
干旱胁迫下不同性别野牛草生理及抗氧化酶基因表达差异
2023-10-21 05:41刘牧野郭丽珠岳跃森武菊英范希峰肖国增滕珂
草业学报 2023年10期
刘牧野,郭丽珠,岳跃森,武菊英,范希峰,肖国增*,滕珂*
(1. 长江大学园艺园林学院,湖北 荆州 434052;2. 北京市农林科学院,草业花卉与景观生态研究所,北京 100097)
干旱是制约植物生长的重要胁迫因素之一[1]。对于草坪草而言,草坪坪观质量是其主要的观赏价值之一。干旱胁迫会通过引起植物功能的改变使草坪质量严重下降,影响其景观和生态功能[2]。干旱胁迫会导致植物体内产生大量的活性氧(reactive oxygen species,ROS)[3],ROS 过量积累会导致植物细胞死亡,进而使植物受到损害[4]。植物体内存在一套由抗氧化酶以及非抗氧化酶组成的抗氧化系统,从而抵御ROS 对植物的伤害[5]。抗氧化酶系统包括超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)、过氧化氢酶(catalase,CAT)、抗坏血酸过氧化物酶(ascorbate peroxidase,APX)等[6]。SOD 作为植物逆境防御的第一道防线,保护细胞免受超氧化物诱导的氧化胁迫[7];APX、CAT 则清除由SOD 特异性地歧化超氧阴离子(O-2)产生的H2O2[8]。Mittler[9]发现APX 与CAT 对H2O2的清除机制不同,APX 负责调节ROS 中间体的信号传导,而CAT 负责清除过量的ROS。POD 作为活性较高的适应性酶,能够反映植物生长发育的特点、体内代谢状况以及对外界环境的适应性[10]。
干旱胁迫不仅使植物抗氧化酶活性在生理水平发生变化,在分子水平上也会发生抗氧化酶基因表达量的变化。Nasirzadeh 等[11]研究了干旱胁迫对3 种耐旱性小麦(Triticum aestivum)的抗氧化酶基因表达的影响 ,结果表明3 种小麦抗氧化酶相关基因在干旱胁迫条件下的表达都增强。田又升等[12]研究表明耐旱型棉花(Gossypiumspp.)相较于干旱敏感型棉花在干旱胁迫下抗旱相关基因表达量更高。Zhang 等[13]研究了干旱胁迫下白三叶(Trifolium repens)抗氧化酶基因的表达模式,结果表明白三叶在干旱3 h 后CAT、Fe-SOD、Mn-SOD、Cu/Zn-SOD表达量迅速上升。连玲等[14]研究水稻(Oryza sativa)在干旱胁迫下抗氧化酶基因的表达水平变化发现,干旱导致水稻的抗氧化酶基因表达量上调,且大部分基因都呈先上升后下降的趋势。
