
番茄果实降异戊二烯类香气物质合成与代谢调控研究进展
2023-12-15 04:33高媛何昭颖崔广禄代艳侠平华马智宏
现代食品科技 2023年11期
高媛,何昭颖,崔广禄,代艳侠,平华,马智宏
(1.北京市农林科学院质量标准与检测技术研究所,北京 100097)(2.北京市大兴区种植业技术推广站,北京 102600)
番茄果实含有丰富的糖类、Vc、类胡萝卜素、酚酸、矿物质等多种营养素,其独特的风味也受到广大消费者的青睐。其中挥发性香气物质是番茄风味品质的重要组成部分,与糖、酸一起,共同影响着消费者对番茄产品的感官选择及喜爱程度[1-3]。
目前,番茄中已经有400多种挥发性组分被分离鉴定,根据其前体物质不同主要分为三类,包括源于脂肪酸降解产生的C6/C9类化合物,源于氨基酸代谢生成的芳香族类,以及源于类胡萝卜素代谢的降异戊二烯类。其中,降异戊二烯类芳香物质虽然在番茄果实中含量不高,但由于其极低的嗅觉阈值,少量存在即可产生较大的感官影响,从而对番茄风味起着举足轻重的作用。
降异戊二烯类物质种类较多,合成路径也比较复杂。目前对番茄果实中降异戊二烯类物质的合成及调控机理的分析还较少。本文介绍了番茄果实中类胡萝卜素降解产生的降异戊二烯类香气物质组成及其生物合成途径,并对其代谢相关酶和调控机制进行概述,以期为开展番茄风味研究工作提供参考。
1 番茄中的降异戊二烯组分
番茄果实中的香气物质主要来源于三条合成路径,分别是:1)脂氧合路径:以脂肪酸为前体物,通过一系列氧化降解生成直链脂肪醇、醛、酸、酯;2)氨基酸代谢路径:即以支链氨基酸和芳香族氨基酸为前体,产生支链脂肪族和芳香族类化合物;3)异戊二烯代谢路径:以类胡萝卜素为前体,生成降异戊二烯(Norisoprenoids)及其衍生物[4]。这些挥发性组分相互作用,共同决定了番茄果实的风味品质。香气成分对果实风味所产生的感官贡献,一般以香味活力值(香气物质含量与其阈值的比率,Odor Activity Value,OAV)来表征。目前,人们普遍认为番茄中的16种化学物质,具有最积极的香气活力系数(OAV>1)[2,5-6],包括顺式-3-己烯醛、己醛、1-戊烯-3-酮、反式-2-己烯醛、反式-2-庚烯醛、顺式-3-己烯醇,来源于脂氧合路径,主要提供青草香气;2,3-甲基丁醛、2-异丁基噻唑,1-硝基-2-乙基苯、2-苯乙醛、2-苯乙醇、3-甲基丁醇、水杨酸甲酯均来自于氨基酸代谢路径,赋予番茄果实花香/泥土味;β-紫罗兰酮,β-大马士酮,6-甲基-5-庚烯-2-酮,则来源于类胡萝卜素代谢路径,呈现花果香气。从中可发现,番茄中的花香、果香大多来自于类胡萝卜素降解生成的降异戊二烯类物质及其衍生物,特别是β-紫罗兰酮和β-大马士酮,分别位于16种香气组分OAV排序中的第3和第5位[2,5]。降异戊二烯类芳香物质在多种蔬菜、水果的风味品质中均起到了举足轻重的作用。
