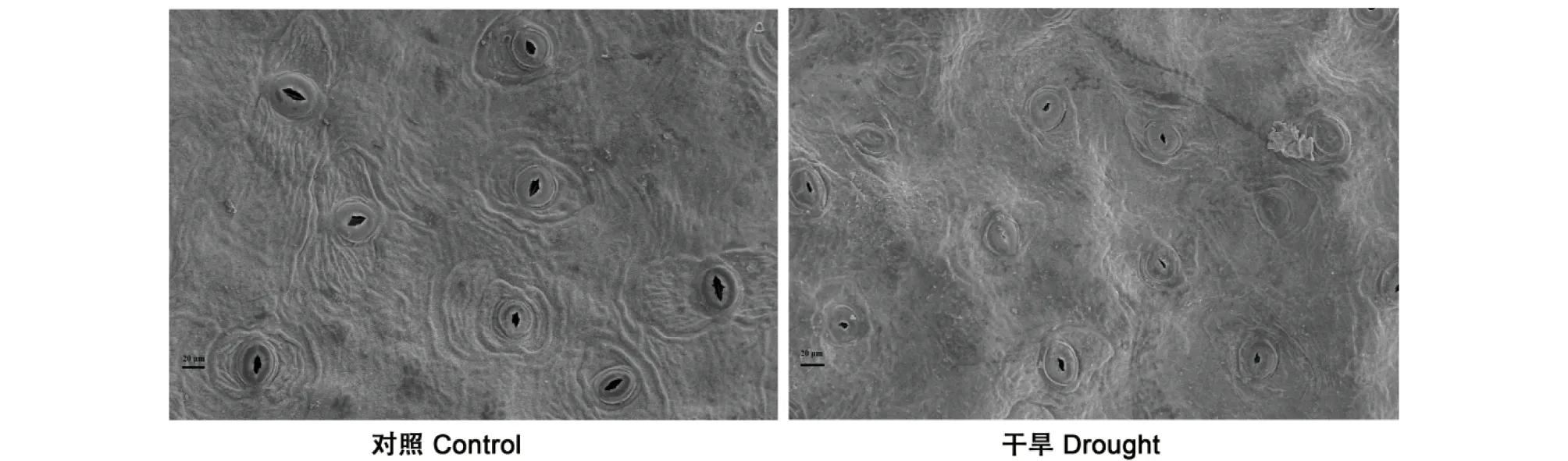
图3 干旱胁迫对地枫皮气孔的影响Fig.3 Effects of drought stress on stomata of Illicium difengpi
2.2 干旱及干旱复水处理对地枫皮MDA含量和抗氧化酶活性的影响MDA是脂质过氧化的产物,膜脂质过氧化的程度可以通过MDA水平反映。由图4可知,对地枫皮进行干旱胁迫后,MDA含量较对照上升了14.5%,但未达到显著差异(P>0.05);复水后,地枫皮叶片MDA含量迅速恢复至对照水平。

图4 干旱复水对地枫皮叶片MDA含量和抗氧化酶活性的影响Fig.4 Effect of drought rehydration on MDA content and antioxidant enzymes activities in leaves of Illicium difengpi
植物体内的抗氧化酶SOD、CAT、POD通过协同作用,清除逆境所产生的活性氧。SOD催化植物体超氧阴离子(O2-)为毒性较小的过氧化氢(H2O2)和O2,进一步通过CAT、POD等其他抗氧化酶清除H2O2。CAT也是植物体内重要的抗氧化酶类之一,它能清除细胞内过多的H2O2,与SOD协同作用清除羟基自由基(·OH)。POD既可将H2O2转化成H2O表现为保护效应,还可参与活性氧的形成表现为伤害效应。干旱处理下,地枫皮的SOD、CAT活性降低,较对照分别降低了4.8%、6.5%(P>0.05),而POD活性显著降低(P<0.05)。复水后,地枫皮CAT活性迅速恢复至对照水平,SOD活性较对照降低了15.2%(P>0.05),POD活性并未恢复,表明POD清除氧的能力出现紊乱。
3 讨论
叶片相对含水量代表植物的实际生长发育状态,反映植物的保水能力[10]。干旱处理的地枫皮叶片含水量下降,相对含水量维持在63.4%,表明干旱处理导致地枫皮细胞失水。干旱胁迫下红树莓(RubusidaeusL.)和钩藤[Uncariarhynchophylla(Miq.)Miq.ex Havil.]的相对含水量均显著降低[11-12]。与其他同属或同种植物如八角(IlliciumverumHook.F.)、莽草(IlliciumlanceolatumA.C.Smith)等相比,在中度或轻度水分胁迫处理下,地枫皮表现出较强的耐旱能力,主要通过调控叶片气孔减少水分流失[7,13-14]。与前人研究不同的是,该研究在极端干旱处理(土壤含水量10.5%)下发现地枫皮仍表现出较强的水分控制和适应能力,叶片气孔关闭的现象。值得注意的是,干旱复水后地枫皮的叶片相对含水量迅速恢复到对照水平,表明地枫皮干旱复水后植株表现出极强的恢复能力,可能是地枫皮耐极端干旱的原因之一。
MDA含量被认为是衡量植物脂质过氧化的一个指标[15]。干旱胁迫下,MDA含量的变化程度可以反映植物的抗旱性强弱[16]。干旱处理诱导地枫皮的MDA含量增加,表明干旱处理下的地枫皮叶片细胞内活性氧的产生超过了抗氧化系统的解毒能力,导致膜脂过氧化,从而产生了MDA,对地枫皮造成了胁迫伤害。崔鹏等[17]研究干旱胁迫下裸果木(GymnocarposprzewalskiiBunge ex Maxim.)MDA含量的变化,也得出相似的结论。但复水后,MDA含量又迅速恢复到对照水平,一定程度上反映了地枫皮较强的抗旱能力。
