
2021—2022年我国部分地区猪繁殖与呼吸综合征病毒ORF5基因变异分析
2023-10-09 07:08王志远刘博奇许志颖徐思佳邢家宝张桂红孙彦阔
畜牧兽医学报 2023年9期
王志远,刘博奇,许志颖,徐思佳,邢家宝,张桂红,王 衡*,孙彦阔*
(1.华南农业大学兽医学院,广东省临床重大疾病综合防控重点实验室,广州 510642;2.岭南现代农业科学与技术广东省实验室茂名分中心,茂名 525000;3.华南农业大学国家生猪种业工程技术研究中心,广州 510642;4.广东海大畜牧兽医研究院有限公司,广州 511400)
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)在全球范围内普遍流行,俗称蓝耳病,其病原是猪繁殖与呼吸综合征病毒(PRRSV),PRRSV是一种单股正链RNA病毒[1],直径50~65 nm[2],基因组长度约15 kb,编码至少10个开放阅读框(open reading frame,ORF)。PRRSV共分为两种基因型,分别为欧洲型毒株(EU genotype)和北美型毒株(NA genotype),欧洲型毒株即PRRSV-1,代表毒株为Lelystad;美洲型毒株即PRRSV-2,代表毒株为ATCC VR2332[3],两种基因型之间的相似性约为60%[4]。我国境内流行的毒株主要是PRRSV-2,根据ORF5基因分型主要分为4个谱系,即谱系1(NADC30-like毒株和NADC34-like毒株),谱系3(QYYZ和GM2),谱系5.1(VR2332和BJ-4),谱系8.7(CH-1a和JXA1)。1996年我国首次分离到PRRSV,命名为CH-1a,十年后在我国江西省暴发高致病性猪繁殖与呼吸综合征(HP-PRRS),代表毒株为JXA1,后证实为CH-1a的突变毒株[5]。2010年,QYYZ毒株首次在我国大陆地区发现;2012年,GM2毒株被发现并被推测为是疫苗毒株与QYYZ的重组毒株[6]。2013年,NADC30-like毒株在我国首次报道,自传入我国以来流行至今,我国流行的NADC30-like毒株表现为中等毒力,具备强大的遗传进化能力[7]。2018年,我国辽宁省首次检测到NADC34-like毒株,2020年该毒株以后在我国多个省份流行[8]。
PRRSV ORF5和ORF7基因长期以来一直被视为监测PRRSV遗传进化的重要靶基因。ORF5基因序列长度约为603 bp,编码GP5蛋白,该基因拥有重要的中和抗原表位,在诱导免疫应答过程中发挥重要作用,GP5蛋白存在多个潜在的N-糖基化位点,对病毒感染、免疫逃避有着非常重要的作用,因此ORF5是最常被用作分子流行病学监测的基因[9]。ORF7基因序列长度约372 bp,编码N蛋白,该基因相比ORF5较为保守,也常用于PRRSV的临床检测及毒株分型[10],近年来,PRRSV ORF5基因的变异程度日趋复杂,导致ORF5基因序列获取难度加大,因此ORF7基因具备替代ORF5基因作为PRRSV分子遗传变异分析的潜力。自2018年非洲猪瘟在我国暴发以来,不断扩大的疫情对我国养猪行业造成严重的打击,导致PRRSV流行的关注力度明显下降,进而导致我国PRRSV的分子流行病学数据相对较少。本研究在2021—2022年开展了PRRSV分子流行病学调查,通过获取PRRSV阳性样本的ORF5和ORF7基因序列分析PRRSV的主要流行谱系,所有样品涵盖13个省份,为了解近年来我国PRRSV的流行变异情况提供参考依据。
