
入侵植物飞机草和南美蟛蜞菊种子萌发特征对热带珊瑚岛生境的响应
2023-12-21 05:34孙彰镁旷远文王俊简曙光张玲玲
热带亚热带植物学报 2023年6期
孙彰镁, 旷远文, 王俊, 简曙光, 张玲玲*
(1. 中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广东省应用植物学重点实验室,广州 510650;2. 中国科学院大学,北京 100049)
我国热带海洋珊瑚岛主要由第四纪珊瑚、贝壳碎屑等在生物的长期作用下堆积形成[1],其土壤由磷质石灰土和滨海盐土组成,碳酸钙、盐分含量高[1–2],但土壤肥力弱,且大部分为砂壤土, 孔隙比高、持水能力差[3]。受高温、强光、季节性干旱等影响,加上岛屿面积较小且远离大陆,物种迁徙难度大,热带海洋珊瑚岛原生植物群落形成与发育缓慢,群落结构简单[4]。受全球变化和人为干扰双重影响,热带珊瑚岛生境已出现飞机草(Eupatorium odoratum)、南美蟛蜞菊(Sphagneticolatrilobata)入侵踪迹[5],随着其扩张进程加快,热带珊瑚岛植被生态系统结构、健康稳定受到严重影响[6]。
植物入侵过程复杂,目前已经存在多种植物入侵机制的假说[7–8],如入侵植物内在优势[9]、入侵种与本地种相互作用[10–11]、入侵地环境的可侵入性[12]等。近年来,基于入侵植物内在优势的繁殖压力假说(propagule pressure hypothesis, PrPr)[11,13–14]逐渐得到认同,种子是植物最重要的繁殖体之一,入侵植物通过改变种子萌发特征来适应新生境[15],萌发过程是物种入侵的关键因素[16]。种子萌发过程受温度、水分、光照等多因素调控,响应机制复杂[17–19]。Bradley 等[20]研究表明温度变化对植物入侵有重要影响,且与种子萌发过程显著相关[16,21]。多数研究表明,入侵物种较原生物种具有更高的表型可塑性,对气候变暖响应更敏感[22],温度上升可为入侵创造有利条件[23]。但目前对入侵植物种子在不同生境条件下萌发特征的研究较少,种子萌发及出苗过程在入侵过程中所起作用的关注仍较为缺乏。研究入侵植物在不同生境条件下种子萌发特征及出苗过程有助于解析种子萌发性状改变的原因,为揭示其入侵潜力变化和成功入侵热带珊瑚岛生境机理提供科学依据。
发芽率(germination percentage, GP)、发芽势(germination energy, GE)、发芽指数(germination index, GI)、活力指数(vigor index, VI)和出苗速度(T50)是评价种子萌发性能和幼苗生长的常用指标[24]。发芽率反映种子总发芽能力;发芽势大小影响着种子发芽快慢和整齐度;发芽指数反映种子活力;活力指数从种子生长势和发芽速度综合反映种子萌发能力和生长趋势[25–26]。本研究以3 种菊科(Compositae)植物:入侵植物飞机草、南美蟛蜞菊和原生植物林泽兰(E.lindleyanum)为对象,通过“增温+海砂基质+水”模拟热带珊瑚岛生境,以“水培+常温”模拟大陆生境,测定3 种植物种子的上述萌发指标对不同生境的响应特征,探讨入侵植物飞机草、南美蟛蜞菊与原生植物林泽兰种子在热带珊瑚岛生境与大陆生境下的萌发及出芽特性差异;以及入侵植物飞机草、南美蟛蜞菊在热带珊瑚岛生境下与大陆生境扩张潜力差异,为预测入侵植物在热带珊瑚岛的扩张潜力提供理论基础,并为入侵植物的预警和防控提供科学参考。
1 材料和方法
1.1 试验设置
