
Expression of VEGF165 and VEGF165b during ovarian follicular development
2023-12-12 02:55ChinnaratChangsangfaBongkochTurathumMorakotSroyrayaKhwanthanaGrataitong
Chinnarat Changsangfa ,Bongkoch Turathum ,Morakot Sroyraya ,Khwanthana Grataitong
1Institutes of Molecular Biosciences,Mahidol University,Nakhon Pathom,73170,Thailand
2Department of Basic Medical Science,Faculty of Medicine Vajira Hospital,Navamindradhiraj University,Bangkok,10300,Thailand
3Department of Anatomy,Faculty of Science,Mahidol University,Rama 6 Road,Bangkok,Thailand
ABSTRACT Objective:To investigate the role of vascular endothelial growth factor (VEGF)165a,VEGF165b,and VEGF receptor (VEGFR) in the development of bovine follicles.Methods:We cultured follicular cells that were collected from small,medium,and large sized bovine follicles with estrogen and measured the expression of VEGF,VEGFR2 and VEGF165b by Western blot analysis and immunofluorescence.Results:The expression of VEGF165 increased in all follicle sizes and the expression of VEGF165b was increased in the small and large follicles after culturing in an estrogen containing medium.The expression of VEGFR2 was increased in the medium and large follicles after culturing with estrogen for 96 h.VEGF165 was activated at 100 ng/mL estrogen in the large follicles for 96 h.In addition,VEGFR2 was upregulated in the medium and large follicles after treated with 100 ng/mL estrogen for 96 h.Conclusions:This evidence suggests that the expression of VEGF165 and VEGFR is associated with estrogen stimulation during the development of bovine follicles and in an autocrine or paracrine manner.This reveals an advantage during oocyte maturation in vitro.
KEYWORDS: Ovarian follicles;Vascular endothelial growth factor;VEGF;VEGFR2;VEGF165b;Estrogen stimulation
1.Introduction
The ovarian follicle is an important structure that supplies the necessary factors for oocyte growth and maturation[1,2].During ovarian follicular development,angiogenesis plays an important role by increasing blood supply to antral follicles.Vascular epithelial growth factor (VEGF;also referred to as VEGFA) is a major regulator of angiogenesis in the ovary[3,4].VEGF and its receptors (VEGFR) comprise a family of tyrosine-kinase-type receptors,e.g.,Flt-1 (VEGFR1),KDR/Flk-1 (VEGFR2),and Flt-4 (VEGFR3),and are presented in many cell types and have been detected in mammalian ovaries[5,6].
In the female reproductive system,VEGF is necessary for follicular and corpus luteum development and is a biomarker of oocyte maturation[7,8].It has been found that VEGF promotes the transition of primary to secondary follicles by increasing vascularity leading to an increased delivery of intraovarian and systemic factors to the primary follicles in cattle[9].VEGFA is composed of many isoforms (e.g.,VEGF121,VEGF145,VEGF165,VEGF189 and VEGF206) created by the alternative splicing of VEGF mRNA[10].Alternative splice site selection in exon 8 generates 2 families of isoforms,the proangiogenic (e.g.VEGF165) and antiangiogenic family (e.g.VEGF165b) which have different functions[11,12].Previous studies have been shown that VEGF165 expression increased in the follicles during development leading to an increase in the rate of antral formation,stimulated meiosis resumption,and activated the production of mature oocytes in goats[13,14].When the intraovarian VEGF/VEGFR2 pathway is blocked,the secretion of follicular hormones is altered,indicating that the intraovarian effect of VEGF is mediated by this receptor in monkeys[15].VEGFR2 is expressed in the oocytes of caprine ovarian follicles at all developmental stages and in the granulosa cells of developing follicles[16].As VEGFA is an important role for ovarian follicular development,VEGF165b is localized within the different cell types in the marmoset ovary and levels are decreased in the corpus luteum of the ovary.It has been shown that over-expression of VEGF165b in the mouse ovary reduced the secondary and tertiary follicles after mating and increased atretic follicles[17].The regulation of follicular development,meiosis resumption,and oocyte maturation are controlled by the differential splicing of VEGF.However,the mechanism of VEGF to switch from VEGF165b to VGF165 during ovarian follicular development in bovine has not been studied.
Significance
Angiogenesis and vascular regression are essential for the growth of ovarian follicles and the ovulatory cycle which are modulated by estrogen.This study revealed that the expression of VEGF165 and VEGF receptors are involved in an autocrine and paracrine manner in the stimulation of estrogen during the development of the bovine follicles.It demonstrated the benefit during in vitro oocyte maturation.
The overall objective of this study was to study the expression and mechanism of VEGF165,VEGF165b,and its receptor in follicular development in cattle.
