
桉树组培苗生产污染菌的调查与分析
2023-12-27 03:20蔡华杰黄向东马天驰易润华何普林
桉树科技 2023年4期
蔡华杰,黄向东,马天驰,易润华,何普林*
桉树组培苗生产污染菌的调查与分析
蔡华杰1,黄向东1,马天驰1,易润华2,何普林1*
(1. 中林集团雷州林业局有限公司,广东 遂溪 524348;2. 广东海洋大学滨海农业学院,广东 湛江 524088)
采用形态学和分子生物学方法对桉树组培苗污染菌的种类进行了调查,为减少桉树组培苗生产污染提供依据。结果表明:污染菌主要包括25个属50个种,其中污染细菌6个属7个种,污染真菌19个属43个种,细菌、真菌分别占14.0%、86.0%。继代苗的污染菌主要是丽水黄色杆菌()和青霉属(spp.)、曲霉属(spp.)各5个种;外植体的污染菌为11属21个种的真菌和1个属1个种的细菌。
桉树组培;污染菌;组培苗;形态学鉴定;扩增子
桉树(spp.)集生态价值、经济价值和观赏价值于一体[1-4],在我国广东、广西、福建、云南、海南等省(区)广泛种植[5]。桉树造林用苗主要来源于种子苗、扦插苗和组培苗,其中以组培苗最常用。
桉树组培可在短时间内快速获得大量无病种苗,但在生产中常遇到外植体和继代苗污染的问题,严重影响组培苗的生产[6-7]。广东省湛江市中林集团雷州林业局有限公司在生产桉树组培苗时经常出现污染,影响了种苗的生产效率。为此,文章采用形态学和分子生物学方法对桉树组培苗生产车间的污染菌进行研究,以期为桉树组培苗生产减少污染提供理论依据。
1 材料与方法
1.1 样品的采集
2021年9月—2022年9月,在广东省湛江市中林集团雷州林业局有限公司林业科学研究分公司的桉树组培苗生产车间中收集被污染的组培瓶共107个。将收集的组培瓶带回实验室进行污染菌株的分离、鉴定等研究。
1.2 污染菌的分离
在无菌环境下,从桉树组培苗生产中污染的组培瓶获得污染菌,将其接种到PDA培养基进行分离纯化,并保存在PDA斜面培养基。
1.3 污染菌的形态学鉴定
将保存的菌株接种到PDA平板培养基上,在25 ~ 28 ℃黑暗条件下培养7 d后,用LEICA DM750显微镜观察污染菌的形态特征,鉴定污染菌的种属。
1.4 污染菌的分子鉴定
