
外源CBD 缓解烟草Cd 胁迫的代谢组学分析
2023-12-15 11:47何怡管娇郭笑维杜玉娇余诗尧周鹏保志娟
中国烟草学报 2023年6期
何怡,管娇,郭笑维,杜玉娇,余诗尧,周鹏,保志娟
云南农业大学,烟草学院,昆明 650201
重金属污染影响植株的正常生长发育和产量,而镉(Cd)是烟草最为敏感的重金属之一[1-2]。通过对云南典型地质高背景区土壤重金属含量的检测,发现表层土壤Cd 含量相对较高[3],增加了农用土壤重金属污染风险。在重金属胁迫下,植株中的糖类、氨基酸、有机酸等初生代谢物可作为渗透调节物提高其耐受性,同时次生代谢物激活抗氧化酶,减少重金属积累过程的中间产物介导应激反应[4],以应对胁迫伤害。因此,研究代谢物变化已成为探索重金属胁迫下植物响应机制和应对措施的重要手段[4-5]。
部分外源有机物、植物激素施用能缓解作物的重金属毒害[6-7]。工业大麻是全球新兴发展产业之一,大麻二酚(CBD)是工业大麻中主要的活性大麻素[8]。2018 年,世界反兴奋剂机构将CBD 排除在World Anti-Doping Code International Standard Prohibited List 2021之外[9]。云南省和黑龙江省已通过地方立法,相继于2009 年与2017 年分别制定《云南省工业大麻种植加工许可规定》及《黑龙江省禁毒条例》等规定,允许在公安部门监管下开展工业大麻的培育、种植、销售和加工。2021 年《云南省“十四五”工业和信息化技术进步规划》将工业大麻产业列为重点发展产业[10]。CBD 具有抗痉挛、抗焦虑、抗炎等药理活性,国外在医疗、食品、化妆品和农业等领域都有应用。CBD作为多基配体也能与金属离子络合形成环状螯合物,在治理重金属污染方面潜力巨大[11-12]。在符合管理规定的条件下,本课题组开展了CBD 在烟草农业方面的研究。前期研究表明,CBD 对重金属胁迫烟株的生长发育具有一定的调节作用,但是具体的机理尚不清楚。代谢组技术可以量化植物生理生态的变化,为此本研究采用靶向和非靶向分析结合的代谢组学探讨CBD对Cd 胁迫下烟草代谢物的影响,以期为降低烟草重金属污染提供理论依据与参考措施。
1 材料与方法
1.1 供试材料
2021 年在云南省昆明市进行盆栽试验,供试烟草品种为K326(玉溪中烟种子公司)。土壤采自云南自然背景低肥力红壤(pH 6.84,有机质6.97 g/kg,有效磷11.56 mg/kg,速效钾101.33 mg/kg,碱解氮 18.91 mg/kg,全Cd 0.27 mg/kg),装盆前过2 mm 筛,充分混匀。CBD 购于云南汉盟制药有限公司,纯度99%。将其溶于含0.1%的二甲基亚砜和0.07%的吐温80 混合水溶液中,配制成1 mg/L 的CBD 溶液(现配现用)。