野牛草(Buchloe dactyloides),隶属禾本科野牛草属,是一种重要的多年生暖季型草坪草[15]。野牛草因为其形态柔顺、抗逆性强、耐贫瘠等优点在我国广泛栽培[16]。前人研究表明野牛草是所有草坪草中抗旱性最强的物种[17],研究其抗旱机理对其他草坪草抗旱性研究具有指导意义。雌雄异株植物是指只具有单一性别的单性花植株[18],被子植物中只有6%的物种是雌雄异株[19]。野牛草是草坪草植物中少有的雌雄异株植物[20],并且前人研究发现除了在抗叶锈病 (Puccinia kansensis)[21]、匍匐茎生长能力[22]以及保水能力[23]外,二者在扩繁、生长习性、生物产量上并无太大差异。目前对于不同性别野牛草抗旱的生理及分子差异机制的研究较少,本研究旨在通过对生理以及分子水平野牛草雌、雄株之间抗旱能力的比较,探究野牛草雌、雄株之间的抗旱性强弱;同时明确野牛草雌、雄株之间抗氧化酶活性及基因表达在抗旱中的差异,以期为研究雌雄异株植物的抗旱机制提供参考。
1 材料与方法
1.1 试验材料
试验植物为野牛草‘Texoka’品种,2020 年8 月在北京市农林科学院草业花卉与景观生态研究所温室播种。2021 年6 月植物开花,根据野牛草雌、雄株开花后花型不同从而分辨性别后进行雌、雄株分株扩繁培养至2022 年2 月获得遗传背景一致、基因型相同的单性别无性系群体。
1.2 试验设计
于 2022 年 6 月25 日在北京市农林科学院智能温室中进行移苗盆栽试验,采用顶部直径19 cm、底部直径10 cm、高度15 cm 的花盆,以野牛草雌株或雄株无性种苗分蘖6~7 个为一株进行栽植,土壤采用园土。缓苗至2022年9 月22 日后进行干旱处理,2022 年10 月6 日进行复水。本研究设置2 个性别分组雌(♀)与雄(♂),每个性别设置4 种干旱处理,分别为:D0d(干旱处理0 d)、D7d(干旱处理7 d)、D14d(干旱处理14 d)、R7d(复水处理7 d),每个处理3 个生物学重复。干旱胁迫前,每盆浇水至底部渗水,干旱胁迫后不浇水,在干旱胁迫第14 天取样后浇水至底部渗水进行复水,复水7 d 内不浇水。在干旱至0、7、14 d、复水7 d 的下午3:00 进行取样,叶片鲜样用于测定叶片相对含水量以及叶绿素,其余指标采用液氮以及-80 ℃冰箱保存样进行测定。
1.3 试验方法
1.3.1 叶片相对含水量测定 称取样品1 g,记录其鲜重(fresh weight,WF);将称好的叶片放入50 mL 三角瓶中,加满蒸馏水,封口,放置24 h;取出叶片,快速轻轻擦干表面水分,称其饱和重(saturation weight,WT);将饱和叶片放入铝盒内,105 ℃杀青15 min;80 ℃烘至恒重;取出后,自然冷却,称干重(dry weight,WD)。计算公式:
1.3.2 叶绿素含量测定 称取野牛草雌、雄株叶片0.05~0.08 g,记下具体质量,剪碎后置于离心管中;加入8 mL 95%的乙醇,避光静置 24 h;于665 和649 nm 下测定其吸光度值A665、A649;95%乙醇调整。计算叶绿素含量:
式中:V为提取后的体积(L,0.008 L);W为称取的叶片质量(g)。
1.3.3 PSII 最大光化学量子效率测定 采用Handy PEA 植物效率分析仪(Handy PEA+、PP SYSTEMS,英国)进行PSII 最大光化学量子效率(the maximum quantum yield of PSII,Fv/Fm)测定,用配备的暗适应夹对叶片进行充分的暗适应(30 min),将探头置于暗适应夹上的圆形槽之中,确保探头与暗适应夹紧密接触,无光线进入。用手按紧探头与暗适应夹,拉开适应夹上金属遮光片,按探头一侧的快捷键,开始测定。测定结果通过PEA软件进行分析总结。选取每盆野牛草雌、雄株叶片中部4 个部位使用暗适应夹进行暗适应,结果取4 个暗适应部位数据平均值。测定时间为每次取样当天的上午10:00。
1.3.4 抗氧化酶测定 用苏州科铭生物技术有限公司试剂盒(SOD-1-Y、POD-1-Y、CAT-1-Y、APX-1-W),依据试剂盒说明书方法,测定0.1 g 野牛草健康生长叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX),分别用酶标仪(Multiskan FC,Thermo,美国)、紫外分光光度计(UV2600,Techcomp,中国)在520、560、470、240 和290 nm 波长下测量吸光值并计算酶活性。
1.3.5 抗氧化酶基因表达量测定 使用百迈客公司RNA 试剂盒(RK02004)提取健康生长的野牛草叶片总RNA,用Takara 公司反转录酶试剂盒(RR036A)在PCR 仪中(C1000,Bio-Rad,美国)反转录成cDNA。反应体系为20 μL,反应条件:Lid 50 ℃,37 ℃ 30 min,反应后 85 ℃ 热变性5 s,4 ℃保存,以反转录获得的cDNA 为模板,用Actin引物进行PCR 检测,电泳检测有单一条带后,用于后续的qRT-PCR 试验。