将污染菌活化后,采用菌落PCR扩增污染菌的基因序列。扩增污染真菌的引物为ITS1(TCC GTA GGT GAA CCT GCG G)和ITS4(TCC TCC GCT TAT TGA TAT GC)[8]。扩增污染细菌的引物为27f(AGA GTT TGA TCM TGG CTC AG)和1492r(TAC GGY TAC CTT GTT ACG ACT T)。PCR反应体系采用TaKaRa公司的MightyAmp DNA Polymerase Ver.2试剂盒。PCR产物送至生工生物工程(上海)股份有限公司进行测序。所得序列在NCBI(https://blast.ncbi.nlm.nih.gov)进行BLAST在线比对和同源性分析确定污染菌的种属。
2 结果与分析
2.1 污染菌种的调查
共分离了107份样品,其中有3个组培瓶中的污染菌未分离成功,获得污染菌株111株,其中真菌99株,细菌12株。菌株测序后,在NCBI进行BLAST,并结合形态学特征,确定污染菌的种属。结果显示:这111株菌分别属于15个目、20个科、25个属、50个种,其中真菌有9个目、14个科、19个属、43个种,细菌有6个目、6个科、6个属、7个种。
间座壳菌属(spp.)、青霉属(spp.)、曲霉属(spp.)和炭疽菌属(spp.)被鉴定出污染的种的数量比较多,分别为7、6、5、5个,镰刀菌属(spp.)有3个种,叶点霉属(spp.)、枝孢霉属(spp.)和篮状菌属(spp.)各有2个种,其他属均出现1个种(图1)。
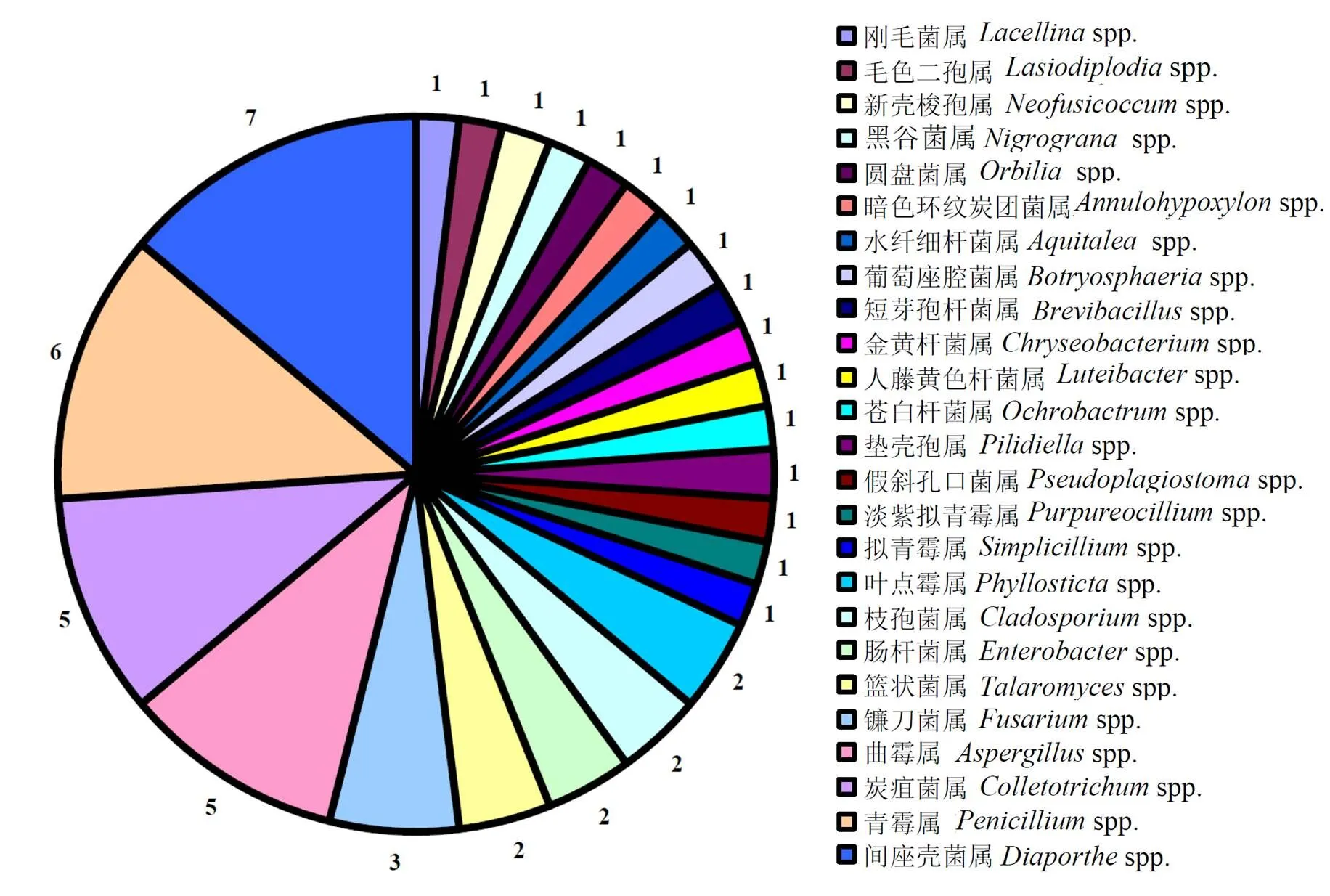
图1 各属内污染菌种数量的分布
2.2 污染菌的污染分析
在生产继代苗的组培瓶中,共鉴定出18个属31种污染菌,包括污染细菌4个属、4个种,污染真菌14个属、27个种,其中曲霉属和青霉属分别有5个种,镰刀菌属有3个种。分离到的真菌烟曲霉()菌株数最多,为10株,其次为奥林豪森曲霉()、橘青霉()和灰黄青霉(),均分离到5株(表1)。在培养外植体的组培瓶中共鉴定出12个属、24个种的污染菌,包括细菌2个属、3个种,真菌10个属、21个种,其中,间座壳菌属有7个种,炭疽菌属有5个种,其他11个属各1个种(表2)。细菌的2个属分别为为水纤细杆菌属(spp.)和肠杆菌属(spp.)。
青霉属和曲霉属的真菌仅生长在培养基上,在外植体上不生长。在实验室没有接种的空组培瓶中,分离到青霉属的2个种和曲霉属的1个种,分别为顶青霉()、灰黄青霉()和烟曲霉()。

表1 接种继代苗组培瓶中的污染菌

表2 接种外植体组培瓶中的污染菌
2.3 主要的污染菌