1.2 试验设计
试验共设3 个处理:正常生长(CK),Cd 胁迫(60 mg/kg),Cd 胁迫烟株喷施外源CBD(Cd-CBD)。每株1 盆,每个处理6 盆。Cd 胁迫处理:将二水乙酸镉溶解以污灌的形式施入花盆土壤中,根据土壤湿度,每盆灌注2 L,使土壤中Cd 含量为60 mg/kg,腐熟一个月之后进行烟苗(苗龄55 d)移栽。Cd-CBD 处理:在移栽后50 d 上午,取Cd 胁迫处理烟株,第一次喷施1 mg/L CBD 液,使溶液附着于叶片正反面(湿润不流滴),3 d 后喷施第二次。同时,对CK 与Cd 处理以同样的方法喷施含有二甲基亚砜和吐温80 的空白混合水溶液。间隔3 d 后,用打孔器均匀地取总重约5 g 中部叶(自上而下第六位叶),根系部位用剪刀在主根与侧根距根尖10 cm 处剪断。每个部位均取6 个生物学重复样本,迅速保存到液氮中。随后将烟叶与根系冷冻真空干燥,研磨成粉末状,-80℃保存待进行初生代谢物、生物碱与植物激素的测定。
1.3 重金属含量的测定
各处理选取3 株烟分部位收获,先用自来水洗净根、茎、叶表面,然后用0.01 mol·L-1EDTA 与去离子水将烟株清洗干净,再用吸水纸吸干水分,于105℃下杀青30 min,65℃烘干至恒重,称量每株烟地上、地下部分干重。将干燥后每株烟的地上部分、地下部分分别混合均匀,采用电感耦合等离子体质谱法分别测定地上和地下部分的Cd 含量。
耐受系数=各处理烟株地上(下)部生物量/对照烟株地上(下)生物量
单株积累量=单株植物生物量×单株植物Cd 含量
1.4 代谢物提取及测定方法
初生代谢物及生物碱测定:准确称取50 mg 冻干烟根粉末参照Li 等[13]方法进行初生代谢物的提取与衍生,以木糖醇做内标,基于GC-MS 对初生代谢物进行非靶向测定。另分别取50 mg 冻干烟根与烟叶粉末参照郑庆霞等[14]方法对生物碱进行超声萃取,以2,4’-二联吡啶做内标,基于GC-MS 对8 种生物碱进行靶向检测。
植物激素测定:准确称取100 mg 冻干烟叶粉末,加入400 μL 甲醇/乙腈/水(1:1:1,v/v/v),匀浆后,加入600 μL 甲醇/乙腈/水(1:1:1,v/v/v),冰浴中超声60 min,16000 g 4℃离心30 min,过滤后取上清,真空干燥。样本复溶于80 μL 50%乙腈溶液中,20000 g离心15 min,取上清液以备LC-MS/MS 进样分析。