根据前期实验室获得的野牛草全长转录组数据设计野牛草SOD(Cu/Zn-SOD、FeSOD)、APX、CAT、POD的qRT-PCR 引物(表1)。利用Bio-Rad CFX connect 荧光定量仪(美国)进行qRT-PCR 反应。SYBR Mix 采用Takara 公司的试剂(RR820A)。反应体系为20 μL,反应条件:95 ℃ 30 s ,95 ℃ 5 s,60 ℃ 30 s,40 次循环。定量表达结果依据CT 数值,根据 2-ΔΔCt法[24]分别计算目标基因在不同处理下的相对表达量。
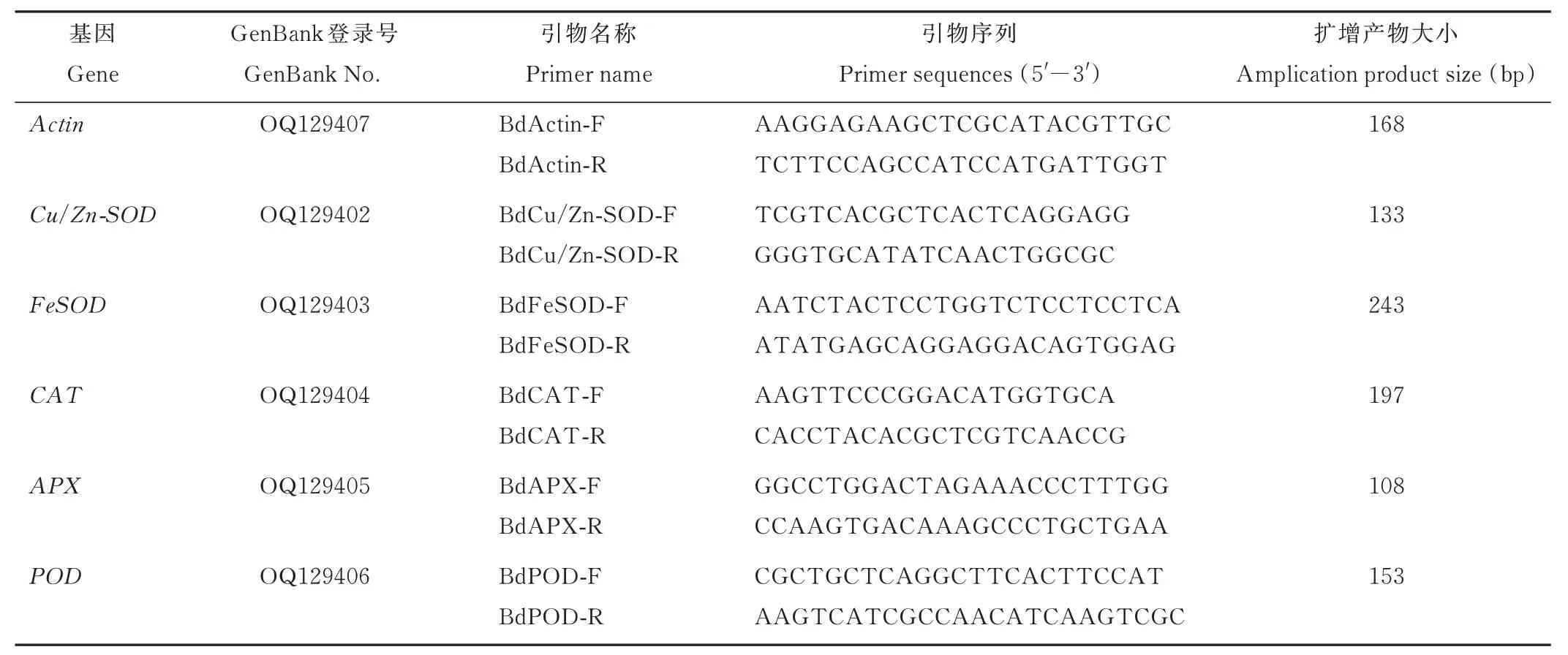
表 1 实时荧光定量引物信息Table 1 Primer information for qRT-PCR analysis
1.4 数据处理
利用Excel 2019 对原始数据进行记录整理,采用SPSS 27. 0 对数据进行显著性分析,叶绿素荧光数据采用PEA 软件进行分析,使用GraphPad Prism 9.4 作图。
2 结果与分析
2.1 干旱胁迫对野牛草草坪质量的影响
野牛草雌、雄株叶片随着干旱时间的增加开始变黄萎蔫。第7 天野牛草雌株叶片未发生明显变化;雄株少部分叶片前端开始变黄萎蔫。第14 天雌、雄株叶片都明显变黄,叶片前端发生弯曲萎蔫现象,大部分叶片蜷缩。复水后,雌株大部分叶色恢复,叶片舒展,但叶片前端仍呈现黄色弯曲现象,少部分叶片枯黄无法恢复;雄株叶片仍呈现黄色萎蔫状,且较多数叶片枯黄死亡无法恢复(图1)。以上结果表明,野牛草雌株与雄株相比具有较好的耐旱性和抗旱恢复潜力。

图1 干旱胁迫条件下野牛草雌、雄株表型的变化Fig.1 Phenotypic changes of female and male buffalograss plants under drought stress
2.2 干旱胁迫对野牛草叶片含水量和光合作用的影响
干旱胁迫降低了野牛草雌、雄株的叶片RWC 和Chl 含量,随着干旱时间的增加,野牛草雌、雄株的RWC 都呈下降趋势,干旱第7 天和干旱第14 天与第0 天相比都有显著性差异,复水后RWC 与干旱第14 天相比显著上升(图2A)。野牛草雌株Chl 随着干旱时间的增长而下降,干旱第14 天与干旱第0 天相比显著下降,复水后仍显著下降(图2B);野牛草雄株Chl 随着干旱时间的增长呈先上升后下降的趋势,干旱第14 天与干旱第0 天相比显著下降,复水后仍呈下降趋势,与第0 天相比有显著差异(图2E)。随着干旱胁迫时间增加,野牛草雌、雄株Fv/Fm都在第14 天显著下降,复水后回升(图2C 和F)。
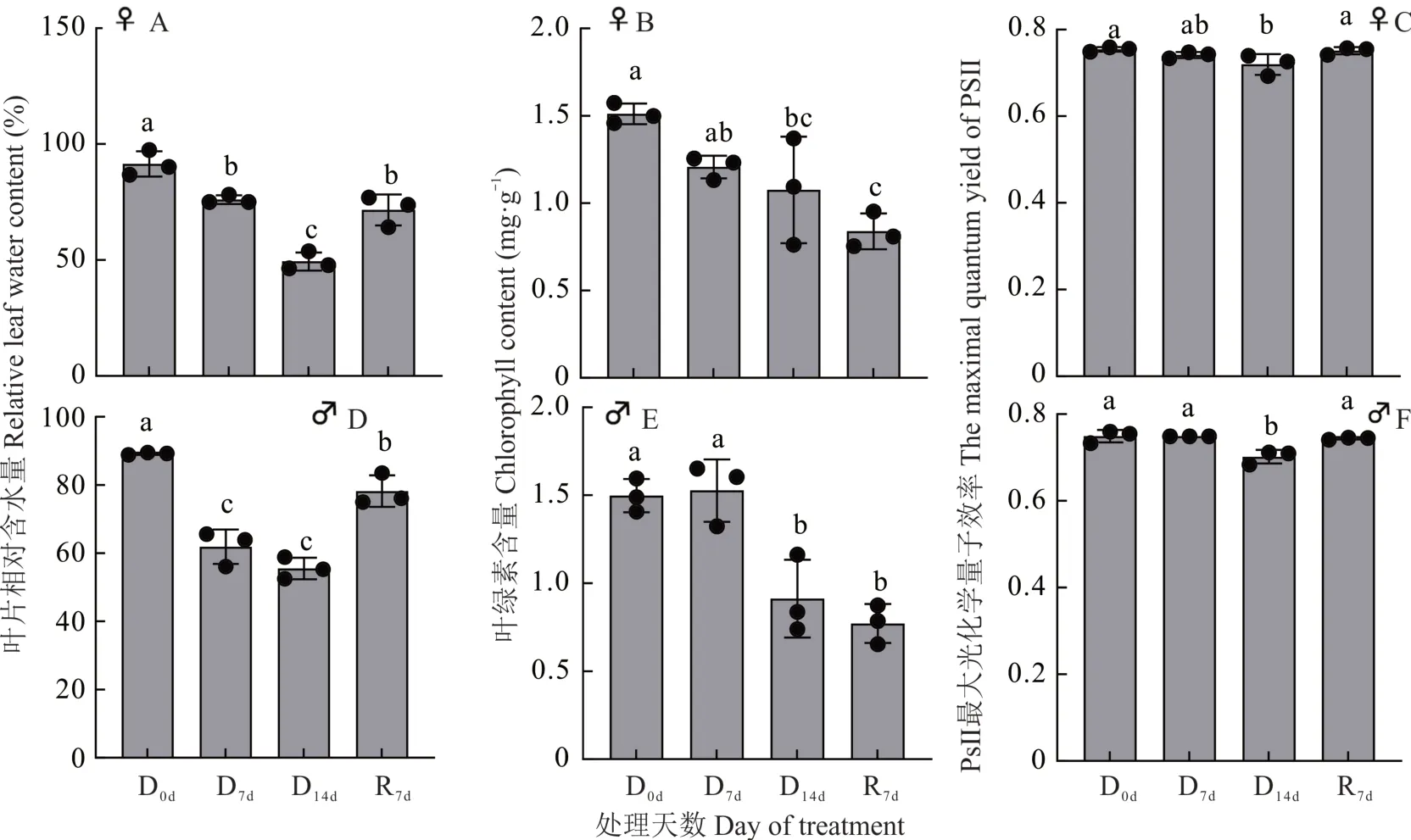
