
基于表型性状构建桑树初级核心种质研究
2023-11-23 06:12万永辉阿不都赛买提艾买提阿巴白克扎克阿不都拉买提肉孜丁天龙
北方蚕业 2023年3期
万永辉 阿不都赛买提·艾买提 龚 明 阿巴白克·扎克 阿不都拉·买提肉孜 丁天龙
(新疆维吾尔自治区和田蚕桑科学研究所,新疆和田 848000)
新疆作为“丝绸之路”的重要节点,栽桑养蚕历史至少有1700多年,形成类型丰富、分布广泛、独具特色的桑树种质资源[1, 2]。种质资源是作物遗传改良和相关基础研究的物质基础。拥有作物种质资源的数量和质量,以及其研究和利用的深度和广度,直接影响到现代种业的可持续发展,种质资源保护和利用已日益成为世界各国驱动农业科技创新的重要部分[3]。根据FAO(1996)的统计资料世界范围内作物种质资源收集品已达740万份[4]。种质资源数量及资源库容量的不断增大给保存、评价、研究和利用带来了众多困难[5]。Frankel首次提出核心库(Core Collection)概念,即能代表一个作物种的遗传多样性,且具有最小样品重复的物种子集[6],此后核心种质的概念不断得到完善和发展。核心种质概念的提出使种质资源的研究有了方向和重点,为种质资源的利用创造了基础[7]。
本研究以新疆维吾尔自治区和田蚕桑科学研究所种质资源圃内收集保存的652份桑种质资源为试验材料,基于表型数据进行遗传多样性分析和核心种质的构建,并对不同方法构建的核心种质代表性进行评价,最终确定选出核心种质,旨在为新疆桑树种质资源收集保存、评价鉴定及创新利用提供依据。
1 材料与方法
1.1 试验材料
从新疆维吾尔自治区和田蚕桑科学研所桑树种质资源圃中挑选652份桑种质作为初级核心种质构建的原始样品集,来自国内种质资源603份,其中新疆432份、四川58份、浙江43份、山东17份、黑龙江14份、陕西8份、贵州7份、河北5份、江苏5份、湖北4份、重庆3份、安徽2份、吉林2份、山西2份、广西1份;来自国外种质资源49份,其中乌兹别克30份、日本16份、朝鲜3份。种质资源圃种植株距为1 m,行距为1.5 m,树龄为8 a。
1.2 方法
1.2.1 数据调查与整理
参照《农作物种质资源鉴定技术规程 桑树》[8]调查652份桑种质11个表型性状,其中种质类型、花性为质量性状;发芽期采取按天调查的方式,转化成质量性状;一年生枝条长度难以精确测量,按照枝条的长短分为3级,转化成质量性状处理;质量性状的标准化见表1。叶长、叶幅、节间长度、发芽率、生长芽率、春季米条叶产量和秋季米条叶产量为数量性状,分别用直尺和天平测定。

表1 桑种质质量性状赋值
1.2.2 遗传多样性分析
遗传多样性指数H'=-∑(Pi)(lnPi),式中Pi为某个性状中第i个级别的材料占总样本的百分比。参照芮文婧等[9]、郑福顺等[10]和王振江等[11]的方法,基于平均值和标准差对所有种质每个数量性状划分等级,每0.5σ(标准差)为一个等级,共划分成10级。
1.2.3 数量性状相关性分析
选取枝条长度、叶长、叶幅、节间长度、发芽率、生长芽率、春季米条叶产量和秋季米条叶产量为数量性状8个性状进行相关性分析。
1.2.4 表型性状主成分分析