超高效液相色谱条件:样品采用 NexeraX2 LC-30AD 超高压液相色谱仪(Shimadzu)进行分离。流动相:A 液为10 mol/L 乙酸铵水溶液pH 8.0,B 液为100%乙腈。
质谱分析:采用5500 QTRAP 质谱仪(AB SCIEX)在正/负离子模式下进行质谱分析。正电离电压:5500 V,负电离电压:4500 V。采用MRM 模式检测待测离子对。
1.5 数据分析
GC-MS 数据经MS-DIAL 软件的默认参数对样本的原始仪器数据文件进行反卷积分析,图片采用MetaboAnalyst 5.0、Origin 2021 进行绘制。非靶向代谢组数据经NIST 14 标准质谱库进行检索,利用保留指数(RI 值)与Chemspider 公共数据库中进行比对定性,最佳鉴定结果以质谱相似度和RI 值接近度最高的化学结构为准。生物碱与植物激素靶向代谢组数据分别采用ChemicalStation 与MultiQuant 软件提取色谱峰面积及保留时间,并使用生物碱与植物激素标准品校正保留时间,进行代谢物鉴定。通过方差分析(ANOVA,P<0.05)与偏最小二乘判别分析(VIP值>1)筛选差异代谢物,并利用MetaboAnalyst 软件(https://www.metaboanalyst.ca/)进行差异物的相关性分析与代谢通路富集分析。
2 结果与分析
2.1 外源CBD 对Cd 胁迫下烟株吸收Cd 的影响
由表1 可知,在Cd 胁迫下,烟株的生物量显著减少。添加CBD 后,生物量较单一Cd 胁迫升高了13.93%(地上)和31.63%(地下),且耐受系数较单一Cd 胁迫上升了13.75%(地上)和30.56%(地下)。原土壤中(CK)含有微量Cd,这时烟株地上部分的Cd 含量较低,主要富集在地下部分。Cd 胁迫下,烟株对Cd 的吸收量较CK 极显著增加,且地上部分Cd的含量大于地下部分Cd 的含量。喷施CBD 后,地上部分Cd 单株积累量较单一Cd 处理下降了8.81%。
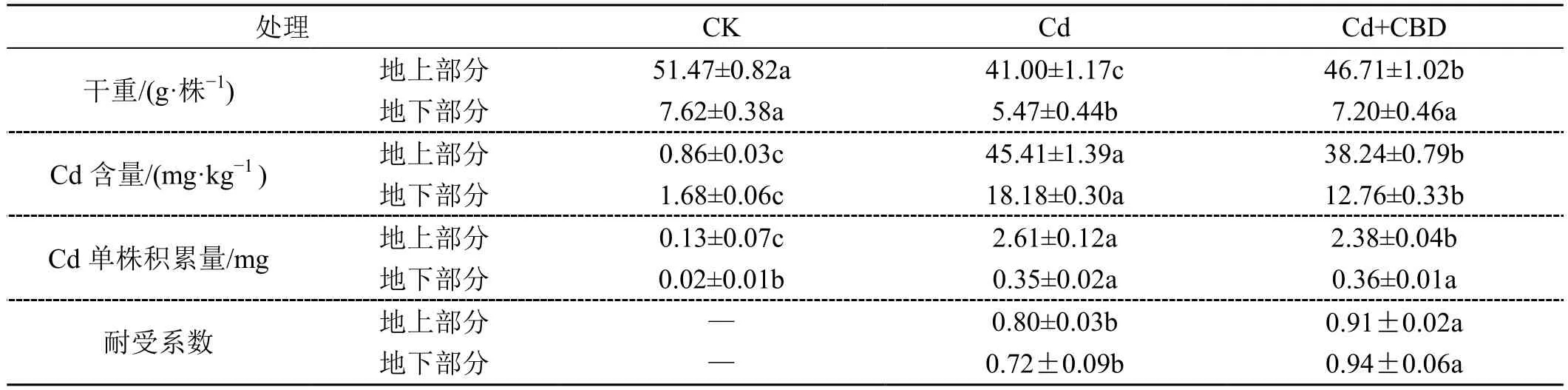
表1 Cd 在烟株体内分布情况与各处理烟株耐受系数Tab.1 The distribution of Cd in tobacco plants and the tolerance coefficient of each treated plant
2.2 外源CBD 对Cd 胁迫下烟根初生代谢物的影响
2.2.1 初生代谢物的主成分分析
