
天目山常绿落叶阔叶混交林细叶青冈种群结构与空间分布格局
2023-11-03 09:45宋思婧庞春梅余树全
浙江农林大学学报 2023年5期
宋思婧,丁 山,庞春梅,郑 枭,王 通,余树全
(1.浙江农林大学 林业与生物技术学院,浙江 杭州 311300;2.浙江天目山国家级自然保护区管理局,浙江 杭州 311311)
森林群落中优势树种的种群结构及空间分布是当前森林生态学的研究热点。目前,关于种群结构和空间分布的研究大多基于样地初次调查结果,对其动态监测的研究尚不多见,由于缺少长期样地监测数据,其结果存在一定局限性[1]。生态系统的长期监测能够为研究生态系统的演替过程提供可靠的资料[2],为发展和验证现代生态学理论提供平台,对生态系统动态理论的发展具有重要意义[3]。近年来,利用永久监测样地已经成为森林动态研究的主要手段[4]。
细叶青冈Cyclobalanopsisgracilis为壳斗科Fagaceae 青冈属Cyclobalanopsis常绿乔木,是中亚热带常绿落叶阔叶混交林主要建群种。木质坚韧,用途多样,具有较高的经济价值和生态效益,广泛分布于华东及湖北、湖南、广东、广西、贵州、四川、甘肃、陕西等省(自治区)。目前,关于细叶青冈的研究主要集中在生理特性、气候响应等方面[5-7],种群结构与空间分布格局的研究较少。本研究基于天目山国家级自然保护区典型常绿落叶阔叶混交林固定样地2012 和2017 年的调查数据,分析自然状态下常绿落叶阔叶混交林中细叶青冈种群结构和空间分布格局,探讨其动态规律及影响机制,旨在为该区域森林生态恢复、细叶青冈种群发展以及常绿落叶阔叶混交林的保护管理提供科学依据。
1 研究区概况
天目山国家级自然保护区位于浙江省杭州市临安区(30°18′30″~30°24'55″N,119°24′11″~119°28'21″E),主峰仙人顶海拔1 506 m[8]。保护区地处中亚热带北缘,属亚热带季风气候,四季分明,年平均气温8.8~14.8 ℃,年均降水量1 390.0~1 870.0 mm。保护区植被类型多样,植被分布存在明显的垂直带谱,沿海拔自下而上依次为常绿阔叶林(850.0 m 以下)、常绿落叶阔叶混交林(850.0~1 100.0 m)、落叶阔叶林(1 100.0~1 380.0 m)、落叶矮林(1 380.0 m 以上)[9]。
2 研究方法
2.1 样地设置与数据来源
为监测中亚热带森林生态系统的动态变化,1996 年在研究区选择典型常绿落叶阔叶混交林,建立了1 块1 hm2(100 m×100 m)的固定监测样地。样地地理位置位为30°20′22″N,119°25′47″E,海拔为1 066.2 m。将样地划分为25 个20 m×20 m 的小样方,对样地内胸径(DBH)≥10.0 cm 的木本植株挂牌编号,记录胸径、冠幅、树高、样方内坐标、生长状况等特征。考虑到小径级个体数量大且对生态过程较为敏感,因此在2012 年对样地进行复查时将起测胸径调整为1.0 cm,对样地木本植株进行每木检尺,对新增树木(DBH≥1.0 cm)进行挂牌编号,对死亡个体的状态进行记录。