类胡萝卜素中因其大量共轭双键的存在,导致其性能很不稳定,可以经过降解反应形成大量带有羰基结构的如C8、C9、C10、C11、C13、C14等衍生物,其中一些有着很重要的香气功能,如降异戊二烯类化合物。在番茄果实中,C8降异戊二烯类有6-甲基-5-庚烯-2-酮,C10降异戊二烯类物质有β-环柠檬醛、香叶醛,而C13降异戊二烯类较多,包括β-大马士酮、β-紫罗兰酮、α-紫罗兰酮、香叶基丙酮、5,6-环氧-3-羟基-β-紫罗酮、假紫罗兰酮等。Jeyaprakash等[7]从番茄中检测到了5种降异戊二烯类的芳香化合物,Simkin等[8]还检测到了多种β-紫罗兰酮的衍生物。在这些降解产物中,C13型对番茄果实的风味品质贡献最大。
番茄果实中降异戊二烯类物质的含量虽然低于C6醛类,但由于其具有极低的阈值和怡人的香味特性,对番茄果实香气尤为重要。其中,最受关注的是β-大马士酮和β-紫罗兰酮。β-大马士酮在水果中存在很普遍,表现出复杂的花香、热带水果或“熟苹果”的香气。在水溶液中阈值极低,仅为2 ng/L,在乙醇溶液中的阈值为50 ng/L[9]。β-紫罗兰酮和α-紫罗兰酮都带有紫罗兰/覆盆子的香味,其中β-紫罗兰酮在水溶液和酒中的阈值分别为7 ng/L和90 ng/L[10]。β-大马士酮和β-紫罗兰酮在番茄中的浓度大多都超过了其阈值。此外,也有一部分香气物质OAV<1,但仍可通过协同作用或者改变其它化合物的阈值而加强果香,如香叶基丙酮、香叶醛等。这也表示了香气物质被感知不仅是因为它自身的浓度和阈值,还取决于不同组分之间的相互作用[11,12]。
2 降异戊二烯类芳香物质的生物合成路径
类胡萝卜素既是重要维他命和抗氧化剂的合成前体,也是降异戊二烯类芳香物质的合成前体[13,14]。科学研究已经证实,在富含β-类胡萝卜素的番茄果实中,其β-紫罗兰酮、β-环柠檬醛的含量也较高,而α-紫罗兰酮却只在积累δ-胡萝卜素的红橙色果肉的番茄突变体中被检测到;且具有柠檬味道的香叶醛也来源于果蔬中的番茄红素[15]。目前报道的与香气直接有关的类胡萝卜素一般有β-胡萝卜素、玉米黄质、α-胡萝卜素、番茄红素等,它们能够通过化学降解或酶促降解产生降异戊二烯类化合物,其生物合成途径已基本得到探明(图1)。
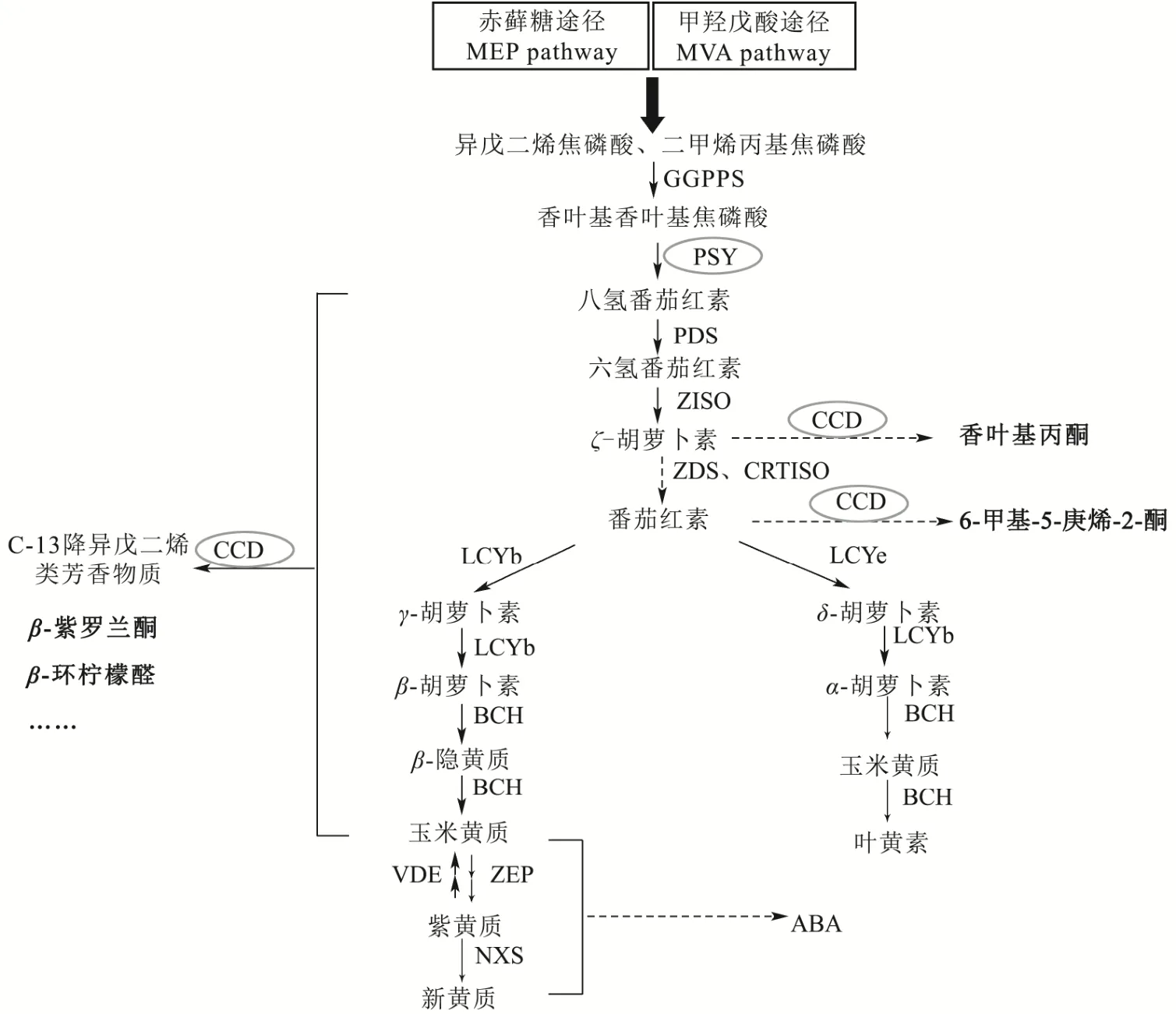
图1 类胡萝卜素及降异戊二烯生物合成途径Fig.1 Biosynthesis pathway of carotenoids and norisoprenoids.
类胡萝卜素是由8个异戊二烯单体缩合而成的C40的四萜类色素,而异戊二烯来源于质体中的赤藓糖路径(Methyl-Erythritol-Phosphate,MEP Pathway)和细胞质中的甲羟戊酸路径(Mevalonate,MVA Pathway),由于类胡萝卜素在质体中产生,MEP途径被认为是类胡萝卜素所需异戊二烯的主要来源。但也有证据表明,两条途径的中间体可在质体和细胞质溶液中互相转移。