热带珊瑚岛属于热带海洋性季风气候,年均温为(27±2.3) ℃[27],该地区土壤由珊瑚和贝类等海生动物残体组成,钙、磷含量高,pH 值为8.5 左右[1],与大陆生境土壤差异迥然。本研究以热带珊瑚岛海砂为基质,通过人工增温至28 ℃模拟热带珊瑚岛生境;以纯水为基质,保持常温(25 ℃)模拟大陆生境作为对照。
本研究选取3 种菊科植物为研究对象,其中入侵植物飞机草(Eupatoriumodoratum)又名香泽兰,是菊科泽兰属多年生草本或亚灌木植物;南美蟛蜞菊(Sphagneticolatrilobata)是菊科蟛蜞菊属植物,2种植物适应性强、繁殖快、生长力旺盛,一旦入侵草地、林地和荒地会快速形成单种优势群落,降低入侵地的生物多样性[5,28–29]。原生植物林泽兰(E.lindleyanum)为菊科泽兰属多年生草本植物,分布广泛、适应性强[30]。
试验选取外观饱满无虫害的飞机草、南美蟛蜞菊和林泽兰种子,分别播撒至添加50 mL 纯水的海砂基质(砂培)和50 mL 纯水基质(水培)中,置于恒温培养箱[白天光照强度1 000μmol/(m2∙s),相对湿度54%]培养。培养箱温度设置2 种处理:常温(白天25 ℃, 12 h;晚上22 ℃,12 h)和增温(白天28 ℃,12 h;晚上25 ℃, 12 h),即常温+海砂、增温+海砂、常温+纯水、增温+纯水4 种处理方式,每种温度处理3 个重复,每种处理重复50 粒种子,试验周期共14 d。种子萌发以胚根出现为标志。在萌发过程中,每24 h 监测1 次种子发芽数,并记录已萌发的种子胚根长度(从胚根萌发处到根尖长度,cm), 随后计算出苗速度(T50),即总数50%的种子出苗所对应天数,T50越小说明出苗速度越快,反之越慢。
计算发芽率(GP)、发芽势(GE)、发芽指数(GI)和活力指数(VI)[24,31]:GP (%)=萌发种子数/样本种子数×100%,GE (%)=7 d 内萌发种子数/样本种子数×100%,GI=ΣGt/Dt,VI=GP×平均胚根长度,平均胚根长度=萌发胚根长度/萌发种子数, 其中,Gt为在时间t时的发芽数,Dt为发芽日数。
1.2 数据处理和分析
数据经Shapiro-wilk 和方差齐性检验,对不满足正态分布的进行平方根或反正弦转换。采用双因素方差分析(Two-way ANOVAs)评估培养基质、温度及其交互作用对种子萌发性状如发芽率、发芽指数和活力指数的影响,并采用邓肯(Duncan)新复极差法进行多重比较检验(α=0.05)。方差分析采用SPSS 27.0 统计软件,数据可视化分析均在R 语言(v.4.1.3)和Origin 2022 中进行。
2 结果和分析
2.1 萌发性状变异来源
基质与温度交互作用可以解释飞机草发芽率67.2%的变异来源;飞机草发芽指数的变异主要来源是培养基质(24.2%)、温度(24.2%)及其交互作用(46.1%);培养基质(62.0%)、基质与温度的交互作用(30.18%)可以解释飞机草种子活力指数的变异来源(P<0.05)。南美蟛蜞菊和林泽兰种子发芽率、发芽指数和活力指数的变异主要来源是培养基质(P<0.05) (图1)。

图1 基质、温度及其交互作用对3 种植物种子发芽率、发芽指数和活力指数的影响。1: 飞机草; 2: 南美蟛蜞菊; 3: 林泽兰; *: P<0.05。Fig. 1 Effects of medium, temperature and their interaction on seed germination character of three species. 1: Eupatorium odoratum; 2: Sphagneticola trilobata;3: E. lindleyanum; *: P<0.05.
2.2 不同处理对3 种植物种子萌发的影响