采用2012 和2017 年2 期调查数据进行比较,分析细叶青冈种群结构与分布格局的动态特征。其中,2012 年样地内共记录到木本植物4 369 株,隶属44 科71 属121 种,平均树高为3.96 m,平均胸径为5.65 cm;2017 年样地内共记录到木本植物4 375 株,隶属45 科73 属113 种,平均树高为4.80 m,平均胸径为6.10 cm。样地中的主要树种有细叶青冈、交让木Daphniphyllummacropodum、大果山胡椒Linderapraecox、杉木Cunninghamialanceolata、中国绣球Hydrangeachinensi、青钱柳Cyclocarya paliurus、短尾柯Lithocarpusbrevicaudatus、缺萼枫香Liquidambaracalycina、微毛柃Euryahebeclados、宜昌荚蒾Viburnumerosum等。
2.2 龄级划分
采用径级结构代替年龄结构,根据样地调查数据的实际情况并参考相关文献[10],将细叶青冈胸径(DBH)划分为8 个等级:Ⅰ级(1.0≤DBH<2.5 cm)、Ⅱ级(2.5≤DBH<7.5 cm)、Ⅲ级(7.5≤DBH<12.5 cm)、Ⅳ级(12.5≤DBH<17.5 cm)、Ⅴ级(17.5≤DBH<22.5 cm)、Ⅵ级(22.5≤DBH<27.5 cm)、Ⅶ级(27.5≤DBH<32.5 cm)、Ⅷ级(DBH≥32.5 cm)。根据划分的径级结构将细叶青冈种群分为幼苗(Ⅰ级)、幼树(Ⅱ级)、中树(Ⅲ~Ⅴ级)、大树(Ⅵ~Ⅷ级)4 个生长阶段。
2.3 种群动态
根据新增与死亡株数,分析细叶青冈种群的死亡率[M=(lnN0-lnSt)/T]、更新率[R=(lnNt-lnSt)/T]和种群年增长率(λ=R-M)。其中:N0为某种群在第1 次调查时的个体数,Nt为该种群在第2 次调查时的个体数,St为该种群在第2 次调查时存活的个体数,T为2 次调查时间间隔。死亡个体是指复查时死亡或无法找到的植株;新增个体是指2017 年调查中新出现的胸径(DBH)≥1.0 cm 的个体(将DBH≥5.0 cm 的个体作为第1 次调查时的漏测个体,因而不计入新增个体)。
2.4 种群空间格局分析
2.4.1 种群不同生长阶段的空间分布格局分析 空间分布格局采用Ripley 提出的K(r)函数分析方法[11]:
其中:r为以目标树为圆点的取样圆半径;A为样地面积;n为样地树木株数;uij为点i和点j之间的距离,当uij≤r时,Ir(uij)=1;uij≥r时,Ir(uij)=0;wij为边界效应校正系数,是以点i为圆心,uij为半径的圆落在样地面积A中的比例,为一个点可以被观察到的概率。为了更直观地解释实际的空间格局,通常用Ripley’sL(r)函数表示:
其中:当L(r)=0 时,表示种群在空间尺度r内随机分布;当L(r)<0 时,表示种群在空间尺度r内均匀分布;当L(r)>0 时,表示种群在空间尺度r内聚集分布。
2.4.2 种群不同生长阶段的空间分布关联分析 空间分布关联基础函数计算如下:
其中:K12(r)表示径级1 和径级2 之间的K12(r)估计值,n1表示径级1 的植物个体数;n2表示径级2 的植物个体数。同样,用L12(r)取代K12(r),公式为:
其中:当L12(r)= 0、L12(r)<0 和L12(r)>0 时,分别表示径级1 与径级2 在空间尺度r下无相关、负相关、正相关。
本研究使用完全空间随机模型。采用Monte-Carlo 拟合检验,经过199 次重复拟合得到置信度为99%的置信区间,进行结果偏离随机状态显著性检验。若L(r)和L12(r)值在置信区间之内,表示种群呈随机分布或2 个径级之间无空间关联;在置信区间之上,表示种群呈聚集分布或2 个径级之间呈空间正相关;若在置信区间之下,表示种群呈均匀分布或2 个径级之间呈空间负相关。为减小边缘校正对数据的影响,将分析尺度限定在0~25 m,步长为1 m,数据分析过程通过生态学软件Programita[12]完成。
3 结果与分析
3.1 细叶青冈种群结构与动态
3.1.1 细叶青冈种群数量特征变化 2012 年样地内记录到细叶青冈共276 株,2017 年为291 株,较2012 年增加15 株,种群年增长率为1.06%,种群个体总数变化不大。5 a 间,样地内新增个体40 株,占2012 年细叶青冈种群总个体数的14.49%,死亡个体25 株,占2012 年细叶青冈种群总个体数的9.06%。细叶青冈种群平均胸径从10.65 cm 增加到10.82 cm,胸高断面积从4.71 m2·hm-2增加到5.15 m2·hm-2(表1)。
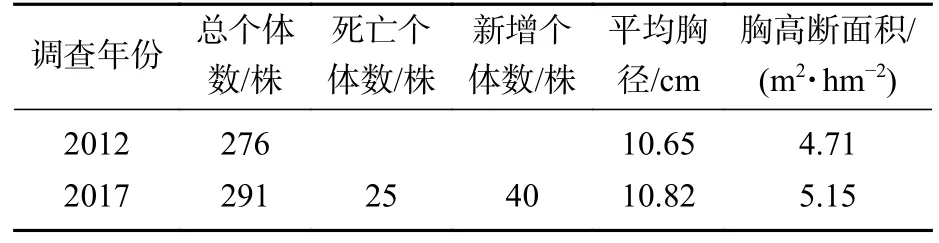
表1 细叶青冈种群数量特征Table 1 Quantitative characteristics of C.gracilis population
3.1.2 种群年龄结构 2 次调查中细叶青冈种群龄级结构均呈倒“J”型(图1),低龄个体占比较高,2012 和2017 年细叶青冈种群幼苗、幼树(Ⅰ~Ⅱ龄级)个体数分别占总个体数的53.99%和54.64%,随着龄级的增加,个体数量逐渐减少,表明种群具有较高的更新苗木储备,属于增长型种群。