通过MEP和MVA路径,得到的异戊二烯焦磷酸(Isopentenyl Diphosphate,IPP)和二甲基烯丙基焦磷酸(Dimethylallyl Diphosphate,DMAPP)在香叶基香叶基焦磷酸合酶(Geranyl-Geranyl Diphosphate Synthase,GGPPS)的相互作用下产生香叶基香叶基焦磷酸(Granylgranyl Pyrophosphates,GGPP);之后二分子GGPP在八氢番茄红素合成酶(Phytoene Synthase,PSY)催化下生成C40的类胡萝卜素-八氢番茄红素(Phytoene),而这也是生成的第一个无色的类胡萝卜素,此反应即为类胡萝卜素代谢合成途径的第一步,也是限速步骤。八氢番茄红素经脱氢、异构等酶催化反应后生成番茄红素,而番茄红素则是类胡萝卜素向后进一步合成与代谢过程的主要分支产物。随后再在番茄红素环化酶的作用下生成β-胡萝卜素和α-胡萝卜素两条分支,进而再经环化、羟化等作用生成叶黄质、玉米黄质、紫黄质、新黄质等化合物[16]。最后这些类胡萝卜素在类胡萝卜素裂解双加氧酶(Carotenoid Cleavage Dioxygenase,CCD)的催化下生成降异戊二烯类物质。另外,类胡萝卜素又是脱落酸(Abscisic Acid,ABA)的前体物,可以在9-顺式环氧类胡萝卜素双加氧酶(9-Cis-Epoxycarotenoid Dioxygenase,NCED)等催化酶的作用下选择性断裂不同位点上的双键,生成ABA。
3 降异戊二烯类物质合成的关键酶基因
3.1 八氢番茄红素合成酶(PSY)
降异戊二烯类芳香物质来源于植物类胡萝卜素代谢。PSY催化二分子GGPP形成八氢番茄红素,此过程也是进入类胡萝卜素生成路径的第一步。PSY是植物类胡萝卜素生物合成的限速酶,最先从番茄中克隆并得到验证[17,18]。随后也在多种植物如玉米、水稻、小麦、胡萝卜、葡萄等中分离获得[19-22]。在番茄中已证实有3条PSY基因存在[23],其中PSY1在番茄成熟过程中上调表达,与果实中番茄红素的积累密切相关[24];PSY2主要在叶片细胞内表达,参与叶绿体内类胡萝卜素的生成[25];PSY3则被推测在胁迫条件下在番茄根部表达,参与植物建立共寄生关系以及适应营养缺乏[26-28]。
PSY作为类胡萝卜素合成途径的关键酶,其基因转录变化趋势与番茄红素含量具有一致性[29],它的缺失会阻碍番茄果实成熟过程中类胡萝卜素的合成,而它的过表达则会增加包括番茄和马铃薯在内的多种作物器官中类胡萝卜素的生成[30,31]。果实的颜色主要取决于PSY1的表达[32,33]。在玉米中,PSY1仅在黄色胚乳中有表达,而在类胡萝卜素缺乏的白色胚乳中无表达[34];而在稻米中,PSY1的过表达会导致β-胡萝卜素积累,从而使得白色胚乳变成黄色[35]。Efremov等[36]对4个野生番茄品种和1个栽培品种的PSY1进行克隆及生物信息学分析结果表明,PSY1具有品种和组织特异性,与成熟果实中的类胡萝卜素含量直接相关;且PSY1在红色栽培品种中的表达显著高于绿果野生番茄。此外,红果番茄与黄果番茄的差别主要体现在番茄红素和β-胡萝卜素的合成积累上,成熟期红果番茄的含量是黄果番茄的4~5倍,这也主要源于PSY1的表达差异[37]。Chen等[31]发现了一个反式剪接的PSY1可以改变番茄果实的颜色,该PSY1的过表达增加了黄果樱桃番茄中类胡萝卜素的含量并导致其果实呈现红色。