在生产继代苗的组培组培瓶中,丽水黄色杆菌出现的污染频次最高,主要出现在继代苗的基部,在每次继代培养时均有发现,而且比其他的菌污染更严重。出现污染频次较高的污染真菌为曲霉属和青霉属的真菌,在生产过程中主要出现在培养基中,其中烟曲霉、橘青霉和灰黄青霉在污染菌中出现的频率较高,分离率分别为11.7%、5.4%和5.4%。
在培养外植体的组培瓶中共分离到11个属、21个种的真菌,每种真菌出现污染的概率相当,除青霉属和曲霉属的真菌生长在培养基上污染外,其余均在外植体组织上生长,从而导致外植体污染。
2.4 主要污染菌的识别
2.4.1 丽水黄色杆菌
在不同的继代培养基上形成的菌落形态不同,颜色有橘黄色、乳白色和黑色三种(图2)。该菌在继代苗生产车间常年出现污染,主要出现在继代苗切口部位或继代苗与培养基的交界面,引起污染非常严重,有时继代接种后,污染瓶中所有的继代芽基部都长菌。
2.4.2 曲霉属
曲霉属真菌最初生长在培养基上,菌落圆形,呈绒毛状,与培养基结合不紧密;菌落扩展速度快,蔓延至继代苗上生长,后期覆盖整个培养基,菌落表面有粉状物。从污染组培瓶共鉴定出曲霉5种,分别为桉树生曲霉()、烟曲霉()、黑曲霉()、奥林豪森曲霉()和爪甲曲霉()(图3),其中烟曲霉污染最严重,占曲霉污染的54.2%。
2.4.3 青霉属
青霉属真菌在组培瓶中初期形成小、灰绿色的菌落,菌落表面密毡状或松絮状,与培养基结合紧密,菌落扩增速度慢,后期菌落表面布满粉状物。污染菌与继代苗接触后不在继代苗上生长,共鉴定出6种青霉属真菌,分别为橘青霉()、顶青霉()、灰黄青霉()、鲜红青霉()、草酸青霉()和白色青霉菌()(图4),其中橘青霉和灰黄青霉污染的频率最高,各占青霉污染菌的37.5%。

图2 丽水黄色杆菌在不同继代培养基的菌落形态

A:桉树生曲霉;B:烟曲霉;C:爪甲曲霉;D:奥林豪森曲霉;E:黑曲霉。A: Aspergillus eucalypticola; B: A. fumigatus; C: A. unguis; D: A. oerlinghausenensis; E: A. niger.

A:橘青霉;B:顶青霉;C:灰黄青霉;D:草酸青霉;E:白色青霉菌。A: Penicillium citrinum; B: P. corylophilum; C: P. griseofulvum; D: P. oxalicum; E: P. albidum.