1 材料与方法
1.1 样品收集
2021—2022年,本课题组在我国黑龙江、吉林、河北、山西、河南、四川、贵州、湖南、广西、广东、海南、江西和福建13个省的50个猪场收集了117份疑似PRRS症状样品,样品类型主要为肺、淋巴结、血清。患病猪临床表现为体温升高、厌食、呼吸道症状、生长迟缓、耳鼻发绀、死亡,以及妊娠母猪出现流产、死胎、产木乃伊胎等。
1.2 引物设计
参考NCBI GenBank中收录的JXA1、VR2332、CH-1a等毒株序列,利用Oligo7.0软件分别设计PRRSV ORF5和ORF7基因引物(表1),同时设计PRRSV-1毒株实时荧光定量PCR引物,参照福建省地方标准(DB35/T 1852—2019)设计PRRSV-2实时荧光定量PCR引物,不同基因型引物序列由北京睿博兴科生物技术有限公司合成。

表1 PRRSV ORF5和ORF7引物及扩增片段长度Table 1 PRRSV ORF5 and ORF7 primers and amplified fragment lengths
1.3 样品处理及测序
所有PRRSV阳性样品保存于-80 ℃超低温冰箱,使用超低温组织研磨仪(JXFSTPRP-CLN-48,上海净信)将样品匀浆,使用RNA Fast200(Fastagen)试剂盒提取病毒总RNA,使用qRT-PCR酶(HiScript III U+ One Step qRT-PCR Probe Kit,南京诺唯赞生物科技股份有限公司)鉴定阳性样品及基因型,反应程序:55 ℃ 15 min;95 ℃ 30 s;95 ℃ 10 s、60 ℃ 30 s,共45个循环。将病毒RNA反转录制备cDNA,利用RT-PCR扩增样品ORF7和ORF5基因,反应程序:95 ℃ 3 min;95 ℃ 15 s、60 ℃ 15 s、72 ℃ 60 s,共35个循环;72 ℃ 5 min。PCR产物在10 g·L-1琼脂糖凝胶中进行电泳,电泳结果在凝胶成像系统中进行可视化分析,将条带大小位置一致的扩增产物进行Sanger测序以获取目的序列。
1.4 系统发育学分析及基因变异分析
应用DNA STAR 7.1软件和MAFFT 7将PRRSV ORF5和ORF7基因序列进行比对,利用Megalign软件分析不同毒株ORF5和ORF7基因的序列相似性。利用IQ-Tree软件构建最大似然系统发育树,应用Figtree v1.4.3软件将系统发育树进一步可视化分析。利用EditSeq软件将PRRSV ORF5基因核苷酸序列转换为氨基酸序列,利用Jalview软件分析PRRSV GP5氨基酸突变位点。
2 结 果
2.1 2021—2022年我国部分地区PRRSV临床样品检测结果
2021—2022年,本研究共收集疑似PRRS症状患猪样品117份,其中大部分样品来自于华南地区,部分样品来自于我国西南和中部及北部地区(表2),经实时荧光定量PCR鉴定48份样品为PRRSV阳性,且均为PRRSV-2毒株,应用 RT-PCR成功扩增出38份样本ORF5基因,使用ORF7引物成功扩增出剩余10份样品的ORF7基因。