3.2 类胡萝卜素裂解双加氧酶(CCD)
类胡萝卜素裂解双加氧酶(Carotenoid Cleavage Dixoygenase,CCD)是类胡萝卜素裂解氧合酶(Carotenoid Cleavage Oxygenases,CCO)中重要的一支,参与植物中多种类胡萝卜素的降解,是控制类胡萝卜素降解速率和降异戊二烯类物质合成的关键酶。在拟南芥中,CCO家族共有9条,其中4条属于CCD亚家族(AtCCD1、AtCCD4、AtCCD7、AtCCD8),5条属于NCED亚家族(AtNCED2、AtNCED3、AtNCED5、AtNCED6、AtNCED9)。NCED家族主要参与植物ABA的合成,CCD主要催化类胡萝卜素在9-10和9’-10’位裂解产生降异戊二烯及其衍生物。CCD在一些模式植物(拟南芥[38])、经济作物(如玉米[39]、水稻[40]等)、花卉(如菊花[41]、兰花[42]等)和水果(如西瓜[43]、柑橘[44])中已有过研究报道。拟南芥AtCCD1和AtCCD4均可裂解类胡萝卜素生成降异戊二烯类化合物。在欧亚种葡萄、柑橘和菊花花瓣等的研究中也证实了类胡萝卜素的降解速率是由CCD决定的[38,41,44,45]。目前已报道的番茄中的CCD基因包括SlCCD1a、SlCCD1b、SlCCD4a,SlCCD4b,SlCCD7和SlCCD8。其中SlCCD7和SlCCD8在番茄果实/植株的所有器官和组织中均有表达,但在根中表达量最高,主要参与独脚金内酯的合成[46]。研究表明,植物中的CCD1和CCD4通过降解类胡萝卜素调控植物颜色及风味,与降异戊二烯类香气物质的合成密切相关。
番茄中的CCD1含有2条家族成员,串联在同一条染色体上。SlCCD1a和SlCCD1b序列长度分别为10635 bp和12921 bp,均含有14个外显子,翻译的氨基酸序列长度为545 aa。研究表明,SlCCD1a在番茄的根部和老叶中表达量最高,而SlCCD1b则在成熟的红色果实中表达量最高。在不同植物中,CCD1的主要裂解位点是9,10 (9'~10')碳双键。番茄的SlCCD1[47]、月桂(Laurus nobilis)的LnCCD1[48]能裂解各种不同结构的顺式和全反式的类胡萝卜素以及各种形式的阿朴类胡萝卜素形成α-紫罗酮、β-紫罗酮等各种芳香型化合物以及6-甲基-5-庚烯-2-酮和香叶醛[49]。在番茄中,LeCCD1b还能够催化紫黄质和新黄质在9,10/9',10'位断裂形成5,6-环氧-3-羟基-β-紫罗酮。在葡萄果实中,VvCCD1也能够催化玉米黄质的降解,从而生成3-羟基-β-紫罗兰酮。在番茄果实中,SlCCD1研究较多,被认为与降异戊二烯类香气物质合成密切相关。Simkin等[50]的研究表明,LeCCD1A和LeCCD1B的沉默则会导致β-紫罗兰酮含量的降低。