对652份桑树种质资源的种质类型、花性、发芽期、节间长度、叶长、叶幅、枝条长度、发芽率、生长芽率、春季米条产叶量和秋季米条产叶量11个性状进行主成分分析。
1.2.5 核心种质构建
利用QGA Station 2.0软件,采用欧氏距离计算遗传距离,利用不加权类平均法(unweighted pair-group method with arithmetic means,UPGMA)进行聚类,并分别选用随机取样法、优先取样法和偏离度取样法进行取样,取样比例为25%,以构建核心种质。
1.2.6 核心种质代表性评价
参考章秋平等[12]和刘遵春[13]等的方法,构建的核心种质须同时符合2个条件:1)均值同原始种质资源群体存在显著差异(α=0.05)的性状占全部性状的百分数小于或等于20%;2)核心种质和原始种质资源群体的极差符合率不低于80%。利用极差符合率、变异系数变化率、均值差异百分率、方差差异百分率对核心种质的代表性进行评价。利用均值t检验对原群体和核心种质表型性状均值间是否有显著性差异进行检验,利用方差F检验分析两者表型性状变异的同质性。
1.3 数据分析
使用Excel 2016进行数据整理,使用SAS 9.4进行相关性分析、主成分分析和聚类分析,使用Origin 2022绘制聚类图,使用QGA Station 2.0抽取核心种质。
2 结果与分析
2.1 桑树种质遗传多样性分析
2.1.1 质量性状遗传多样性分析
652份桑树种质资源4个质量性状遗传多样性见表2。4个质量性状共计14个变异类型,各类型表现出不同的分布频率。遗传多样性指数分布在0.71~1.18之间。花性遗传多样性指数(H')最小为0.71,雌花分布频率最大为75.61%,雌雄同株分布频率最小为8.13%。发芽期遗传多样性指数(H')最大为1.18,4月11日~13日发芽的种质数量最多,分布频率为56.14%,4月17日~19日发芽的种质数量最小,分布频率为2.91%。种质类型遗传多样性指数为0.82,以野生型为主,分布频率为65.34%,其次为栽培品种,分布频率为27.91%,品系数量最小,分布频率为6.75%。枝条长度遗传多样性指数为0.77,以长枝条类型为主分布频率为70.86%,短枝条类型数量最小,为7.82%。
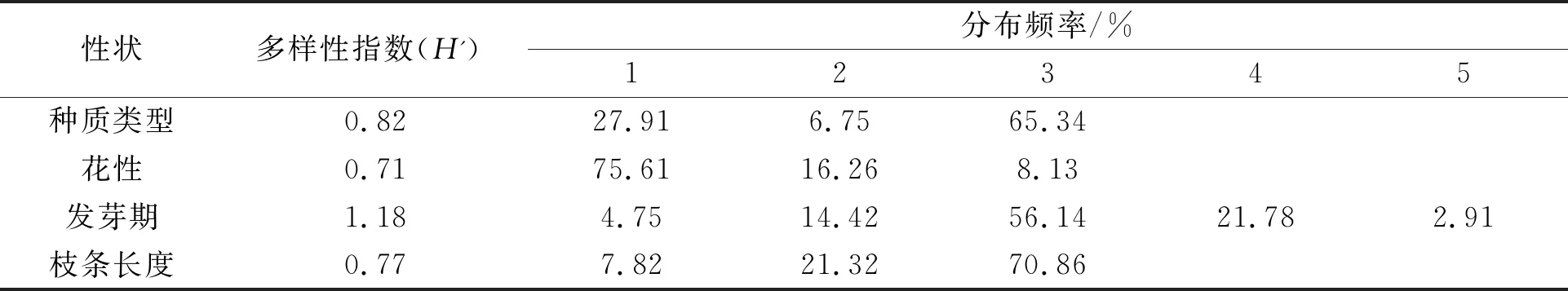
表2 桑种质质量性状遗传多样性分析
2.1.2 数量性状遗传多样性分析
652个桑树种质数量性状遗传多样性见表3。7个数量性状变异系数在16.36%~53.49%之间,其中发芽率变异系数最小为16.36%,秋季米条产叶量变异系数最大,达到53.49%。7个数量性状遗传多样性指数(H')在1.82~2.03之间,其中秋季米条产叶量遗传多样性指数(H')最小,为1.82,叶长遗传多样性指数(H')最大,达到2.03。由此可见,供试的 652个桑树种质材料具有丰富的遗传多样性,是重要的种质资源。

表3 652份桑种质资源数量性状遗传多样性分析
2.1.3 数量相关性分析