表2 2021—2022年PRRSV临床样品收集地分布表Table 2 Geographic distribution of PRRSV clinical sample collections, 2021—2022
2.2 PRRSV ORF5和ORF7基因相似性分析
以PRRSV-2不同谱系代表毒株作为参考毒株,38条ORF5基因序列之间核苷酸相似性为77.1%~99.8%,与 NADC30相似性为82.4%~95.2%,与NADC34相似性为81.9%~96%,与QYYZ相似性为81.1%~91.4%,与GM2相似性为80.4%~90.7%,与VR2332相似性为81.4%~99.3%,与CH-1a相似性为82.9%~96.5%,与JXA1相似性为82.3%~97.7%;其中,GDHZ29与IA/2014/NADC34的相似性高达96%(图1A)。10条ORF7序列间核苷酸相似性为83.3%~98.7%,与NADC30相似性为86%~96.8%,与NADC34相似性为84.1%~96%,与QYYZ相似性为85.2%~89.8%,与GM2相似性为84.4%~88.4%,与VR2332相似性为87.9%~93.3%,与CH-1a相似性为86.8%~98.1%,与JXA1相似性为86%~98.9%(图1B)。
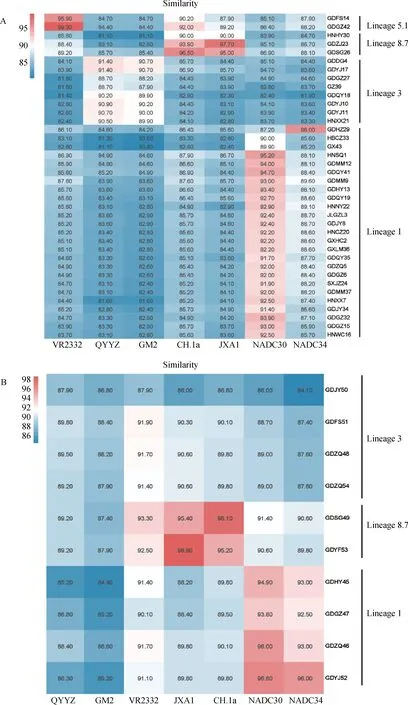
图1 PRRSV ORF5(A)及ORF7基因(B)与不同谱系参考毒株基因相似性分析Fig.1 Gene similarity analysis of ORF5 and ORF7 genes with reference strains of different lineages
2.3 PRRSV ORF5和ORF7基因系统发育学分析
ORF5基因系统发育学分析表明,所有毒株均属于PRRSV-2,进而划分为4个分支:谱系1(谱系1.8 NADC30-like毒株24株,谱系1.5 NADC34-like毒株1株),谱系3(QYYZ-like毒株和GM2-like毒株 8株),谱系5.1(VR2332-like毒株2株),谱系8.7(CH-1a-like毒株 2株和JXA1-like毒株 3株)(图2)。ORF7基因系统发育学分析同样将所有毒株分为4个分支,其中4株属于谱系1,4株属于谱系3,2株属于谱系8.7(图3)。根据ORF5和ORF7基因系统发育学分析可知,谱系1毒株共29株(60%),占比最高且地理分布最广,共在8个省份(8/9)均有检出;谱系3毒株12株(25%),主要在华南地区检出;谱系5.1毒株2株(5%),均在广东省检出;谱系8.7毒株5株(10%),均在华南地区检出(图4,表3)。
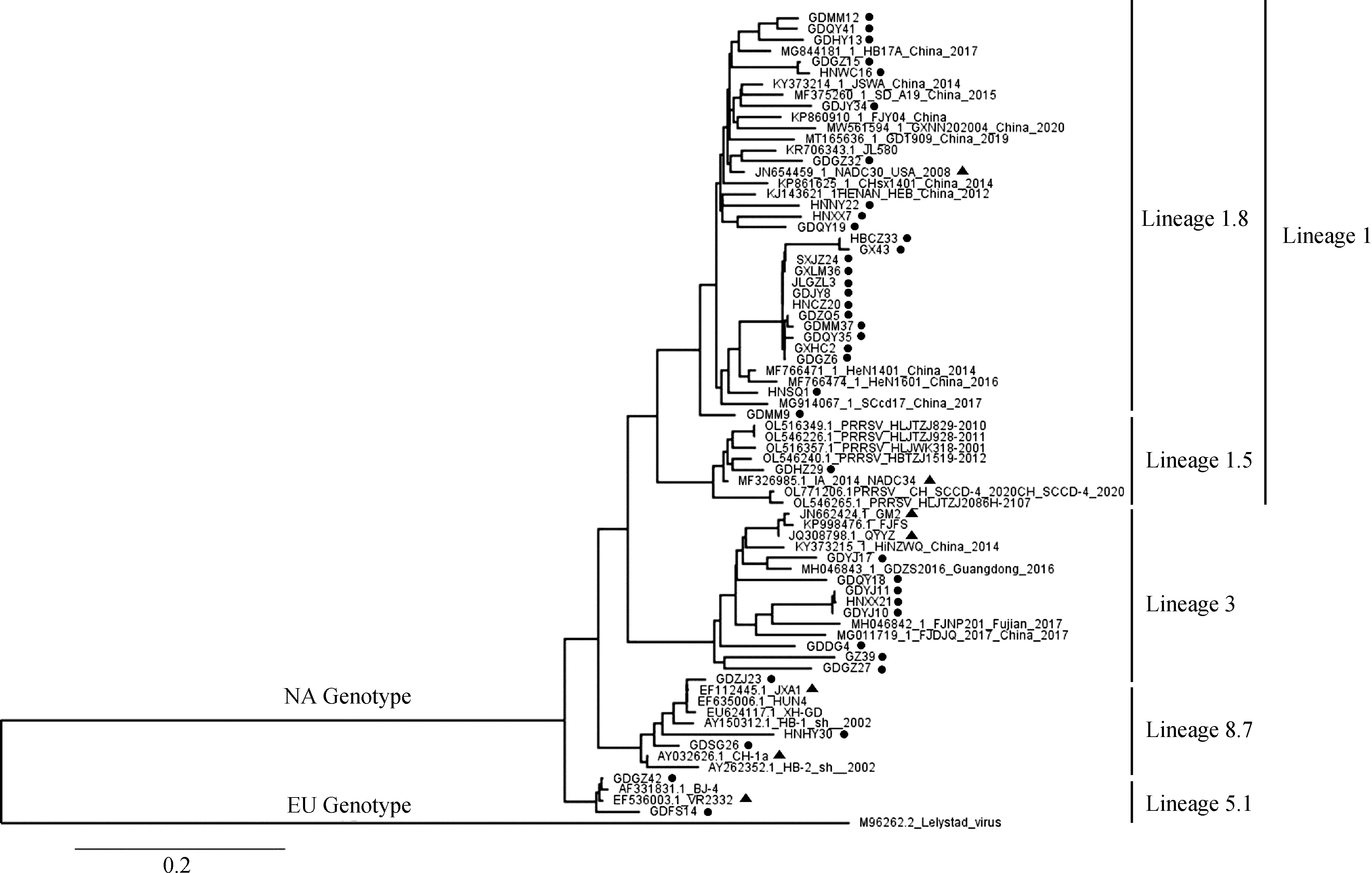
▲. 不同谱系代表毒株;●. 本研究中的毒株▲. Representative strains of different lineage; ●. The strains of this study图2 基于PRRSV ORF5基因的系统发育学分析Fig.2 Phylogenetic analysis based on PRRSV ORF5 gene