2.Materials and methods
2.1.Collection of ovaries
Ovaries were obtained from healthy adult female bovine (>15 months old) from a slaughterhouse.Ovaries were placed in 0.9% NaCl saline (containing 100 IU/mL penicillin) and transported on ice to the laboratory.Ovaries were washed 3 times with phosphate buffer saline (PBS) and the ovary cortex was cut into small pieces.The ovaries were fixed in 4% paraformaldehyde and embedded in paraffin blocks.Sections were cut at 5 m thickness and subjected to immunohistochemistry/immuno-fluorescence (VEGFR1/VEGFR2).For primary cell culture,follicles with a diameter greater than 1.0 cm were classified as large follicles,those with a diameter of between 0.5 and 1.0 cm were considered to be medium sized,and follicles with a diameter of less than 0.5 cm were characterized as small.
2.2.Immunofluorescence
After fixation for 12 h,the follicles were dehydrated in a graded series of ethanol,cleared in xylene,and embedded in paraffin wax using conventional procedures.Serial sections of 5 µm thickness were cut with a microtome and processed for immunofluorescence.Immunofluorescence demonstration of VEGF and VEGFR2 was performed.To expose the antigenic sites for VEGF,dewaxed sections were heated four times to 95 ℃ with 1% glycine in a 600 W microwave oven for 15 min and allowed to cool for 20 min.The sections were then treated with 3% bovine albumin serum (BSA) diluted in PBS with Tween 20 (PBST) for 2 h.To reduce nonspecific background staining,sections were treated with polyclonal anti-VEGF antibody 1:300 in PBST and incubated overnight at 4 ℃.For VEGFR2,the primary antibody was antirabbit VEGFR2 (1:50;Cell Signaling).
2.3.Primary granulosa cell culture
To determine the effect of estrogen stimulation on the total VEGF,VEGF165b,and VEGFR2 expression,in vitro experiments were performed on freshly isolated bovine granulosa cells.Briefly,ovaries collected from the slaughterhouse were rinsed in 70% ethyl hydroxide.Granulosa cells were isolated from small,medium,and large follicles and plated in DMEM/F12 supplemented with 10% fetal bovine serum (FBS) and 2% penicillin/streptomycin (Gibco BRL,Burlington,ON,Canada).At 70% confluence (approximately 24 h),cells were switched to serum-free DMEM/F12 (supplemented with 2% penicillin/ streptomycin) in the presence or absence of various concentrations of estrogen (Sigma,Oakville,ON: 1,10,and 100 ng/mL luteinizing hormone) for 0,24,and 48 h.After treatment,granulosa cells were lysed and total protein was isolated for VEGFR2,VEFG165b,and VEGF and were detected by western blot analysis.
2.4.Protein extraction
Protein extraction from mouse tissues was conducted via liquid nitrogen homogenization followed by radioimmunoprecipitation (RIPA) buffer resuspension (5 µL/µg).Protein concentration was determined by Bradford Assay (BioRad) and 100 µg of protein denatured for polyacrylamide gel electrophoresis (PAGE) with laemmli buffer and 5% β-mercaptoethanol for 5 min at 100 ℃.
2.5.Western blotting (VEGF)
Protein extracted from primary granulosa cells was denatured and 20 µg was run on sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE).Recombinant human VEGF165a and VEGF165b (R&D Systems) were run as positive controls (50 ng each).Proteins were separated using 4%–20% precast gels (456–1094,BioRad) run for 2 h at 90 V and transferred to nitrocellulose membrane (15V,10 min,170–4150,BioRad).Membranes were blocked overnight in 2.5% (w/v) BSA in 0.1% (v/v) Tris-buffered saline with Tween20 (TBST) at 4 ℃ and co-immunoblotted with VEGFA antibody (0.2 µg/mL,AF293NA,R&D) and VEGF165b specific antibody (1.2 µg/mL,MAB3045,R&D) in 2.5% BSA/TBST.Blots were washed 5 times with 0.1% TBST for 5 min and co-immunoblotted with antimouse and antigoat fluorescent antibodies (926–32212 and 926–68074,respectively;1: 10 000;Licor Biosciences) for 1 h at room temperature.The blots were washed and imaged using the Odyssey®Fc imaging system (Licor Biosciences).
2.6.Statistical analysis
The data were analysed using GraphPad PRISM 9 software (GraphPad Software Version 9,La Jolla California USA) for data manipulation,calculation,and graphical display.The protein expression level was expressed as mean±standard deviation (mean±SD).The independent sample two-way ANOVA was used to assess statistical significance,and P<0.05 was considered statistically significant.