图2 干旱胁迫条件下野牛草雌、雄株叶片相对含水量、PSII 最大光化学量子效率以及叶绿素含量的变化Fig. 2 Detection of relative leaf water content,the maximum quantum yield of PSII and chlorophyll content of female and male buffalograss under drought stress
2.3 干旱胁迫对野牛草抗氧化酶活性的影响
干旱胁迫条件下,野牛草雌株SOD 活性在干旱第14 天显著上升,复水后相较于干旱第14 天显著下降(图3A);野牛草雄株SOD 活性随着干旱时间的增加呈先下降后上升的趋势,复水后显著下降(图3E)。野牛草雌株在干旱处理下APX 活性呈先下降后上升的趋势,在干旱第14 天达到最高,显著高于第0 和7 天,复水后又下降(图3B);野牛草雄株APX 活性随着干旱时间的增加而上升,在第14 天达到峰值,显著高于第0 天以及第7 天,复水后APX 活性显著下降,显著低于干旱条件下APX 活性(图3F)。野牛草雌株CAT 活性在干旱条件下呈先上升后下降的趋势,复水后又显著上升(图3C);雄株在整个处理过程中呈先下降后上升又下降的趋势,干旱第0 天显著高于其他处理时间(图3G)。野牛草雌、雄株POD 活性整体趋势相同,呈先上升后下降的趋势,在干旱第14 天达到峰值,显著高于其他处理时间(图3D 和H)。

图3 干旱胁迫条件下野牛草抗氧化酶活性的检测Fig.3 Detection of antioxidant enzyme activity in buffalograss under drought stress
2.4 干旱胁迫对野牛草抗氧化酶系统相关基因的影响
随着干旱时间的延长,野牛草雌株FeSOD的表达水平没有显著变化,复水后显著上升(图4A);雄株FeSOD的表达水平随着干旱时间的延长先下降后上升,复水后下降,干旱第14 天达到峰值,显著高于其他处理时间(图4F)。野牛草雌株Cu/Zn-SOD的表达水平随干旱时间的延长呈先下降后上升的趋势,在复水后达到峰值,显著高于干旱处理7 和14 d(图4B);雄株Cu/Zn-SOD的表达水平随着干旱时间的延长先下降后上升,复水后下降,干旱第14 天达到峰值(图4G)。野牛草雌株APX的表达水平随着干旱时间的延长先下降后上升,复水后持续上升,在复水7 d 后达到峰值且显著高于其他处理时间(图4C);雄株APX的表达水平随着干旱时间的延长呈下降趋势,复水后上升(图4H)。