利用非靶向组学的方法进行根部初生代谢物的测定,共检测出103 种化合物。经NIST 库定性,保留匹配度大于80%的化合物,共定性出33 个化合物(表2),其中氨基酸3 种,有机酸10 种,糖7 种,烷烃7种,其它类化合物6 种;苹果酸、蔗糖为代谢分泌物的主要成分,含量均超过了1 mg/g。利用内标(Xylitol)进行归一化后,对数据进行PCA 分析。如图1A,各处理样本点均落于95%置信区间内,且CK 与另外两种处理各自聚为一类,说明数据处理结果可信。为排除初生代谢物的组内差异的干扰,进一步分清不同处理下初生代谢物的组间差异,作PLS-DA模型(图1B),并进行置换检验,结果显示R2=0.998、Q2=0.994,均大于原始R2、Q2,未发生过拟合现象,证明模型可靠。

图1 PCA(A)与PLS-DA(B)Fig.1 PCA (A) and PLS-DA (B)

表2 根部初生代谢物平均含量Tab.2 Average content of primary metabolites in roots
2.2.2 初生代谢物差异分析
为了分清Cd 和CBD 对烟株的影响,利用PLS-DA模型以及ANOVA 分析筛选出VIP 值>1 且P<0.05 的化合物,将其作为潜在差异初生代谢物。本次实验3个处理共筛选出15 个含量差异的初生代谢物(表2),其中大多化合物属于微量成分,有机酸(6 种)与烷类(5 种)数量较多,另外还包含2 种氨基酸、1 种糖类和1 种胺类。与CK 相比,Cd 胁迫下烟根中的葡萄糖酸含量下降,苏糖酸、棕榈酸、丁二酸、乙醇酸、肌醇半乳糖苷、丙氨酸、正二十一烷、正二十二烷、正二十三烷、正二十四烷、正二十五烷等11 个化合物的含量升高。与Cd 处理相比,施入CBD 后,烟根中肌醇半乳糖苷、苏糖酸、棕榈酸、丁二酸、乙醇酸、丙氨酸含量降低,正二十一烷、正二十二烷、正二十三烷、正二十四烷、正二十五烷、葡萄糖酸含量升高。
2.3 CBD 对Cd 胁迫烟草生物碱的影响
对CK、Cd、Cd-CBD 处理烟叶的生物碱进行分析(图2)。由图2 可知,烟碱、降烟碱、假木贼碱、新烟草碱为烟株根、叶中的主要生物碱。烟根中的烟碱和降烟碱含量显著高于叶部,而假木贼碱含量叶和根部差异不大,其他5 种生物碱皆是叶>根。在Cd 胁迫下,根部烟碱显著升高,除二烯烟碱无明显变化外,其它6 种生物碱含量下降;叶片内生物碱变化较根部显著,烟碱、降烟碱、新烟草碱含量升高,其余5 种生物碱含量降低。施用CBD 后,根部降烟碱、麦斯明、N-甲基假木贼碱、假木贼碱、新烟草碱等含量均有回升;而叶片中的生物碱均呈下降趋势,除降烟碱外,其它7 种生物碱的总含量与Cd 相比差异显著。

图2 不同处理下8 种生物碱在烟株叶片与根部的浓度Fig.2 The concentrations of 8 alkaloids in leaves and roots of tobacco plants under different treatments