652份桑树种质材料9个数量性状相关分析见表4。发芽期与叶长、叶幅、春季米条产叶量、秋季米条产叶量呈极显著正相关,与生长芽率极显著负相关。节距与叶长、叶幅、枝条长度、秋季米条产叶量成极显著正相关,与生长芽率成显著负相关。叶长与叶幅、春季米条产叶量和秋季米条产叶量成极显著正相关,与发芽率和生长芽率呈极显著负相关。叶幅与春季米条产叶量和秋季米条产叶量呈极显著正相关,与发芽率和生长芽率成极显著负相关。枝条长度与发芽率和生长芽率成显著负相关。发芽率与生长芽率成极显著正相关,与秋季米条产叶量呈极显著负相关。生长芽率与秋季米条产叶量呈极显著负相关。春季米条产叶量和秋季米条产叶量呈极限显著正相关。

表4 652份桑树种质数量性状相关性分析
2.1.4 表型性状的主成分分析
652份桑树种质材料11个性状主成分分析结果见表5。前4个主成分特征值大于1,方差贡献率达69.103%,包含了11个性状的绝大部分信息,表明这4个主成分能代表11个表型性状的遗传信息。其中,第一主成分特征值3.528,方差贡献率为32.069%,主要代表为叶长、叶幅,特征向量值在0.8以上,这一主成分主要与桑树叶片大小有关。第二主成分特征值为1.693,方差贡献率为15.395%,主要代表为发芽率和生长芽率,特征向量值均在0.7以上,这一主成分主要与桑树物候特征有关。第三主成分特征值1.246,方差贡献率为12.325%,主要代表为节距和枝条长度,特征向量值均超过0.7主要与枝条形态相关。第四主成分特征值为1.135,方差贡献率为10.314%,主要代表为花性,特征向量值为0.766。

表5 652份桑树种质资源表型性状主成分分析
2.1.5 桑树种质表型形状聚类分析
使用SAS 9.4软件中的CLUSTER过程进行层次聚类分析,在分类数为7的条件下CCC值(立方聚类条件值)达到峰值为-14,伪F统计量达到峰值为346,伪t方值达到最小值为29.6,R方值等于0.763,半偏R方值等于0.0063。因此,将652份桑种质划分成7类,此时聚类之间的平均距离为0.730。聚类结果见图1。
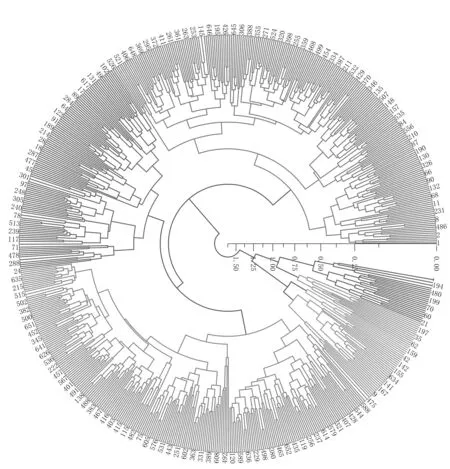
图1 652份桑种质资源聚类分析
第I类包含229个种质,主要表现为发芽期、节间长度、叶长、叶幅、条长、发芽率、生长芽率、春季米条产叶量和秋季米条产叶量与群体平均值相近。
第II类包含119个种质,主要表现为发芽期、节间长度、叶长、叶幅、条长与群体平均值相近,发芽率和生长芽率较低,春季米条产叶量较低,秋季米条产叶量较高,秋季米条产叶量高于春季米条产叶量。
第III类包含234个种质,主要表现为发芽期较早,节长度较短,叶长和叶幅较短,条长与群体平均值相近,发芽率和生长芽率高于群体平均值,春季米条产叶量和秋季米条产叶量均较低。
第IV类包含26个种质,表现为发芽期较晚,节间长度较长,叶长和叶幅较大,枝条长度和群体平均值相近,发芽率和生长芽率与群体平均值相近,春季米条产叶量较高,秋季米条产叶量与群体平均值相近。
第V类包含7个种质,均开雌花,表现为发芽期较晚,节间长度较长,叶长和叶幅较大,枝条长度较长,发芽率和生长芽率与群体平均值接近,春季米条产叶量在7个群体中最高,秋季米条产业量与群体平均值相近。
第VI类包含13个种质,均为栽培种和品系,表现为发芽期较晚,节间长度与群体平均值相近,叶长和叶幅较大,枝条长度较小,发芽率和生长芽率较低,春季米条产叶量和秋季米条产业量均较高。