2.7.Ethics statement
The study was approved by Institutional review board,Faculty of Medicine Vajira Hospital (COE_NOT HS: 15/2019).The ovaries were processed following ethical standards,and were obtained from dead animals.
3.Results
3.1.Expression of VEGF and VEGFR2 from different sized follicles of bovine ovaries
Using immunofluorescence,we found that the expression of VEGF was higher in large follicles compared to the medium and small follicles and the expression of VEGF was increased in the corpus luteum (Figure 1).The expression of VEGFR2 was higher in the large follicles compared to the medium and small follicles and the expression of VEGFR2 was also increased in corpus luteum (Figure 2).These data demonstrated that the expression level of VEGF was increased during follicle development and related to the expression of VEGFR2.

Figure 1.Expression of ascular epithelial growth factor (VEGF) in the bovine ovarian follicle and corpus luteum by immunofluorescence.The intense green fluorescent signals are observed in ovarian follicle and corpus luteum using an anti-VEGF specific antibody.Bars,50 µm.
3.2.Expression of VEGF,VEGF165a,and VEGF165b during ovarian follicular development when treated with estrogen
To determine the effect of estrogen on VEGF165,VEGF165b,and its receptor (VEGFR2) during ovarian follicular development,we cultured follicular cells collected from small,medium,and large sized bovine follicles with estrogen and measured the VEGF165b and VEGF165 expression by western blot and immunofluorescence (Figure 3).The results showed that the expression of VEGF165 was significantly decreased in the small sized follicles after cultured with 100 ng/mL estrogen for 48 h compared to the control group.After 4 days,the expression of VEGF165 in the small follicles was significantly increased compared to other estrogen treated groups (100 ng/mL).The expression of VEGF165 in medium follicles was not different between the control and treated groups after culturing for 48 h,but the expression of VEGF165 was significantly increased when cultured with 100 ng/mL estrogen after 96 h.In the large follicle,the expression of VEGF165 was not different between the four groups after culturing for 48 and 96 h,but it was significantly increased between the 10 and 100 ng/mL groups when compared at 48 and 96 h.This data demonstrated that the level of VEGF165 (proangiogenic factor) was higher after treated with 100 ng/mL estrogen in the small and large follicles when treated with 10 and 100 ng/mL estrogen after culturing for 96 h.
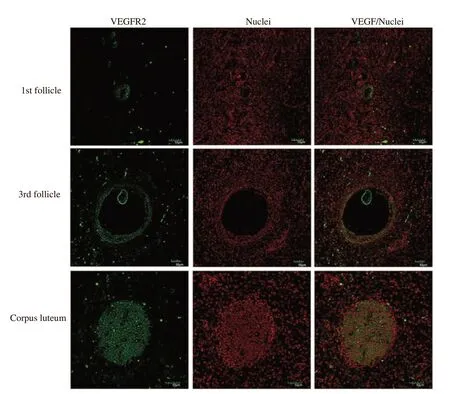
Figure 2.Expression of VEEGFR2 in the bovine ovarian follicle and corpus luteum by immunofluorescence.The intense green fluorescent signals are observed in ovarian follicle and corpus luteum using an anti VEGFR2 specific antibody.Bars,50 µm.
The expression of VEGF165b (antiangiogenic factor) in small follicles were not significantly different between the control and 1 and 10 ng/mL estrogen groups after culturing for 48 and 96 h but was significantly increased when treated with 100 ng/mL for 96 h.In the medium follicles,the expression of VEGF165b was significantly increased after treated with 1 and 10 ng/mL estrogen after cultured for 48 h.In the large follicles,the expression of VEGF165b was significantly increased after treated with 10 and 100 ng/mL estrogen.The expression of VEGF165b was increased in the follicular cells of small and large follicles after culturing in a medium containing various concentrations of estrogen.The expression of VEGFR2 was increased in all medium and large follicles after they were cultured with estrogen for 96 h.
4.Discussion
VEGF expression during follicular development depends on follicular size.VEGF plays an important role in follicular growth.This evidence indicates that the interaction of VEGF and its receptor in granulosa cells acts as a mitogenic factor in the follicular development[18].Additionally,previous reports have indicated that VEGF inhibits the follicular apoptosis in cows and heifers[19] and also modulates the effects of gonadotropins in bovine granulosa cells[20].Moreover,the reduction of follicle angiogenesis and lack of development of the mature antral follicles results from the inhibition of VEGF[16].In the present study,we observed a correlation between the size of the follicle and VEGF expression in bovine folliculogenesis.Our results demonstrated that the expression level of VEGF is upregulated in the large sized follicle compared to the medium and small follicles and was associated with the level of VEGFR2.This evidence suggests that VEGF and its receptor play a critical role in follicular development.