2.4 CBD 对Cd 胁迫烟叶激素的影响
利用LC-MS/MS 技术对3 种处理烟叶中的12 种植物激素进行测定,采用标准曲线法计算样品含量,并进行显著性分析,结果如表3 所示。与CK 相比,Cd 胁迫使叶片中脱落酸、赤霉素1、二氢茉莉酸、3-吲哚丁酸和水杨酸含量显著降低;而3-吲哚乙酸、茉莉酸、茉莉酸-异亮氨酸、褪黑素、反式玉米素含量显著增加。在施入CBD 后,除赤霉素3、茉莉酸和二氢茉莉酸外,上述激素含量有向CK 回调的趋势,其中脱落酸、3-吲哚乙酸以及水杨酸的含量与CK 无显著差异。而反式玉米素核苷在三种处理下变化不明显。
2.5 相关性分析
本次实验基于GC-MS 和LC-MS/MS 的靶向和非靶向组学方法共获得35 个差异合物(15 种初生代谢物、8 种生物碱、12 种植物激素)。利用MetaboAnalyst 5.0 对上述35 个差异化合物进行相关性分析,发现共有1189 对呈显著相关性的化合物对,其中有346 对呈负相关,844 对呈正相关(图3)。呈显著正相关的代谢物之间具有相似的生物学功能、化学成分或同属的特征[15]。与烟碱含量正相关的化合物有27 个,其中包含6 个生物碱,3 个有机酸(葡萄糖酸、乙醇酸、乳酸),1 个糖类(肌醇半乳糖苷),1 个醇类(甘油),6个烷烃类(正二十一烷、正二十二烷、正二十三烷、正二十四烷、正二十五烷、正二十六烷),10 个植物激素(茉莉酸-异亮氨酸、水杨酸、赤霉素1、赤霉素3、3-吲哚乙酸、脱落酸、3-吲哚丁酸、反式玉米素、茉莉酸、二氢茉莉酸)等,以上物质可能是重金属胁迫下,调控烟碱积累的重要代谢物。与烟碱含量呈负相关的化合物有7 个,包括1 个胺类(酪胺)、2 个氨基酸(焦谷氨酸、丙氨酸)、3 个有机酸(硬脂酸、丁二酸、奎宁酸)、1 个植物激素(褪黑素)。

图3 差异代谢物相关性Fig.3 Correlation of differential metabolites
2.6 CBD 及Cd 胁迫对烟株代谢通路的影响
结合CK、Cd、Cd-CBD 三种不同处理下烟草的差异代谢物进行KEGG 富集分析。CK 与Cd 代谢通路富集分析结果表明(图4A,表4),差异代谢物主要富集的代谢通路为半乳糖代谢、硫代谢、TCA 循环、甘油脂代谢、色氨酸代谢、谷胱甘肽代谢、类胡萝卜素生物合成等。在这7 种代谢途径中富集了6 个差异代谢物,肌醇半乳糖苷、甘油、丁二酸、焦谷氨酸、3-吲哚乙酸与脱落酸。焦谷氨酸、丁二酸、褪黑素与烟碱呈负相关,3-吲哚乙酸、脱落酸与生物碱类化合物呈正相关。