番茄中有两条CCD4,为SlCCD4a和SlCCD4b,分别在雌蕊和老叶中表达量最高。研究结果表明,CCD1主要定位在细胞质中,而CCD4主要定位于类胡萝卜素大量合成的质体中,比CCD1更能够催化裂解类胡萝卜素。同时,由于CCD4功能的正常行使与功能缺失均会造成果实、花等器官颜色的变化,其基因表达差异也是导致不同桂花品种颜色变化的关键因素[51]。CCD4已经被报道能够在藏红花[52]、苹果[53]、柑橘[54]中分别降解番茄红素、β-胡萝卜素、ε-胡萝卜素,并产生6-甲基-5-庚烯-2-酮、β-紫罗兰酮和α-紫罗兰酮。有研究表明,在葡萄果实中,较高水平的降异戊二烯含量与VvCCD4b的上调表达密切相关[55]。在番茄果实中,普遍认为SlCCD1与降异戊二烯类香气物质的合成相关,SlCCD4a和SlCCD4b由于分别在花和叶片中表达量最高,而在番茄果实中报道较少。Wei等[56]的研究表明SlCCD4在番茄果实中同样有表达,表达量低于SlCCD1,但未进一步研究与降异戊二烯类香气物质合成的关系。因此,SlCCD4s在番茄果实降异戊二烯积累中的作用如何,还需进一步探明。
4 番茄果实降异戊二烯的生物合成调控
4.1 品种因素
基因对果实挥发性香气物质组成起决定性作用。不同番茄品种降异戊二烯组成和含量有明显差异。常培培等[57]对5种不同果色的樱桃番茄中的挥发性香气物质进行研究后表明,品种因素对香气成分的种类和浓度影响差异较大,特别是对以类胡萝卜素为主要合成前体而生成的降异戊二烯及其衍生物的影响最为显著。降异戊二烯,如香叶基丙酮、β-紫罗兰酮和β-环柠檬醛,在红色、粉色果实品种中的含量要显著高于黄果和绿果樱桃番茄[57]。Scott等[58]对38个不同基因型的番茄果实挥发性物质进行测定,结果发现香气物质种类和含量差异显著,其中,6-甲基-5-庚烯-2-酮、β-大马士酮、香叶基丙酮、β-紫罗兰酮因受基因型影响含量差异很大。
4.2 环境因素
番茄香气物质的合成受到果实发育过程中温湿度、光照、栽培方式等环境因素的影响[2]。有研究表明,温度可能在关键呈香物质合成中起到了重要作用。杨明惠等发现,不同栽培季节番茄果实芳香物质含量存在差异,春季栽培番茄由于相关酶活性较高,果实风味明显高于秋冬季节栽培的番茄[59]。在20 ℃下转色的紫色番茄,风味指标较好[60]。也有研究认为低温和高温均不利于番茄红素的合成,番茄红素在30 ℃以上停止合成,以25 ℃左右为宜[61]。此外,番茄作为后熟型果实,采后低温贮藏也会抑制番茄果实挥发性芳香物质生物合成相关基因表达,Zou等[62]的研究表明,相比于25 ℃,在10 ℃储藏的樱桃番茄中,有包含CCD1在内的33条基因表达显著降低,并导致了其代谢相关产物如假紫罗兰酮、β-紫罗兰酮等含量的降低。低温暴露会导致风味物质包括6-甲基-5-庚烯-2-酮、β-紫罗兰酮等含量的降低[63,64];而过高的温度处理(如52 ℃处理5 min)同样会降低风味物质的积累[65,66]。