野牛草雌、雄株CAT的表达水平均先下降后上升,其中雌株CAT表达水平复水后达到峰值(图4D)、雄株CAT表达水平在干旱第0 天最高(图4I)。野牛草雌株POD的表达水平先下降后上升,复水后达到峰值(图4E);雄株POD表达水平则在干旱条件下先下降后上升,复水后下降,在干旱第14 天达到峰值且显著高于其他处理时间(图4L)。
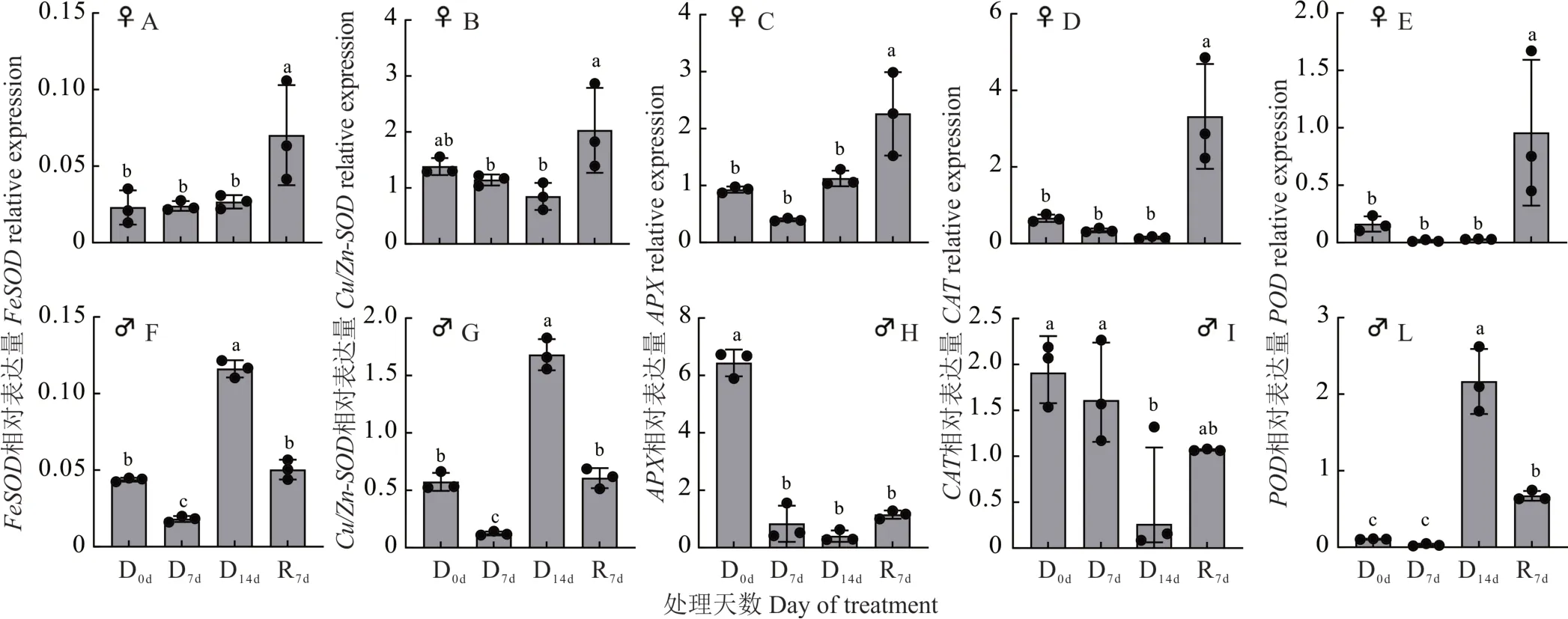
图4 干旱胁迫条件下野牛草抗氧化酶系统相关基因相对表达量检测Fig.4 Detection of relative expression levels of genes related to antioxidant enzyme system in buffalograss under drought stress
2.5 野牛草雌雄株对干旱胁迫的差异性分析
通过SPSS 软件对同一处理天数下野牛草雌、雄株之间差异进行独立样本T检验得到表2。由表2 可知,在干旱处理0 d 时,FeSOD基因相对表达量在野牛草雌、雄株之间差异显著,APX、CAT、Cu/Zn-SOD基因表达量以及CAT 活性存在极显著差异。干旱处理第7 天时,野牛草雌雄株之间Chl、APX 活性以及CAT基因相对表达量差异显著,RWC、CAT、SOD 活性以及Cu/Zn-SOD基因相对表达量存在极显著差异。干旱第14 天时,CAT 活性差异显著,SOD 活性、Cu/Zn-SOD、FeSOD、POD以及APX基因相对表达量存在极显著差异。复水7 d 后,POD、SOD 以及CAT 活性存在极显著差异,Cu/Zn-SOD基因相对表达量差异显著。从表2 还可以看出,除了PSII 最大光化学量子效率外,其他指标均有至少一个处理天数在雌雄株之间存在显著差异。

表2 野牛草雌、雄株同一处理天数之间的差异Table 2 Difference between female and male buffalograss for the same treatment days
3 讨论
草坪质量会随着干旱胁迫的持续而下降[25],本研究中野牛草雌、雄株叶片都随着干旱时间的增加而变黄萎蔫,但是在复水后野牛草雌株叶片恢复状态更好,因此从表型上看,野牛草雌株的抗旱性要强于雄株。叶片相对含水量是植物组织实际含水量占组织饱和含水量的百分比,常用来作为植物在遭受水分胁迫后的水分亏缺的参数[26]。在干旱胁迫对草原六月雪(Koeleria macrantha)影响的研究中,两种不耐旱型品种RWC 显著低于耐旱型品种[27]。