2017 年从Ⅰ龄级进入Ⅱ龄级的个体有20 株·hm-2,从Ⅱ龄级进入Ⅲ龄级的个体有11 株·hm-2,从Ⅲ龄级进入Ⅳ龄级的个体有8 株·hm-2,从Ⅳ龄级进入Ⅴ龄级的个体有7 株·hm-2,从Ⅴ龄级进入Ⅵ龄级的个体有6 株·hm-2,从Ⅵ龄级进入Ⅶ龄级的个体有7 株·hm-2,从Ⅶ龄级进入Ⅷ龄级的个体有4 株·hm-2,除Ⅰ、Ⅳ龄级个体由于生长进入下一龄级的数量较多,而达到径阶的新增个体不足,导致个体数下降外,其余龄级个体数均有增加。5 a 间,由于个体生长进阶补充,细叶青冈种群总个体数有一定变化,但龄级结构变化不显著,龄级结构保持随龄级升高,存活个体数减少的趋势,种群处于稳定增长的状态。
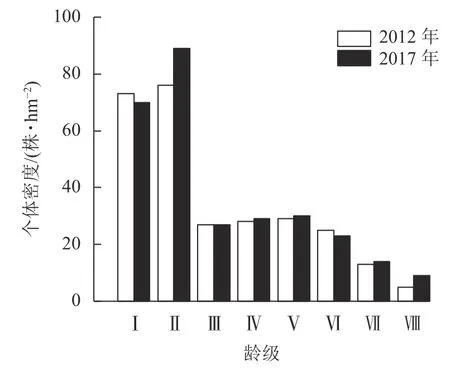
图1 细叶青冈种群龄级结构Figure 1 Age class structure of C.gracilis population
3.1.3 死亡与新增个体分布 种群年死亡率为1.90%,年更新率为2.96%。从不同龄级死亡个体的情况看,死亡与新增个体的龄级分布均呈倒“J”型,不同龄级死亡与新增个体数差异较大(图2)。死亡个体平均胸径为6.68 cm,集中在幼苗、幼树阶段(Ⅰ~Ⅱ龄级),死亡个体数为19 株,占死亡总个体数的76.00%,表明低龄级个体间竞争激烈,死亡率高;Ⅲ龄级死亡个体数为3 株,占死亡总个体数的12.00%,Ⅳ、Ⅴ龄级无死亡个体,Ⅵ、Ⅶ龄级死亡个体数分别为1 和2 株,仅占死亡个体数的4.00%和8.00%,说明高龄级个体死亡相对较少。新增个体平均胸径2.34 cm。Ⅰ龄级的新增个体数最多,为26 株,占新增个体总数的65.00%,Ⅱ龄级新增个体14 株,占新增个体总数的35.00%,Ⅲ~Ⅷ龄级无新增个体。总体来看,死亡与新增个体均集中分布在低龄级,且随龄级增加总体呈下降趋势。
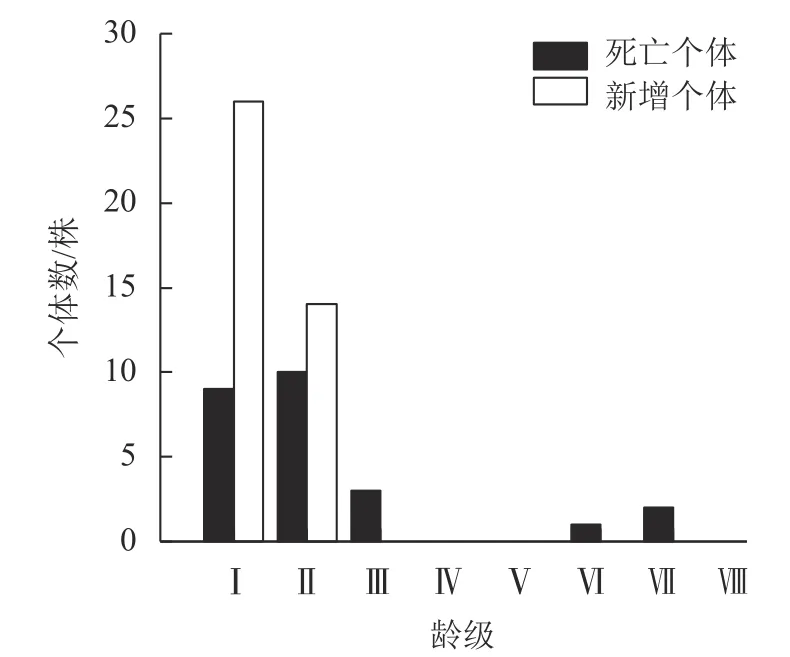
图2 细叶青冈种群新增及死亡个体龄级结构Figure 2 Age class structure of new and dead individuals of C.gracilis population
3.2 种群空间分布格局与空间关联性
3.2.1 种群空间分布格局 对天目山细叶青冈种群进行点格局分析发现(图3):不同生长阶段细叶青冈种群的空间分布格局有所差异。2012 年,细叶青冈种群整体在0~21 m 空间尺度内呈显著的聚集分布,在22~25 m 空间尺度内呈随机分布,其最高聚集强度为3.013,对应空间尺度为1 m;幼苗在0~18 m 空间尺度呈显著的聚集分布,在19~25 m 空间尺度呈随机分布,其最高聚集强度为4.425,对应空间尺度为3 m;幼树在0~11 m 空间尺度呈聚集分布,在12~16 m 空间尺度呈随机分布,随着空间尺度的增加在17~25 m 空间尺度又转为聚集分布,其最高聚集强度为3.621,对应空间尺度为1 m;中树在0~25 m 空间尺度均呈显著聚集分布,其最高聚集强度为4.231,对应空间尺度为23 m;大树仅在0~1、4~5 m 空间尺度呈聚集分布,2~25 m 空间尺度呈随机分布,其最高聚集强度为2.327,对应空间尺度为5 m。