光是调节色素和风味等次生代谢产物的重要环境因素。光照会影响番茄果实中类胡萝卜素的积累和香气物质含量的变化。番茄属中日性植物,对日照时间长短要求比较宽泛,最适宜的光照时间为12~16 h,适当增加光照时间,在一定程度上能促进植株生长,有助于番茄有机物的积累。而光照不足会引起番茄类胡萝卜素及风味物质含量的降低[67]。Bertin等[68]的实验表明,充足的日照环境可以提高采收期番茄果实中芳香物质合成的前体产物,包括脂肪酸及其衍生物、苯丙氨酸衍生物以及类胡萝卜素及衍生物的合成。所以,在适当时间段进行补光处理可以提高番茄果实番茄红素的浓度和相应芳香化合物的生成积累[69]。补光5 h后对番茄红素含量影响较大,比对照组增高了69.5%的番茄红素含量[70],这可能是由于补光处理可以诱导番茄果实中的光敏色素积累[71],同时加剧番茄类胡萝卜素裂解双加氧酶(LeCCDs)对类胡萝卜素化学键的断裂作用[72],从而引起了6-甲基-5-庚烯-2-酮和β-紫罗兰酮含量的增加。
不同光质对番茄营养与风味品质影响也不同。在其他植物(如罗勒、甜菜)中,适当增加蓝光比例,其类胡萝卜素的含量有所上升[73,74],然而在番茄中,蓝光处理有利于提高番茄Vc及可溶性蛋白含量,红光则能极大的促进番茄红素的合成。研究表明,红色光质能促进番茄果实风味物质种类和含量的增加,其中,6-甲基-5-庚烯-2-酮的含量较白光增加了20.2%[75]。Wang等[76]的研究则认为夜间补充红蓝光(7:2)3 h,可显著提高番茄的风味品质。
此外,外源激素处理也会影响降异戊二烯的积累,如外源ABA处理可以促进樱桃番茄果实成熟过程中颜色的发育以及类胡萝卜素的积累。外源ABA还增加了β-大马士酮(1.64倍)等挥发性化合物的积累,同时诱导了与挥发性香气物质生物合成途径有关的关键基因的表达[77]。而外源生长素2,4-二氯苯氧乙酸则会通过抑制乙烯的生成和类胡萝卜素的积累来延缓樱桃番茄的成熟过程,同时影响降异戊二烯类物质(包括β-大马士酮、β-紫罗兰酮等)的含量[78]。
4.3 分子水平调控
番茄类胡萝卜素代谢路径和挥发性芳香物质的生物合成均受光照、发育阶段、环境温度等诸多因素调控,由多种转录因子分别或共同调节特定的下游靶基因完成。番茄PSY活性受各种环境因素影响,如光强、气温、盐分、水份、光周期等。其中,受光照作用更为明显,如红光就可引起PSY的转录丰度上调,进而引起类胡萝卜素水平的增加。而高温会抑制PSY的表达,导致类胡萝卜素含量降低[79,80]。目前,人们已经发现了许多干扰和控制类胡萝卜素生成的转录因子,如MADS-box家族、AP2/ERF家族、MYB家族等[81]。其中,MADS-box家族是目前发现的能够影响类胡萝卜素代谢途径的转录因子中数量最大的一类。研究人员证实,MADS-RIN可以有效整合PSY1的启动子,进而调节类胡萝卜素的生物合成;但在其rin突变体的果实中基本检测不到PSY1的表达,果实中也基本缺乏番茄红素的合成[82],所以,MADS-RIN对番茄果实类胡萝卜素代谢过程起到了促进作用[83]。