从表1 可见,在水培条件下,飞机草种子在增温下的发芽指数、活力指数均显著高于常温;南美蟛蜞菊和林泽兰种子的各项萌发及出苗指标在常温与增温处理间差异不显著。在砂培条件下,飞机草种子在常温处理下发芽势、活力指数和出苗速度均显著高于增温处理;南美蟛蜞菊和林泽兰种子的萌发及出苗指标在常温与增温处理间无显著差异,但林泽兰种子在14 d 常温处理过程中未达到50%的发芽率。在增温处理下,3 种植物种子在水培条件下的最终发芽率、发芽势、发芽指数、活力指数、出苗速度均显著高于砂培。
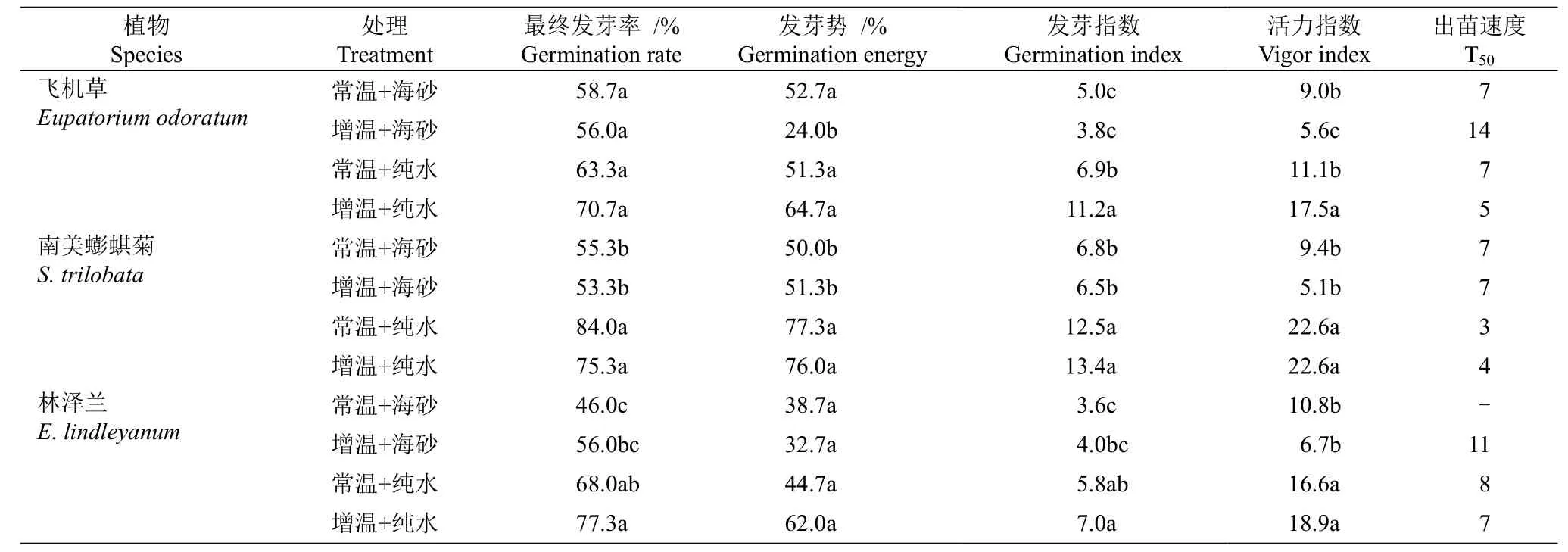
表1 增温对3 种植物种子萌发的影响Table 1 Effect of elevated temperature on seed germination of three species
分析表明,不同培养基质下,3 种植物发芽率对增温的响应存在种间差异。飞机草种子在大陆生境条件下萌发更为迅速,在萌发前期(2~11 d)常温+纯水处理下发芽率显著大于增温+海砂处理(P<0.05)(图2: A);在增温+海砂处理下第5 天发芽率仅为18.7%, 而常温+纯水处理为34.0%;在萌发后期(12~14 d)常温+纯水和增温+海砂处理下飞机草种子发芽率无显著差异,其最高发芽率分别为63.3%和56.0%。南美蟛蜞菊种子在水培下发芽率显著高于砂培(图2:B);在水培和砂培下南美蟛蜞菊种子萌发过程中增温与常温之间的差异均不显著,其中,常温+纯水处理下第2 天开始萌发,第3 天发芽率达51.3%,最高发芽率为84.6%;增温+海砂处理下第3 天开始萌发,第7 天发芽率达51.3%,最高发芽率为53.3%。林泽兰种子在水培处理下种子发芽率均显著高于砂培处理;在砂培条件下,增温+海砂与常温+海砂处理间种子发芽率差异不显著(图2: C),其中,常温+纯水处理下第4 天开始萌发,第8 天发芽率达54.0%,最高发芽率为68.0%;增温+海砂处理下第4 天开始萌发,第11 天发芽率达50.7%,最高发芽率为56.0%。
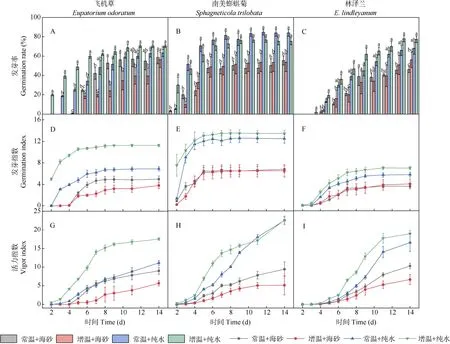
图2 不同处理对飞机草、南美蟛蜞菊和林泽兰种子发芽率(A~C)、发芽指数(D~F)和活力指数(G~I)的影响Fig. 2 Effects of different treatments on germination percentage (A-C), germination index (D-F), and vigor index (G-I) of the seeds of Eupatorium odoratum,Sphagneticola trilobata and E. lindleyanum
3 种植物种子发芽指数对不同处理的响应存在差异。不同处理下飞机草种子发芽指数变化趋势相同,从第7 天开始趋于稳定(图2: D),增温+纯水处理下飞机草种子发芽指数最高,其次是常温+纯水处理,常温+海砂处理发芽指数高于增温+海砂处理。不同处理下南美蟛蜞菊种子发芽指数在第5 天开始趋于稳定(图2: E),而林泽兰种子发芽指数增长较慢,在第11 天开始趋于稳定(图2: F)。南美蟛蜞菊和林泽兰在水培处理下种子发芽指数均显著高于砂培处理,在水培条件下增温处理发芽指数更高,而砂培条件下增温与常温处理间无显著差异。
3 种植物种子活力指数在不同处理存在显著差异。在萌发过程中,飞机草种子在增温+纯水处理下活力指数显著高于其他处理(图2: G);常温+纯水处理下的飞机草种子活力指数均显著高于增温+海砂处理。南美蟛蜞菊种子在水培处理下活力指数显著高于砂培处理(图2: H),其中,常温+纯水处理下南美蟛蜞菊种子活力指数均显著高于增温+海砂处理。林泽兰种子在萌发前期(第2~9 天)活力指数增加缓慢(图2: I),至处理第9天活力指数均快速增加,在萌发后期(11~14 d)常温+纯水处理林泽兰种子活力指数均显著高于增温+海砂处理,但萌发前期(2~9 d) 2 种处理间活力指数差异不显著。