2017 年细叶青冈幼苗聚集程度较2012 年有所降低,在0~12 m 空间尺度呈聚集分布,在13~25 m 空间尺度呈随机分布;幼树聚集程度有所提高,在0~25 m 空间尺度均呈聚集分布;中树空间分布较2012 年小尺度范围聚集强度减弱,大尺度范围聚集强度增强,具体表现为在0~5 m 空间尺度呈聚集分布,在6 m 空间尺度呈随机分布,13~25 m 空间尺度呈聚集分布;大树随机分布强度较2012 年有所增强。总体来看,细叶青冈种群整体呈聚集分布,且随着龄级的增加逐渐转为随机分布。
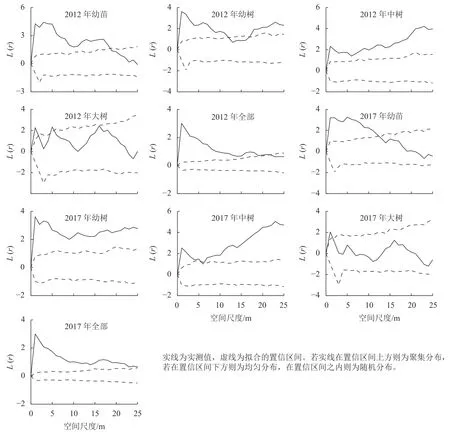
图3 不同生长阶段空间分布格局Figure 3 Spatial distribution pattern of different growth stages
3.2.2 种群不同生长阶段的空间关联 种群各生长阶段之间的空间关联能够直观体现种内关系的变化趋势,对了解不同林层之间的相互作用和种群稳定性变化具有重要意义[13-14]。对细叶青冈种群不同生长阶段的空间关联性进行分析发现(图4):2012 年,幼苗与幼树在1~11 m 空间尺度呈正相关,12~22 m 空间尺度为无相关,23~25 m 空间尺度为负相关;幼苗与中树在1~4 m 空间尺度为正相关,5~25 m 空间尺度为无相关;幼苗与大树在1~2、4~5 m 空间尺度为正相关,3、6~25 m 空间尺度为无相关;幼树与中树在1~10、21~25 m 空间尺度为正相关,11~20 m 空间尺度为无相关;幼树与大树在1~6 m 空间尺度为正相关,7~25 m 空间尺度为无相关;中树与大树在1~3 m 空间尺度为正相关,4~21 m 空间尺度为无相关,22~25 m 空间尺度为负相关。2017 年,细叶青冈种群各生长阶段两两空间关联趋势与2012 年保持一致,幼苗与幼树、幼苗与大树、幼树与中树、中树与大树空间正相关较2012 年有所增强。2017 年,幼苗与幼树在0~11、13~18 m 空间尺度呈显著正相关,幼苗与大树在0~8 m 空间尺度呈显著正相关,幼树与中树除了11、14~15 m 空间尺度外,在其他空间尺度均呈显著正相关。从整体来看,细叶青冈种群各生长阶段之间的空间关联均在小尺度呈正相关,随着尺度的增加过渡到无相关或负相关。
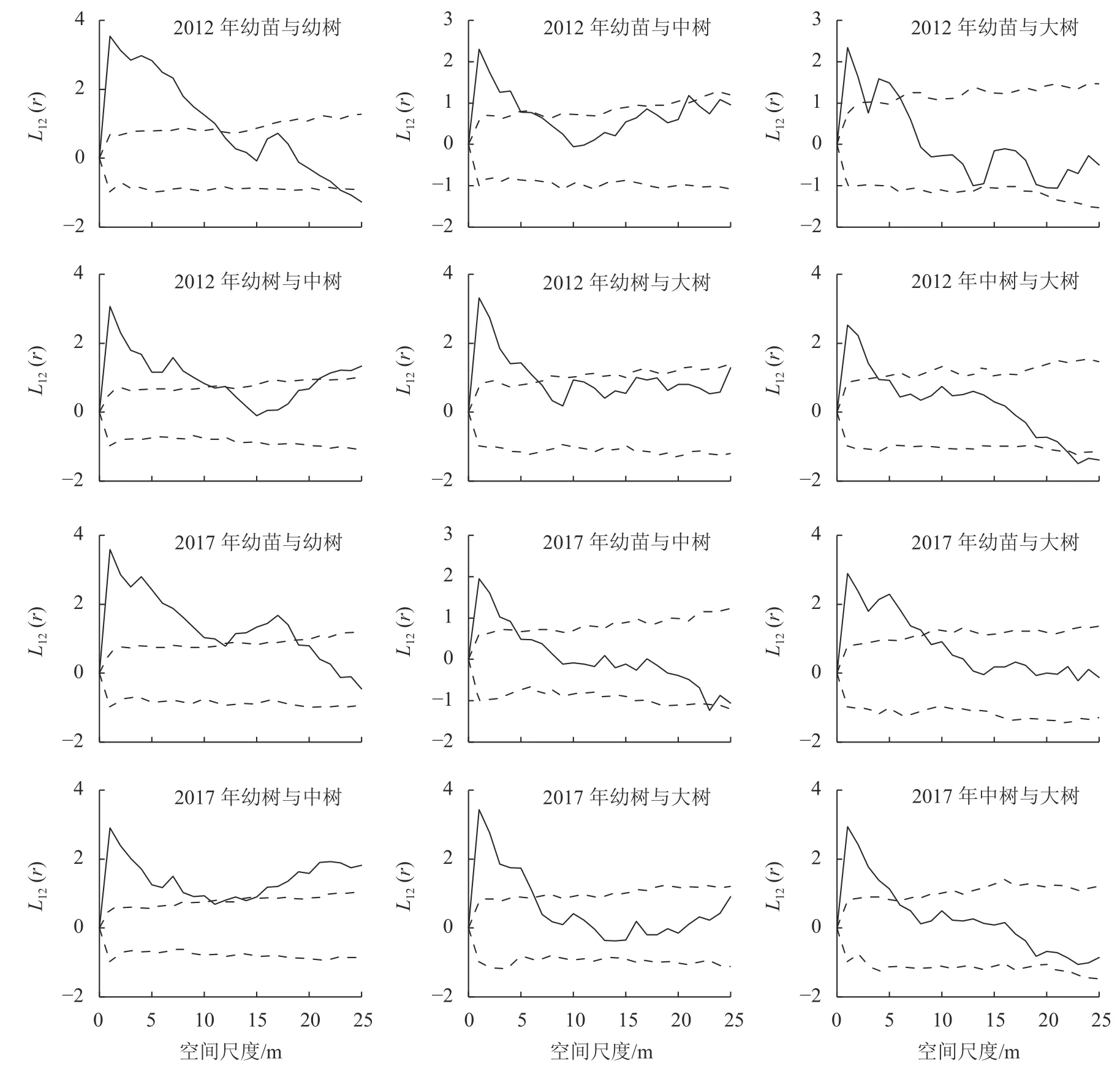
图4 不同生长阶段空间关联Figure 4 Spatial correlation of different growth stages