▲. 不同谱系代表毒株;●. 本研究中的毒株▲. Representative strains of different lineage; ●. The strains of this study图3 基于PRRSV ORF7基因的系统发育学分析Fig.3 Phylogenetic analysis based on PRRSV ORF7 gene

图4 2021—2022年PRRSV不同谱系毒株的检出率Fig.4 Detection rates of different strains of PRRSV lineage from 2021 to 2022

表3 PRRSV不同谱系毒株地理分布表Table 3 Geographical distribution of different strains of PRRSV
2.4 PRRSV GP5氨基酸位点分析
GP5氨基酸序列分析表明信号肽(aa 1—26)、潜在N-糖基化位点(aa 32—35、aa 44、aa 51)及表位C(aa 52—61)区域氨基酸突变程度最复杂,其他区域的氨基酸突变均具有谱系一致性(图5、6)。以VR2332为参考毒株,不同谱系毒株存在个别氨基酸位点相似性(谱系1、谱系3、谱系8.7毒株大部分均为E3G、S16F、A29V、D34 N、F127 L、A137S、R165 G、V185A)。GP5潜在的N-糖基化位点共有6个,其中2个N-糖基化位点44和51相对保守,4个N-糖基化位点32、33、34、35处于高变区,谱系5.1毒株存在2种点突变(S32 N、D34 G),谱系8.7毒株存在6种点突变(S32 N、N33S、D34S/N、S35 N/H),谱系3毒株存在6种点突变(S32 N/E、N33 G/Q/T、D34 N),谱系1毒株存在5种点突变(S32 G/D、N33K、D34 H、S35 N),在表位C区域(aa52-61 中和表位),谱系5.1存在3种点突变(D54E、A57 G、K59 N),谱系8.7存在5种点突变(A57 N、N58Q/D、K59 H、D61S),谱系3存在11种点突变(A57 N、N58 T/K/D/S/E、K59 N/R/S、D61E/Y)。

红色框. 信号肽;黑色框. 跨膜区;黄色框. T细胞表位;黑色曲线框. 潜在的N-糖基化位点;黑色阴影框.非中和表位和中和表位区域Red box. Signal peptide; Black box. Transmembrane region; yellow box. T cell epitope; Black curved box. Potential N-glycosylation site; Black shaded box. Non-neutralizing epitope and neutralizing epitope region图5 谱系3、谱系5.1和谱系8.7 PRRSV GP5氨基酸位点变异分析Fig.5 Analysis of amino acid residue variants in PRRSV GP5 in lineage 3, lineage 5.1 and lineage 8.7