2.4.4 青霉与曲霉的形态
曲霉属的污染菌分生孢子梗顶端膨大成顶囊,上生初生小梗和次生小梗,次生小梗上形成圆形分生孢子(图5A)。青霉属污染菌的分生孢子梗顶端不膨大,多次分枝后产生几轮对称或不对称的小梗,小梗上产生圆形分生孢子,形成帚状分生孢子头(图5B)。
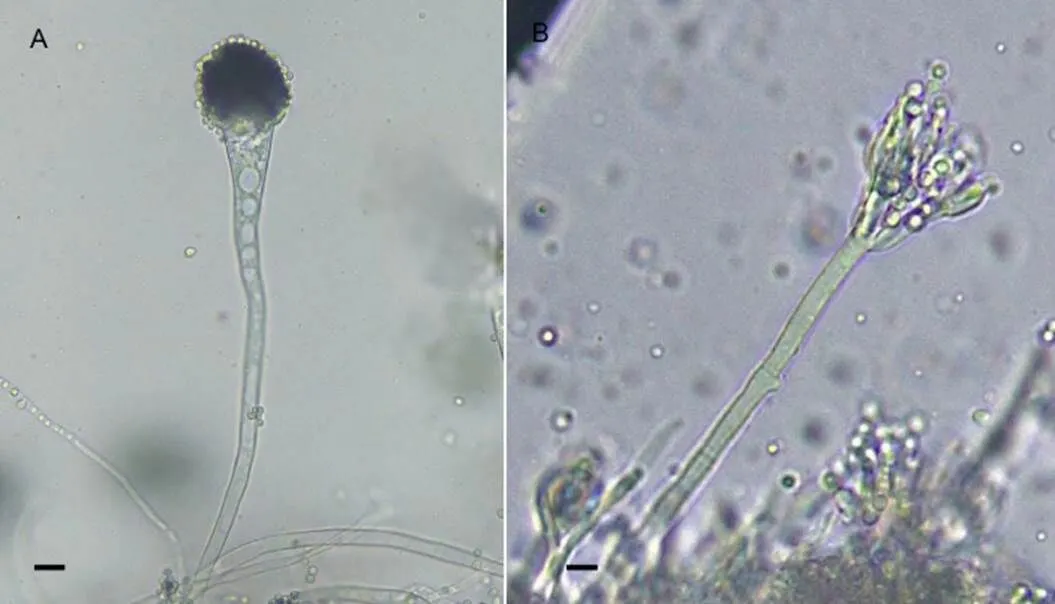
A:烟曲霉;B:灰黄青霉;标尺=10 μm。A: A. fumigatus; B: P. griseofulvum; scale bars=10 μm.
2.4.5 间座壳菌属和炭疽菌属的形态
间座壳菌属的有性阶段在污染菌中未发现,该属无性型的孢子主要有α,β和γ三种类型的分生孢子,以穿透间座壳菌()为例,其α型分生孢子线型,弯曲,β型分生孢子梭形(图6 A),γ型未发现。炭疽菌属的污染菌在PDA培养基上主要形成分生孢子,以暹罗炭疽菌()为例,其分生孢子长圆圆筒形,单胞,无色,有油滴(图6 B)。
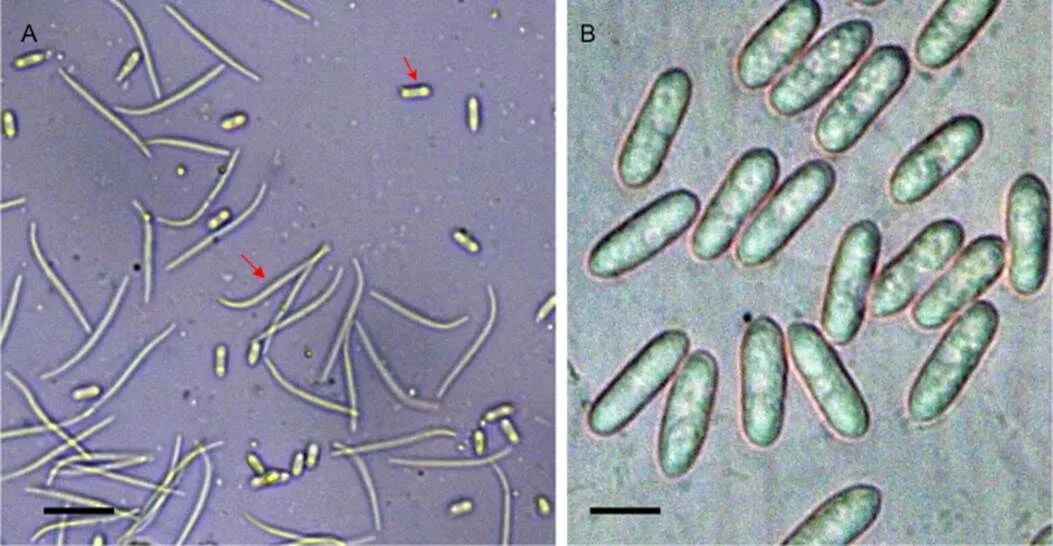
A:穿透间座壳菌的分生孢子;B:暹罗炭疽菌的分生孢子;标尺=10 μm。A:Conidia of D. penetriteum; B: Conidia of C. siamense; scale bars=10 μm.