本研究中野牛草雌雄株RWC 都随着干旱时间增加而下降,复水后上升,在干旱第7 天雌株RWC 显著高于雄株,证明在短期干旱胁迫下野牛草雌株相较于雄株可能更加耐旱。干旱胁迫会导致植物叶绿素含量下降[28],一般来说叶绿素含量越高表明植物光合能力及抗性越强[29]。本研究中野牛草雌雄株Chl 随着干旱-复水处理整体都呈下降趋势,但是雌雄株之间除第7 天外均没有显著差异。金祎婷等[30]研究在全生育期干旱胁迫对‘青燕1号’燕麦(Avena sativa)PSII 最大光化学量子效率影响时发现,干旱胁迫程度越高,对‘青燕1 号’Fv/Fm影响越大,且不同干旱胁迫程度下,其光合作用限制因素不同,重度胁迫下限制因素为非气孔因素,主要表现为 PSⅡ反应中心失活,光合电子传递受阻,其Fv/Fm数值显著下降。本研究中野牛草雌、雄株Fv/Fm在干旱-复水处理过程中都呈相同的趋势,在干旱第14 天都显著降低,二者之间没有显著差异,Chl 含量和Fv/Fm的变化表明本研究中干旱处理抑制了野牛草雌、雄株的光合性能,但雌株和雄株之间的差异并不明显。
抗氧化酶系统是植物应对逆境胁迫的重要机制之一[31]。除CAT 活性外,野牛草雌、雄株抗氧化酶活性在干旱第7 天后都呈先上升后下降趋势,且不同处理天数之间差异显著,证明干旱胁迫下野牛草抗氧化酶都正常发挥作用。野牛草雌、雄株SOD 活性在干旱第7、14 天以及复水7 d 后具有显著差异,且雌株活性高于雄株,证明野牛草雌株O-2的歧化能力强于雄株。这与马少薇等[32]的研究相同,其研究结果表明干旱胁迫下黄柳(Salix gordejevii)雌株比雄株SOD 活性更强。野牛草雄株APX 活性在干旱第7 天显著高于雌株,但雌雄株在干旱第14天以及复水7 d 没有显著性差异,在野牛草应对长时间干旱时,雌株APX 活性变化更加明显,表明雌株应对干旱胁迫时APX 更快积累,应对长时间干旱胁迫的能力可能更强。干旱胁迫下野牛草雌雄株POD 活性变化趋势一致且没有显著性差异,说明野牛草雌雄株之间POD 抗氧化能力相似,这可能与POD 参与其他植物细胞活动有关[33]。干旱第7 天后,野牛草雌、雄株CAT 活性变化趋势完全相反,且在干旱第14 天野牛草雌株CAT 活性达到最低,说明野牛草雌株CAT 可能在干旱第14 天受到干旱胁迫影响导致活性降低,这与闻志彬等[34]发现CAT 活性在重度干旱胁迫下清除H2O2能力降低的结果一致。