红色框. 信号肽;黑色框. 跨膜区;黄色框. T细胞表位;黑色曲线框. 潜在的N-糖基化位点;黑色阴影框.非中和表位和中和表位区域Red box. Signal peptide; Black box. Transmembrane region; yellow box. T cell epitope; Black curved box. Potential N-glycosylation site; Black shaded box. Non-neutralizing epitope and neutralizing epitope region图6 谱系1 PRRSV GP5氨基酸位点变异分析Fig.6 Analysis of amino acid residue variants in PRRSV GP5 of lineage 1
3 讨 论
自PRRSV首次在我国检出以来已有20余年流行史,其遗传进化历程总体分为三个阶段:第一阶段为1996—2006年,优势流行毒株为CH-1a;第二阶段为2006—2013年,优势流行毒株为JXA1;第三阶段为2013年至今,优势流行毒株为NADC30-like[6]。尽管我国进行了广泛的疫苗免疫,由于PRRSV基因组的高度变异性导致新毒株层出不穷,使得常规疫苗的交叉保护效力并不理想。2018年以来,非洲猪瘟疫情的暴发给我国生猪养殖业带来了严重打击,同时使我国养猪业生物安全防护意识极大提高。然而,由于PRRSV自身遗传进化、对外频繁引种、疫苗使用不当等原因导致猪群免疫选择压力增大,使PRRSV得以传入并迅速发生遗传变异;此外,非洲猪瘟大流行背景下使养殖户对PRRSV防控意识下降,伴随着全国范围内生猪大批量复养,猪存栏数量剧增,使PRRSV得到了再一轮的流行[11]。因此,即便在非洲猪瘟流行的大背景下,PRRSV的流行力度未能降低[12]。同时,由于对PRRSV关注力度下降,导致PRRSV分子流行病学调查的数据相对较少。开展PRRSV流行病学调查研究有重要意义,流行病学调查将有助于了解PRRSV最新的遗传变异情况。在2021—2022年,本研究共收集了117份临床疑似PRRSV感染样品,经实时荧光定量PCR检测均为PRRSV-2,总体阳性率为41%(48/117),ORF5和ORF7基因系统发育分析将所有的序列分为4个谱系(谱系1、谱系3、谱系5.1、谱系8.7),谱系1毒株数量最多(29/48),其次是谱系3(12/48),谱系8.7(5/48)和谱系5.1(2/48)少量检出。有研究表明NADC30-like毒株已成为华中地区PRRSV主要流行谱系[13];福建省PRRSV主要流行毒株是以NADC30-like毒株为主要亲本的重组病毒[14];综合本研究结果显示NADC30-like毒株已代替HP-PRRSV成为全国范围内PRRSV主要流行毒株,NADC30-like毒株基因组变异程度极高,表现出强大的基因重组能力,有报道称NADC30-like毒株极易与HP-PRRSV发生重组,从而增加了田间流行毒株的复杂性,导致不同毒力的新毒株出现[3,15],NADC30-like毒株的广泛流行是当下我国PRRSV防控需要迫切关注的问题。谱系3毒株目前依旧在华南地区广为流行,有研究表明谱系3毒株的核苷酸变异率为9.85×10-3位点·年-1,近几年与谱系8.7毒株重组导致的高毒力毒株在华南地区广泛流行[16]。