4 讨论
4.1 种群结构及动态特征
种群龄级结构能清晰地反映种群的生存状态,对预测种群动态具有重要意义[15]。由2 次调查结果可知:2012—2017 年细叶青冈种群总个体数上升,平均胸径增大。龄级结构无显著变化,均表现为倒“J”型。随着龄级的增大,个体数呈下降趋势,各径级无个体缺失,是典型的增长型种群。5 a 间细叶青冈种群死亡个体共25 株,年死亡率为1.90%,新增个体共40 株,年更新率为2.96%,新增个体的平胸径均较小,说明群落更新状况良好。种群年更新率高于死亡率,种群年增长率为1.06%,表明细叶青冈种群目前处于稳定发展状态,在未来短时间内个体数量不会发生较大幅度的变化。
种群个体的更新和死亡是种群结构变化的重要方式,体现种群种内竞争及其对环境的适应。细叶青冈种群不同龄级死亡个体数量差异明显,死亡个体多以小径级为主,这可能与低龄个体的生存竞争及环境筛选有关。细叶青冈虽为耐阴树种,但随着幼苗、幼树对光照、养分的需求不断增加,除了低龄个体间的相互竞争,还要与大树及其他树种个体争夺空间和养分,一定程度上阻碍了幼苗、幼树顺利进入林冠层[16];中树阶段由于个体处于生长旺盛期,在竞争中处于优势地位,同时因为达到林冠层,光照需求不再受到抑制,死亡率相对较小[17];进入生长末期后,细叶青冈接近生理死亡年龄,种群死亡率随之升高。
4.2 种群不同生长阶段空间分布格局
种群分布格局既受自身生物学特性和外部环境影响,同时与空间尺度密切相关[18-19]。通常在小尺度范围种群的空间分布主要受到种群自身生物学特性的影响,而在较大尺度上则更多受到生境异质性的影响[20]。本研究对细叶青冈种群各龄级的空间分布格局进行分析表明:2 次调查的空间分布格局类似,均在幼苗、幼树、中树阶段呈聚集分布,而在大树阶段呈随机分布,与蔡飞[21]对青冈Cyclobalanopsis glauca种群空间分布格局的研究结果一致,表明青冈属植物具有相似特性。这主要与细叶青冈自身的生物学特性及种群发育过程中因密度制约因素引起的种内种间竞争有关。一方面,由于根系萌蘖和种子的散布特点,细叶青冈幼苗基本聚集在母树周围,所以低龄阶段多呈聚集分布。另一方面,聚集分布能更好地改变微气候和小生境,有利于幼苗、幼树个体间的相互庇护,增强种群对不良环境的抗性[22],因此低龄个体常聚集分布。随着龄级的增大,密度制约引起种内竞争加剧,导致种群个体死亡增加,密度下降,故中树转变为大树后趋向随机分布。这种空间分布格局有利于将有限的环境资源的最大化利用,使植物个体满足自身的生存和发展,体现种群生长过程中对环境适应的生存策略及适应机制[23-24]。
4.3 种群不同生长阶段间的空间关联