植物受到干旱胁迫,体内活性氧产物(如O2-和H2O2)的增加会引起抗氧化酶活性的变化。抗氧化酶可以通过解毒过量的活性氧来维持活性氧形成和消除的平衡[9,18]。在酶系统中,SOD转化O2-为H2O2[19],CAT和POD在不同的细胞位置将H2O2分解为H2O[20]。干旱胁迫后,蟛蜞菊[Wedeliachinensis(Osbeck.)Merr.]的SOD、CAT和POD活性均增加[21],党参[Codonopsisradix(Franch.)Nannf.]的SOD活性无明显变化[22];而极端干旱下,火龙果[Hylocereuspolyrhizus(F.A.C.Weber)Britton &Rose]的SOD、POD活性显著降低,CAT活性显著增加[23]。因此,植物抗氧化物酶对干旱响应的差异可能与活性氧产生的强度、胁迫的严重程度和植物种类有关[24-26]。干旱胁迫下,地枫皮的CAT、POD、SOD活性下降,表明在干旱胁迫前期,地枫皮的抗氧化系统先启动,但随着干旱程度加深,膜脂过氧化逐渐加重,活性氧的形成和清除系统的平衡被打破,抗氧化酶防御系统逐渐失去作用,MDA含量开始上升,植株受到一定损害[27]。这与田晓明等[28]对黄山紫荆(CercischingiiChun.)、易家宁等[29]对紫苏[Perillafrutescens(L.)Britt]的研究结果一致。而韩愈[30]研究发现,10%~20%土壤含水量地枫皮幼苗的SOD、POD活性显著升高,这可能与试验材料和干旱胁迫处理时长不同有关。复水后,除POD外,其余参数指标均恢复到对照水平或稍低于对照,说明旱后复水对地枫皮抗氧化系统具有一定补偿作用。而POD活性未恢复,可能是干旱胁迫严重影响了细胞内POD的合成,复水未能缓解所受到的损伤,这提醒在日常的栽培管理中注意浇水,长时间的干旱会对地枫皮幼苗造成一定损害。
4 结论
综上所述,喀斯特极度耐旱植物地枫皮叶片中过氧化物酶系统对干旱处理和干旱后复水处理均表现出一定程度的响应特征,可能参与其适应喀斯特极端干旱环境。该研究初步分析了喀斯特特色药用植物地枫皮对干旱耐性的潜在响应机制,研究结果有望为理解植物耐极端干旱的机制提供新的见解。需要说明的是,该研究仅对1年生地枫皮在干旱和干旱后复水抗氧化系统的水分响应特征进行初步探讨,而地枫皮全生育期的抗旱机理有待进一步研究。
猜你喜欢
森林工程(2018年4期)2018-08-04
时代农机(2018年11期)2018-03-17
山东农业科学(2017年4期)2017-05-18
电源技术(2016年9期)2016-02-27
山东医药(2015年38期)2015-12-07
食品工业科技(2014年13期)2014-03-11
中国氯碱(2014年10期)2014-02-28
食品科学(2013年24期)2013-03-11
食品科学(2013年8期)2013-03-11