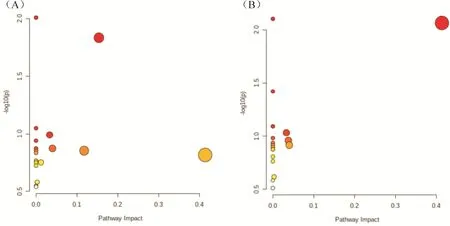
图4 代谢通路富集分析Fig.4 Metabolomics pathway analysis
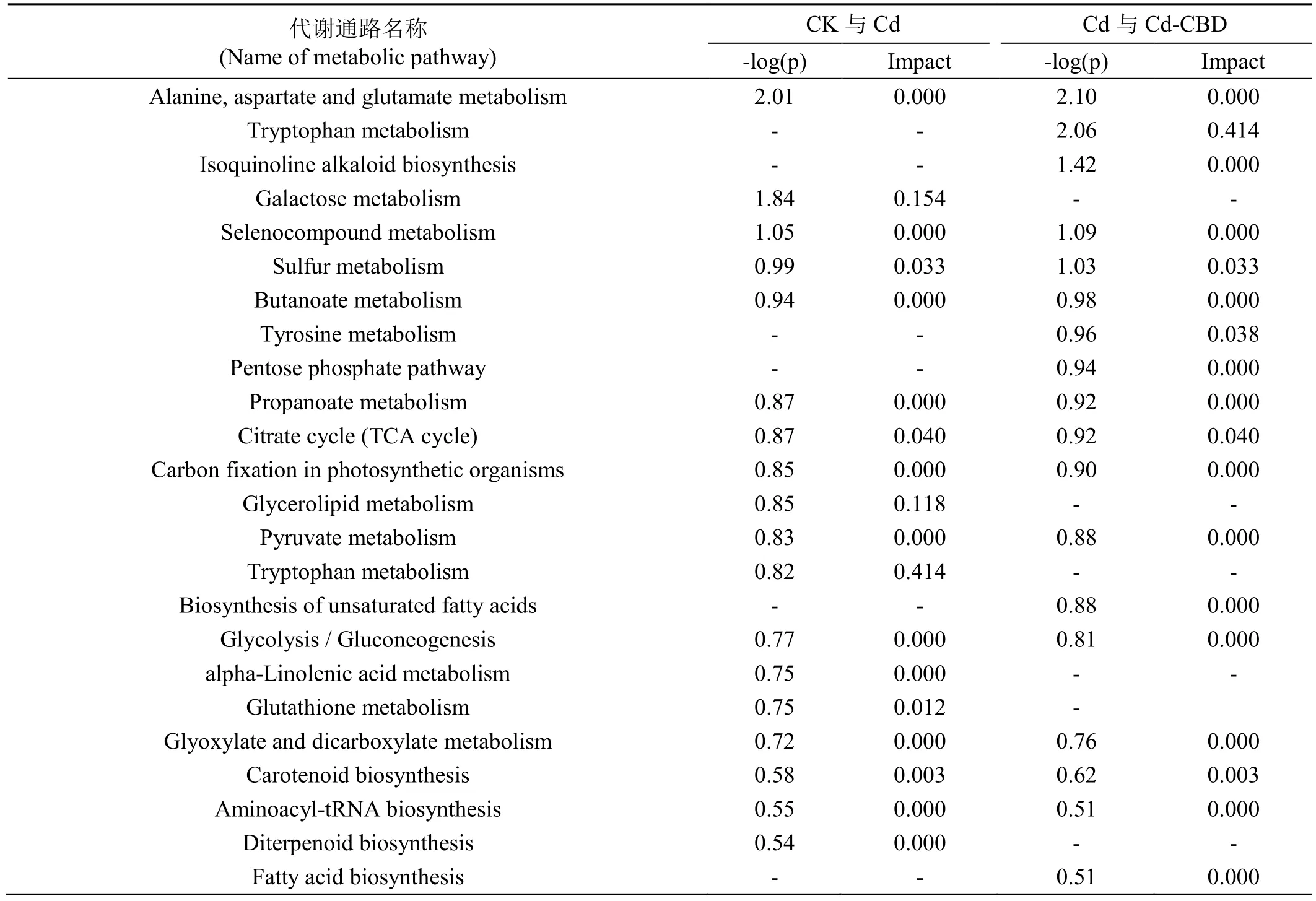
表4 差异组分参与的代谢途径Tab.4 Metabolic pathways involving differentially expressed metabolites.
Cd 与Cd-CBD 代谢通路富集分析结果表明(图4B,表4),差异代谢物主要富集的代谢通路为色氨酸代谢、硫代谢、酪氨酸代谢、TCA 循环、类胡萝卜素生物合成等。在这5 种代谢途径中富集了5 个差异代谢物,丁二酸、酪胺、3-吲哚乙酸、褪黑素与脱落酸。由相关性分析可知丁二酸、酪胺、褪黑素与烟碱呈负相关,3-吲哚乙酸、脱落酸与生物碱合成呈正相关。
3 讨论
逆境胁迫下,植株体内代谢物质发生重排,参与植物在非生物胁迫下的应答反应[16-17]。当作物受到重金属胁迫时,有机酸作为植物抵抗非生物胁迫的一类重要代谢物大量生成[18]。研究表明,暴露于Cd 的植物会释放更多的苏糖酸来迁移Cd,增加植物对Cd 的吸附和积累[19]。同时,根际分泌的低分子有机酸会与Cd 螯合形成“Cd-低分子有机酸”复合物,从而调控Cd 向烟株体内的迁移[20]。在本研究中,烟草根部共检测出6 种有机酸类差异代谢物。其中,苏糖酸、丁二酸、乙醇酸在Cd 胁迫下的含量较CK 均显著升高,Cd 很有可能是通过与这3 种有机酸螯合,以达到向烟株地上部分转移的目的。糖类物质在植株体内积累,在逆境中既可起渗透调节作用,也可作为能量供给、合成代谢的重要底物[21-22]。本研究结果显示,Cd 胁迫下,烟草根中肌醇半乳糖苷含量较CK 极显著升高。肌醇半乳糖苷的合成是棉子糖家族寡糖代谢的第一步,其作为渗透调节剂,响应非生物胁迫[23]。氨基酸是植物体内氮的主要形式之一,具有信号转导、细胞渗透调节、矿物质营养吸收和重金属解毒等多种功能,是重要的代谢产物[24-26]。本研究中Cd 胁迫烟草根系中丙氨酸含量较CK 极显著升高。在施用外源CBD 后,以上发生显著上调的有机酸、糖、氨基酸等初生代谢物含量均有所下降。
CBD 对Cd 胁迫的缓解作用,一方面可能是利用了多酚类化合物的还原性,渗透到脂质双分子层抑制脂质过氧化,阻碍自由基的传播[27]。另一方面可能是作为多羟基配体,为金属离子提供了不同的正离子结合位点[28-29],影响了重金属的生物利用度、含量和迁移率[30],使游离态的Cd 离子减少,烟株胁迫响应降低。植物激素作为化学信使调节细胞间的通信,率先做出响应[31]。