谱系5.1毒株多为疫苗毒株,与临床疫苗(Resp PRRS MLV)广泛使用有密切的联系。NADC34-like毒株在我国出现时间相对较晚,但2020年以来在全国范围内波及的区域越来越广[17],本研究于2021年在广东省惠州市检出一株NADC34-like毒株,与IA/2014/NADC34相似性为96%,证明NADC34-like毒株由我国北方地区(辽宁省、黑龙江省、河南省)[8]传入华东(福建省)和华南地区(广东省),结合相关报道可知NADC34-like毒株已经在广东省发生潜在流行[18]。
PRRSV GP5与病毒免疫逃避及免疫保护密切相关,GP5蛋白区域的突变影响信号肽的裂解、N-糖基化位点的数量以及中和抗体对PRRSV变异体的识别[19-20]。GP5氨基酸突变分析表明,广泛的突变位点分布在整个基因,信号肽、中和表位C及潜在的N-糖基化位点三个区域的突变程度最显著,不同数量的N-糖基化位点可能会影响病毒复制和诱导中和抗体的能力,并可能有助于病毒通过聚糖屏蔽逃避抗体中和[21], 表位C区域的氨基酸突变可能导致同源中和失效[22]。
综上所述,当下我国PRRSV的流行谱系以谱系1为主,同时谱系3和谱系8.7共存。其中,NADC30-like毒株检出率最高且流行区域最为广泛,NADC34-like毒株已经在华南地区出现并流行,多谱系毒株共存及谱系1毒株的广泛流行的现状给国内PRRSV的防控工作带来了巨大的挑战。NADC30-like毒株传入我国以来,遗传进化速度极快,极易与不同谱系毒株发生基因重组,导致PRRSV不同毒株的基因分型更加难以定性[23-24]。同时,我国PRRSV常规疫苗大多基于谱系8毒株研制,对于复杂多变的谱系1毒株的交叉保护效果不佳,不同疫苗的滥用也使得PRRSV控制难度加大[25]。同时,本研究发现谱系3毒株依然局限在华南部分地区流行,但流行率逐年增高,需要我们重视该谱系毒株的流行程度并及时加以控制。面对国内PRRS流行日趋复杂的现状,需要人们更大力度建立生物安全防控,以及加强流行病学调查用以动态分析PRRSV的遗传变异情况。本研究为国内PRRS流行现状给出了最新的数据支撑,也给未来临床防控策略制定提供了参考依据。
4 结 论
多个谱系猪繁殖与呼吸综合征病毒(PRRSV)同时在我国流行,谱系1已替代谱系8.7成为主要流行谱系,并分布于我国大多数省份;谱系3仅次于谱系1,主要在华南局部地区流行;谱系8.7毒株检出率较低。近几年来,PRRSV在我国的流行日趋复杂,谱系1毒株的广泛流行增加了PRRSV的遗传多样性。
猜你喜欢
数学物理学报(2022年5期)2022-10-09
艺术品鉴(2022年16期)2022-07-09
现代畜牧科技(2021年6期)2021-07-16
河池学院学报(2021年1期)2021-07-10
景德镇陶瓷(2021年1期)2021-03-24
河北画报(2020年8期)2020-10-27
东方考古(2017年0期)2017-07-11
浙江大学学报(工学版)(2016年2期)2016-06-05
医学研究杂志(2015年12期)2015-06-10
中国粮油学报(2014年7期)2014-02-06