野牛草雌株抗氧化酶基因表达量在复水后上调至峰值,与抗氧化酶活性变化并不一致,说明在野牛草雌株中抗氧化酶的合成可能同时受到其他基因的调控,但是其表达仍然受到干旱胁迫的诱导[35]。连玲等[14]对水稻抗氧化酶基因表达机制的研究结果表明,水稻抗氧化酶基因均受到干旱胁迫的诱导。野牛草雄株FeSOD、Zn/Cu-SOD、POD相对表达量在干旱第14 天达到峰值,与抗氧化酶活性变化相同,说明野牛草雄株上调FeSOD、Zn/Cu-SOD、POD相对表达量可以激活抗氧化酶的活性[36]。雄株APX以及CAT相对表达量在干旱第0 天最高后下降,复水后上升,抗氧化酶APX、CAT 活性在第14 天显著上升,表明APX 以及CAT 活性与其基因表达水平存在时间上的差异,原因可能在于其他未知因素参与控制了基因的转录过程,也可能是干旱胁迫导致转录后调控过程中酶降解或失活[37]。这与Xu 等[38]研究干旱胁迫下草地早熟禾(Poa pratensis)抗氧化酶活性和基因表达模式对干旱和干旱后恢复的响应研究结果一致。总的来说,野牛草雌、雄株之间抗氧化酶相关基因相对表达量变化趋势不同,表明不同性别野牛草植株抗氧化酶基因表达在响应干旱胁迫的过程中存在性别差异。
4 结论
综上所述,本研究从生理与转录水平探究了野牛草雌、雄株对干旱胁迫响应的差异。在干旱胁迫下,野牛草雌、雄株坪观质量、RWC 变化趋势一致,都呈先下降后上升的趋势。野牛草雌、雄株光合性能都受到干旱胁迫的抑制,两者在干旱-复水过程中光合性能差异不明显。除CAT 活性外,野牛草雌、雄株抗氧化酶活性在干旱第7天后都呈先上升后下降趋势。干旱第7 天后,野牛草雌、雄株CAT 活性变化趋势完全相反,野牛草雌株呈先下降后上升的趋势。通过基因表达分析发现,野牛草雌株抗氧化酶基因表达量与抗氧化酶活性变化并不一致;野牛草雄株FeSOD、Zn/Cu-SOD、POD相对表达量与相应抗氧化酶活性变化相同,APX以及CAT相对表达量与相应抗氧化酶活性变化不一致。综合来说,野牛草雌株抗旱性强于雄株,本研究发现了野牛草雌、雄株抗氧化系统在响应干旱胁迫中存在差异,为雌雄异株植物抗旱的差异性研究提供了参考。
猜你喜欢
食品安全导刊(2020年33期)2020-12-26
广西林业科学(2020年1期)2020-05-14
园林科技(2020年1期)2020-01-18
现代农业科技(2018年17期)2018-10-13
安徽农业科学(2018年12期)2018-05-03
山东农业科学(2017年4期)2017-05-18
江苏农业科学(2014年10期)2014-11-22
江苏农业科学(2014年9期)2014-11-15
现代农业科技(2009年7期)2009-07-29
现代农业科技(2009年20期)2009-03-20