在类胡萝卜素代谢路径中,与降异戊二烯类香气物质合成直接相关的就是CCD基因,但关于其在转录水平上的报道较少。植物顺式和反式作用元件主要负责基因的转录调控。启动子区域的顺式作用元件决定了时间和空间的基因表达模式,以及这些模式对胁迫的反应方式。在海棠中,干旱、涝渍和茉莉酸甲酯处理均会降低CCD7启动子的活性[84]。而反式作用元件,尤其是转录因子,与顺式作用调控区域结合并影响基因表达。目前,对调控CCD表达的转录因子还知之甚少,仅在桂花、柑橘以及葡萄中有报道,这些转录因子分别属于WRKY家族、ERF家族和MADS家族。OfWRKY3和OfERF61可以通过与桂花OfCCD4的启动子结合,增强OfCCD4的表达,导致类萝卜素的降解以及桂花花瓣中β-紫罗兰酮的生成;而CsMADS6可以与柑橘CsCCD1的启动子结合,促进CsCCD1的表达[85-87]。近期研究表明,在葡萄果实中,转录因子VvMADS4可以下调VvCCD4b的基因表达[88],从而抑制降异戊二烯的生物合成。然而,目前还没有关于番茄中SlCCDs转录调控的报道。此外,以往的研究已报道了多种调控番茄类胡萝卜合成的转录因子,这些转录因子对类胡萝卜素降解生成降异戊二烯路径的代谢关键酶基因(CCDs)是否同样发挥作用尚无报道。
5 总结与展望
近年来,对降异戊二烯类香气物质的研究不断深入,涉及类胡萝卜素合成与降解代谢关键酶基因的克隆、表达和调控在多种植物中已有研究,并证明了降异戊二烯类芳香物质是类胡萝卜素在CCD的作用下降解得到的产物,其中起主要作用的是CCD1和CCD4。尽管CCD4在多种植物中均被报道具有降解类胡萝卜素生成降异戊二烯的功能,但在番茄中关于SlCCD4鲜有研究。SlCCD4在番茄果实中的表达特性及在降异戊二烯类香气物质合成中的作用如何,尚不明确。此外,关于番茄中降异戊二烯类香气物质的研究尚需进一步深入,对其合成关键基因CCD的启动子、转录因子研究仍然较少。系统分析番茄果实类胡萝卜素降解产物的积累变化规律以及转录调控过程,将为番茄及其他园艺作物在改善果实风味品质等方面提供更为丰富的理论依据。
猜你喜欢
少年文艺·我爱写作文(2023年2期)2023-02-24
疯狂英语·新读写(2019年9期)2019-09-21
知识经济·中国直销(2018年10期)2018-11-06
南方文学(2017年6期)2018-01-07
小学生(看图说画)(2017年5期)2017-11-06
作文周刊·小学一年级版(2016年20期)2017-06-03
长城(2017年1期)2017-02-17
橡塑技术与装备(2016年21期)2016-02-25
石油化工(2015年9期)2015-08-15