Figure 3.Western-blot analysis and their quantification of total VEGF,VEGF165b,VEGF165 and VEGFR2 from small (A),medium (B) and large follicles (C) treated with estrogen [1 ng/mL,10 ng/mL,and 100 ng/mL for 48 and 96 h by two-way ANOVA (*P<0.05,**P<0.01)].Ctrl: control.
Angiogenesis affects this critical process of follicular development through the angiogenic factor isoform,VEGFA[21].The VEGFA isoforms are produced by the alternative splicing of the VEGFA gene resulting in the generation of proangiogenic (VEGF165) and the antiangiogenic (VEGF165b) isoforms[21].Previous studies have demonstrated that the VEGF165 isoform is expressed in bovine[9] and murine ovaries[22].Several reports demonstrated that VEGF165 is increased during follicular development[23-25].Moreover,several VEGF inhibitors have been shown to inhibit folliculogenesis,including VEGF-TRAP[26],antibodies[27],and VEGFR tyrosine kinase inhibitors[21].However,targets of these inhibitors are both the proangiogenic and the antiangiogenic isoforms of VEGF[28].Previous studies have demonstrated that VEGF165 shows proangiogenic effects associated with follicle development leading to the increased rate of antral formation,stimulates meiosis resumption,and activates the production of mature oocytes in goats[13,14].VEGF165b revealed the antiangiogenic effect in several pathological condition such as ischemic retinopathy mouse model and models of age-related macular degeneration and prostate,lung,renal,skin,and colon cancers[28-33].Previous studies demonstrated that VEGF isoforms (VEGF120 and VEGF164) are associated with the follicular development in the bovine ovary[34].However,a recent report indicated that the balance of VEGF165 isoforms regulated follicle development,luteogenesis,and fertility in mammals[17].Another study reported that the VEGFA isoform showed angiogenic and antiangiogenic actions on bovine folliculogenesis through different signal transduction pathways[35].A previous study also showed that the alteration of VEGFA isoform and its receptor affected the bovine follicular cyst[36].Therefore,these findings suggest that the regulation of the splicing of VEGF results in follicular development and thus fertility.
Previous report suggested that estrogens had modulated VEGF-A for physiological conditions[20].Moreover,recently in vitro studies suggested that the hormonal regulation affected VEGFA gene expression in bovine granulosa and theca cells[18,37].However,there are no reports on the effect of estrogen on the expression of the VEGF165 isoform in cattle.Our results demonstrated that VEGF165 was activated at 100 ng/mL estrogen in the large follicle for 96 h.In addition,VEGFR2 was upregulated in all medium and large follicles after treated with 100 ng/mL estrogen for 96 h.Our results demonstrated that estrogen stimulates the expression of VEGF165 and its receptor during bovine follicular development.This result suggests that estrogen may influence the expression of VEGF165 and its receptor during follicular development.However,the nonevaluation of the effects of estrogen in vivo on the levels of VEGF isoforms involved in folliculogenesis and oogenesis as well as on the quality of the offspring could be considered a limitation of this study.
In conclusion,there is an increase in the expression of VEGFA and its receptor in the development of different size follicles,which may lead to supposition that VEGF isoforms are enhanced to perform its function.The correlation between the effect of VEGF on the vascular development within the bovine ovary may be critical to determine the normal mechanisms of ovarian morphogenesis and to understand ovarian reproductive disorders in the future.
Conflict of interest statement
The authors declare no conflicts of interest.
Acknowledgements
We thank professor David Bates from the University of Nottingham for supporting chemical and research machine.
Funding
This study received no extramural funding.
Authors’ contributions
Khwanthana Grataitong carried out study design,manuscript writing,and critical discussion.Chinnarat Changsangfa and Bongkoch Turathum made study design,experiment execution,data analysis,manuscript writing,and critical discussion.Morakot Sroyraya performed experiment execution and manuscript review.All authors contributed to the article and approved the submitted version.
 Asian Pacific Journal of Reproduction2023年6期
Asian Pacific Journal of Reproduction2023年6期
- Asian Pacific Journal of Reproduction的其它文章
- Myo-inositol versus metformin effects on clinical features,endocrine and metabolic profiles in infertile women with polycystic ovary syndrome: A randomized controlled trial
- Embryonic,genetic and clinical outcomes of fresh versus vitrified oocyte: A retrospective cohort study
- Cryopreserved ovine spermatogonial stem cells maintain stemness and colony forming ability in vitro
- Seasonal stress on semen quality profiles,seminal biochemical and oxidative stress attributes in endangered Teressa goat of Andaman and Nicobar Islands
- Oxidative stress in male infertility and therapeutic approach: A mini-review