研究结果显示,褪黑素、茉莉酸-异亮氨酸、脱落酸、3-吲哚乙酸等9 种激素均在施入CBD 后含量有向CK 水平值变化的趋势。茉莉酸类物质与脱落酸的作用类似,与植物的抗性密切相关[32]。褪黑素、3-吲哚乙酸可与重金属离子发生螯合作用,同时可作为内源性自由基清除剂参与抗氧化调节,缓解重金属对植物的伤害[33-34]。本试验中,在Cd 胁迫下,随着3-吲哚乙酸、茉莉酸、茉莉酸-异亮氨酸、反式玉米素这类与烟碱呈正相关的植物激素的增加,烟碱的含量较CK 显著升高。生物碱是烟草对外界环境长期适应过程中形成的一大类抗逆次生代谢物,在遇到环境胁迫和生物侵扰时会瞬间大量合成[35]。在添加CBD 后,根部中大多数生物碱含量有向CK 回调的趋势,并且使烟叶生物碱的含量整体呈下降趋势。
初生代谢物与次生代谢物的含量变化必然引起代谢途径的改变。通过KEGG 通路分析发现,Cd 胁迫下,外源CBD 的施用对色氨酸代谢、硫代谢、酪氨酸代谢、TCA 循环、类胡萝卜素生物合成这5 条代谢途径的影响最显著。色氨酸代谢是响应非生物胁迫的一条重要代谢通路[36]。酪氨酸代谢途径中的生育酚是植物中重要的抗氧化剂,且酪氨酸是植物中多种特异性次生代谢物的来源(如苄基异喹啉类生物碱)[37]。而硫代谢的恢复,有利于提高植株对养分的吸收利用,从而使TCA 循环等能量代谢通路趋于正常,保证了烟草中生物碱类等特异性次生化合物的稳定合成。这对于缓解烟草镉胁迫具有重要作用。
4 结论
本研究通过对烟株Cd 积累量,以及初生代谢物、生物碱和植物激素进行非靶向与靶向测定,分析外源CBD 对烟草Cd 胁迫的缓解效应。结果表明,CBD 能提高烟株的生物量,显著降低烟株地上部分的Cd 积累量。烟草植株在Cd 胁迫下,各类代谢产物产生了不同的变化。在外源添加CBD 后,部分有机酸、糖类、氨基酸、生物碱与植物激素等代谢产物恢复至正常水平。通过KEGG 通路分析发现,CBD 主要通过调节色氨酸、酪氨酸和硫代谢过程,在一定程度上有效缓解Cd 对植株的毒害。
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
江西农业(2022年2期)2022-02-25
世界农药(2019年4期)2019-12-30
中成药(2017年12期)2018-01-19
中成药(2017年7期)2017-11-22
中成药(2017年8期)2017-11-22
中国烟草学报(2016年1期)2016-11-16
西南农业学报(2016年4期)2016-05-17
中国当代医药(2015年24期)2015-03-01
中国烟草学报(2012年1期)2012-04-09