2.3 种子发芽指标比较
增温+海砂处理下3 种植物种子之间发芽率、发芽指数和活力指数随萌发时间均呈上升趋势。在萌发前期(2~6 d),南美蟛蜞菊种子发芽率、发芽指数增加较快(图3: A, B),且显著大于飞机草和林泽兰种子;萌发后期(7~14 d), 南美蟛蜞菊种子发芽率、发芽指数逐渐趋于稳定,与飞机草和林泽兰无显著差异。在萌发初始阶段(2~4 d),南美蟛蜞菊种子活力指数显著大于飞机草和林泽兰(图3: C),萌发5 d 以后,3 种植物种子活力指数均缓慢升高, 且三者活力指数差异不显著。

图3 增温+海砂处理下3 种植物的种子发芽率(A)、发芽指数(B)和活力指数(C)。柱上不同字母表示差异显著(P<0.05)。Fig. 3 Germination rate (A), germination index (B) and vigor index (C) of seeds of 3 species under warming + marine sand treatment. Different letters upon column indicate significant difference at 0.05 level.
3 讨论和结论
本研究选取的3 种菊科植物均为多年生草本植物,其中入侵植物飞机草和原生植物林泽兰均属于泽兰属植物,主要依赖于种子繁殖[32]。飞机草种子产量极高,每平方米可产生2 000~260 000 粒种子,且20%~46%种子可存活[33],具有较强繁殖能力。南美蟛蜞菊在野外情况下基本上以无性繁殖为主,其种子常不育,有性繁殖效率低下[29],在良好的生存环境下主要依靠匍匐茎节生根占领新的空间,产生新的植株。南美蟛蜞菊在通过克隆分株快速繁殖的同时也产生一些种子,以便在入侵早期或不利的生存条件下通过有性繁殖进行传播,加快对恶劣环境的适应,逃逸出不利的生存环境进而入侵新的领地[34–35]。
发芽行为对入侵种群的建立影响显著[36],发芽特征在一定程度上能表征植物的入侵潜力[37–38],温度、土壤养分等地理或环境因子能对入侵植物的种子萌发过程产生综合影响[39–40]。不同生境条件下植物种子萌发及出苗性状生态适应策略存在差异[15],反映植物对环境的适应,外来植物与原生植物萌发过程的差异有助于外来植物的入侵[38]。本研究结果表明,3 种植物种子的萌发及出苗特性在不同处理下存在差异,飞机草的发芽指数变异的主要来源是培养基质、温度及其交互作用,活力指数的变异来源是培养基质、基质与温度的交互作用,即温度与培养基质均是影响飞机草种子萌发的重要因子。培养基质可以解释入侵植物南美蟛蜞菊和原生植物林泽兰种子发芽率、发芽指数及活力指数变异,这可能是因为南美蟛蜞菊等草本入侵植物种子往往具有萌发温度广泛、萌发速度快、萌发率高等特性,能够良好地适应变温环境[41];而在萌发过程中种子不需要特别的养分供给[32],盐胁迫对种子萌发过程存在抑制作用[42],海砂基质养分匮乏且盐分含量高,对种子萌发性状造成了显著影响。