第VII类包含24个种质,表现为发芽期较晚,节间长度较大,叶长和叶幅较大,枝条长度较小,发芽率和生长芽率较低,春季米条产叶量和秋季米条产业量均高于群体平均值,春季米条产叶量和秋季米条产叶量相近。
2.2 核心种质的构建与评价
使用QGA Station软件,采用欧式距离计算遗传距离,利用不加权平均数法(UPGMA)进行聚类分析,取样比例设为25%,分别采用随机取样法、优先取样法和偏离取样法取样,[14-17]构建3组核心种质R1、P1和D1,各核心种质材料信息见表6。在原群体中栽培种为182份,占群体27.91%;品系为44份,占群体的6.75%;野生种为426份,占群体的65.34%。
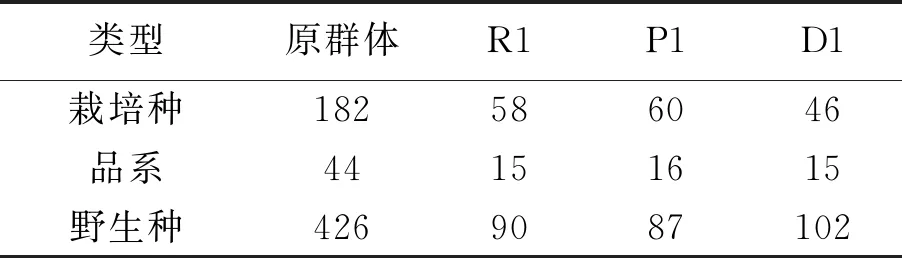
表6 原群体与各核心种质材料信息表
在R1核心种质中栽培种为58份,占群体的35.58%;品系为15份,占群体的9.20%;野生种为90份,占群体的55.21%。在P1核心种质中栽培种为60份,占群体36.81%;品系为16份,占群体的9.82%;野生种为87份,占群体的53.37%。在D1核心种质中栽培种为46份,占群体28.22%;品系为15份,占群体的9.20%;野生种为102份,占群体的62.58%。
核心种质与原群体差异百分率见表7。与原群体相比,核心种质R1和P1均值差异百分率均大于20%,分别为54.55%和45.45%,D1均值差异百分率为18.18%,小于20%。三者极差符合率均大80%,分别为98.92%、100.00%和99.37%。表明D1符合构建核心种质的原则,具有原种质资源的代表性,能够较好地反映出原种质群体的遗传多样性。
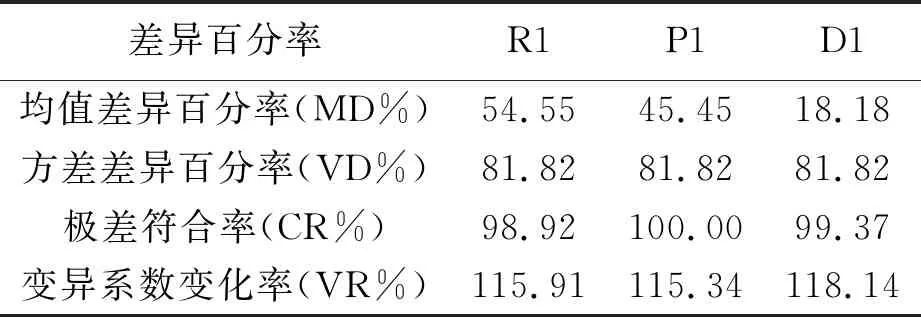
表7 核心种质与原群体差异百分率
利用t检验检验核心种质与原群体表型性状均值之间是否存在显著性差异,利用方差F检验分析两者表型性状变异是否同质。构建的3组核心种质与原群体的均值、方差差异比较见表8。R1有2个指标与均值与原群体存在显著性差异,有4个指标均值与原群体存在极显著性差异。P1有3个指标均值与原群体存在显著性差异,有2个指标均值与原群体存在极限性差异。D1只有2个指标均值与原群体存在极显著性差异。表明R1和P1获得了更大的变异性,而D1变异较小。除秋季米条产叶量外,R1、P1和D1的变异系数均大于原群体,表明所构建的3组核心种质具有良好的异质性。

表8 核心种质与原群体的差异性比较
3 讨 论
我国幅员辽阔,地势复杂,生态各异,栽桑历史悠久,在长期自然和人工选择下形成极为丰富的种质资源[18,19]。据不完全统计,全国保存的各类桑树种质资源超过3000份[20],保存桑种质资源数量居世界之首[21]。开展桑树种质资源遗传多样性研究,对种质亲缘关系鉴定、保存、利用、核心种质的构建、评价及桑树育种都具有重要的实用价值和理论意义[22]。王振江等[11]基于 8 个农艺性状对 569 份桑种质进行了遗传多样性分析,将569 份桑树种质资源进划分为 5 个类群,不同类群间则具有明显差异。本研究基于表型指标采用相关性分析、主成分分析和聚类分析对652份桑种质遗传多样性进行研究,结果表明652份桑种质具有丰富的遗传多样性,可以作为遗传研究的试验材料。