同一物种不同生长阶段间的空间正关联来源于对环境的共同适应性,负关联来源于个体间的相互竞争排斥。细叶青冈种群各生长阶段均在小尺度范围内呈显著的正相关,随着尺度的增加过渡到无相关或负相关。种群空间格局与空间尺度的这种联系,可能与种内关系对空间尺度的依赖性有关[25]。同时,种群不同龄级间的空间关联性与龄级的差距有关。在小尺度范围内,种群个体间龄级差距越小,正关联性越强。细叶青冈幼苗与幼树的空间正相关大于幼苗与中树、大树,幼树与中树的空间正相关大于幼树与大树,说明幼苗与幼树对环境资源的选择具有一致性,所以在小尺度空间分布上表现为相互依存的关系。中树、大树可以为耐阴幼苗的生长提供较好的生存环境,因此幼苗与幼树、中树、大树在一定尺度范围内表现为空间正相关。然而这种空间关联可能会随着演替进程而发生变化,随着个体继续生长,细叶青冈个体抵御外界的胁迫能力增强,对空间资源及养分需求随之增加,导致竞争关系逐渐加强,使得中树与大树在大尺度表现为空间负相关。
5 结论
本研究基于2012 和2017 年天目山常绿落叶阔叶混交林1 hm2固定监测样地2 期调查资料,分析了细叶青冈种群结构和空间分布格局的动态变化。研究发现:天目山常绿落叶阔叶混交林中细叶青冈种群自然更新良好,处于稳定增长状态;2 次调查种群的空间分布格局类似,整体呈聚集分布,随着龄级的增加,种群的空间格局由聚集分布转为随机分布。各生长阶段间的空间关联在小尺度空间呈正关联,且在小尺度一定范围内,龄级差越小,正关联性越强。这是细叶青冈种子散布特性、种内种间竞争及环境等多种因素共同作用的结果,体现细叶青冈种群生长过程中的一种生存策略和适应机制。种群在生长过程中对环境干扰较为敏感,低龄个体因聚集分布受密度制约而竞争激烈,幼苗幼树阶段死亡率较高。为维持种群的长期稳定,必要时可以分阶段进行人工调控。5 a 间,种群中虽然有部分个体死亡与新增,但种群结构与空间分布格局没有发生显著变化,说明自然状态下细叶青冈种群结构与空间分布格局的变化是一个漫长的过程,研究其种群结构和空间分布格局动态需要更长的观测周期,对其进行持续性监测研究。同时,因为生态过程受多种因素影响,不仅与种群本身生物学特性有关,还可能与群落区域小生境、种间竞争、演替进程等因素有关,需要对这些方面进一步深入分析,以更好地掌握细叶青冈种群结构与空间分布的动态变化规律。
猜你喜欢
莫愁(2023年9期)2023-03-17
莫愁·家教与成才(2023年3期)2023-03-15
湖南生态科学学报(2022年4期)2022-11-11
东坡赤壁诗词(2020年3期)2020-07-04
河南农业大学学报(2020年2期)2020-05-22
山西林业科技(2019年1期)2019-05-27
西藏科技(2016年9期)2016-09-26
江苏农业科学(2016年6期)2016-07-25
广西林业科学(2016年3期)2016-03-16
东北林业大学学报(2014年8期)2014-08-02