飞机草种子在常温+纯水处理(大陆生境)条件下的发芽势、发芽指数、活力指数和出苗速度在常温处理下均显著高于增温+海砂处理(热带珊瑚岛生境),在萌发前期常温+纯水处理发芽率显著高于增温+海砂处理,2 种处理间最终发芽率无显著差异,说明在大陆生境下飞机草种子在萌发过程中发芽速度、整齐度、活力更高,能快速占据有利条件, 使其拥有较热带珊瑚岛生境更高的竞争优势,即入侵植物飞机草种子在热带珊瑚岛生境繁殖潜力略有下降,但经过较长时间后,飞机草种子也能达到较高的发芽率。这与Wang 等[43]报道增温在一定范围内有效促进了入侵植物种子萌发进程的结果有所差异,但Kriticos 等[44]和Lu 等[45]研究表明飞机草种子繁殖能力强、偏好温暖的环境,在2 ℃~40 ℃下均可萌发,最佳萌发温度为25 ℃,表明飞机草萌发阶段可塑性较强,能快速适应热带珊瑚岛生境条件,维持一定的繁殖能力。南美蟛蜞菊种子在常温+纯水处理(大陆生境)条件下最终发芽率、发芽势、发芽指数、活力指数和出苗速度均显著高于增温+海砂处理(热带珊瑚岛生境),表明南美蟛蜞菊种子在常温+纯水条件下拥有更强的萌发能力,即与大陆生境相比,南美蟛蜞菊种子在热带珊瑚岛生境的扩张潜力降低。
原生植物林泽兰种子在常温+纯水处理(大陆生境)条件下活力指数和出苗速度显著高于增温+海砂处理(热带珊瑚岛生境),但发芽率、发芽势、发芽指数并无显著差异,在萌发后期,常温+纯水处理下种子活力指数均显著高于增温+海砂处理,但萌发前期2 种处理间活力指数差异不显著。说明在萌发前期,林泽兰种子适应性较强,在2 种生境条件下均能较好的萌发,这与Davidson 等[46]报道非入侵物种在资源有限或压力条件下比入侵物种能更好地保持稳定的适应性的结果一致,热带珊瑚岛高温生境养分匮乏、资源受到限制,原生植物林泽兰在此环境下拥有较强的适应性和繁殖能力。
萌发策略上的差异使入侵植物能快速入侵并占据优势[47]。Godoy 等[48]认为较早的发芽物候或较快的萌发速度可为入侵植物带来竞争优势。对3 种菊科植物种子在增温+海砂处理下萌发和出芽特征进行对比发现,在萌发前期,南美蟛蜞菊种子发芽率和发芽指数均显著大于林泽兰;此外,在萌发过程中其发芽势也大于林泽兰,出苗速度T50小于林泽兰,表明在热带珊瑚岛高温生境下,南美蟛蜞菊种子能更快速、整齐地萌发,而强大的无性繁殖能力[29]更是其快速入侵的基础,即在热带珊瑚岛生境下入侵植物南美蟛蜞菊的扩张潜力远远大于林泽兰,对原生植物具有较大的危害潜力。在萌发过程中,入侵植物飞机草和原生植物林泽兰种子发芽率、发芽指数和活力指数差异不显著,表明2 种植物种子扩张潜力相当。种子库是植物入侵的重要决定因素之一,丰富、持久的种子库能有效提高植物的入侵潜力[49],因此,飞机草丰富的种子库及其种子强大抗干扰能力和传播能力[50]使其依然具有较高的入侵风险。
综上,入侵植物飞机草和南美蟛蜞菊具有强大的入侵潜力,在全球变暖背景下入侵种群可能拥有较原生植物更强的竞争优势,这将对当地生态系统造成一定威胁,在今后的研究中应当着重关注热带珊瑚岛生境入侵植物的防控工作。飞机草与原生植物扩张潜力相当,因此控制飞机草种子成熟及萌发是降低其扩散能力控制入侵的有效方法;南美蟛蜞菊具有很强的无性繁殖能力和一定的有性繁殖能力,因此建议在其花期前对地上、地下部分进行全株清除,避免茎叶重新生成分株或遗留种子,以彻底消除其生态危害。对飞机草和南美蟛蜞菊的开花进行抑制将是我们下一步研究的重点。
猜你喜欢
连云港职业技术学院学报(2021年3期)2021-11-29
中国建材科技(2021年1期)2021-03-31
建材与装饰(2020年21期)2020-08-03
江西建材(2020年11期)2020-02-16
中成药(2019年12期)2020-01-04
西南石油大学学报(自然科学版)(2019年4期)2019-11-04
中成药(2018年12期)2018-12-29
北方人(2018年16期)2018-08-20
中成药(2018年6期)2018-07-11
爱你(2016年4期)2016-12-06