丰富的遗传资源为遗传研究及育种工作提供了大量的材料,然而,如此众多的资源给保存、评价、鉴定及创新利用带来了困难,构建核心种质是解决这一问题的有效措施之一[23]。曾钦朦等[24]以245份贵州核桃种质资源为试材,基于13个数量性状数据,采用随机取样策略、偏离度取样策略和位点优先取样策略,结合8个取样比例(5%、15%、20%、25%、25%、30%、40%、50%),进行多次聚类构建核心种质。结果表明,取样比例设置为50%,采用偏离度取样策略构建的核心种质最具代表性。刘闯萍等[25]利用156份山葡萄种质资源采用采集地分组-花型分组-聚类-分组法构建核心种质,核心种质包含48份材料,较好地代表了总体样本的遗传变异。章秋平等[12]以447份普通杏为供试材料,基于40个农艺性状,设置取样比例为25%,采用逐步聚类随机取样法、最小距离逐步取样法和优化LDSS法构建核心种质,结果显示优化后的ILDSS法构建出的核心种质最具代表性。刘娟等[26]以135份新疆野杏株系为试材,根据35个表型性状的遗传多样性,采用逐步聚类法,以25%的取样比例为例,通过3种取样策略、2种遗传距离和4种系统聚类方法相互组合构建了24个核心种质,并对核心种质的代表性进行了评价。结果表明,取样比例为25%时,采用优先取样策略,使用欧氏距离和离差平方和法进行逐步聚类构建的核心种质最有代表性,是构建新疆野杏核心种质的较好方法。本研究样本容量为652份,根据前人的研究经验将取样比例设置为25%,利用不加权类平均法(unweighted pair-group method with arithmetic means,UPGMA)进行聚类,并分别选用随机取样法、优先取样法和偏离度取样法进行取样,成功构建出3组核心种质。
利用表型数据构建核心种质是一种较为成熟的方法,可以极大地减轻工作量。但表型性状较易受环境因素的影响,无法准确地表现出基因水平上的差异。目前,利用分子标记技术构建核心种质是研究热点,如利用SSR、ISSR、SNP等分子标记方法成功构建了山核桃[27]、新疆野杏[28]、荔枝[29]等园林作物核心种质。我们下一步拟利用分子标记与农艺性状相结合构建桑核心种质,为特异种质挖掘和品种改良提供参考依据。
4 结 论
本研究以652份桑树种质为供试材料,通过相关性分析、主成分分析和聚类分析,发现该 652份桑种质资源具有丰富的遗传多样性。以 25%为取样比例,分别采用随机取样法、优先取样法和偏离取样法进行取样,成功构建出3组核心种质R1、P1和D1,其中D1符合构建核心种质的原则,具有原种质资源的代表性,能够较好地反映出原种质群体的遗传多样性。
猜你喜欢
今日农业(2022年13期)2022-09-15
音乐教育与创作(2022年1期)2022-04-26
今日农业(2020年16期)2020-12-14
种子科技(2018年11期)2018-09-10
许昌学院学报(2018年8期)2018-09-05
小学生作文(中高年级适用)(2018年3期)2018-04-18
中国麻业科学(2018年6期)2018-04-09
学生天地(2016年16期)2016-05-17
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