3 讨论与结论
通常引起植物组培污染的微生物有病毒、细菌、酵母和真菌等,此次调查在桉树组培苗生产中出现的污染物主要是真菌和细菌,未发现病毒、酵母和螨虫等污染。本研究共鉴定出25个属50个种的微生物,包括6个属7个种的细菌和19个属43个种的真菌。LEIFERT等[9]报道植物组织快繁的污染细菌主要有不动杆菌属(spp.)、农杆菌属(spp.)、芽孢杆菌属(spp.)、棒状杆菌属(spp.)、杆菌属(spp.)、欧文氏杆菌属(spp.)、黄杆菌属(spp.)、微球菌属(spp.)、假单胞杆菌属(spp.)、葡萄球菌属(spp.)和黄单胞杆菌属(spp.);污染真菌有红酵母属(spp.)、假丝酵母属(spp.)、镰刀菌属、脉孢菌属(spp.)、曲霉属、微孢菌属(spp.)、枝孢霉菌属(spp.)和瓶霉属(spp.),其中肠杆菌属、曲霉属、镰刀菌属、枝孢霉菌属的微生物在调查中也出现,但在桉树组培苗生产中的污染菌更为复杂,种类更多。
桉树组培继代苗污染的细菌主要来自环境而非植物内生菌,污染真菌主要来自外界环境。由于外在的细菌污染出现比较早,一般外植体或继代苗培养2 ~ 3 d后即出现湿润的菌落,植物内生性污染细菌在外植体培养较长时间后出现的湿润菌落。组培桉树会随着环境温度和湿度、操作人员的不同而出现不同程度的污染[10]。本研究发现马氏水纤细杆菌和阴沟肠杆菌两种细菌在培养1 ~ 2 d后外植体和继代苗组培瓶中出现,多数生长与培养基接触的部位。马氏水纤细杆菌和阴沟肠杆在自然界和水体中广泛分布[11-12],继代苗或外植体被污染可能发生在剪取继代苗或外植体消毒的过程中。
污染桉树外植体的主要是真菌。在接种桉树外植体的污染瓶中共鉴定出13属23个种的真菌,占鉴定出污染菌种类的84.6%。污染桉树组培苗的曲霉属、曲霉属和枝孢霉菌属真菌主要来自空气,是常见的空气污染菌[13]。污染桉树外植体的真菌分别属于间座壳菌属(spp.)、炭疽菌属(spp.)、假斜孔口菌属(spp.)、叶点霉属(spp.)、新壳梭孢属(spp.)、毛色二孢属(spp.)和葡萄座腔菌属(spp.),它们都是一些植物的病原菌或内生真菌[14],这些真菌污染主要是外植体选取不合适或外植体消毒不彻底而造成。
影响植物组培污染的因素有季节变化、培养室温、湿度及卫生状况、操作人员技术、器械的灭菌和外植体等。本研究发现污染桉树组培苗生产的外在因素有培养室的环境、组培操作车间的卫生状况、器具的消毒和操作人员的熟练程度;内在因素有采集外植体带菌较多。因此,需注意选择带菌少的外植体、做好接种器械灭菌和培养环境消毒,接种时严格按无菌操作规程,减少桉树组培中的污染。
[1] 梁学明,杨永忠,杜鹍鹏,等.湛江市雷林桉树人工林生态价值估算量化研究[J].安徽农业科学,2021,49(14): 101-105,109.
[2] 任世奇,罗建中,彭彦,等.桉树无性系的单板出材率与价值研究[J].草业学报,2010,19(6):46-54.
[3] 郭乐东,周毅,钟锡均,等.西江流域桉树生态系统碳贮量与碳汇功能经济价值评价[J].广东林业科技,2009,25(6): 8-13.
[4] 谢耀坚.桉树的园林观赏价值[J].桉树科技,2005(1):1-6.
[5] 王永忠,杜阿朋.广东省桉树人工林固碳释氧价值核算[J].桉树科技,2020,37(1):36-38.
[6] LE ROUX J J, VAN STADEN J. Micropropagation and tissue culture of—a review[J]. Tree Physiology, 1991, 9(4): 435-477.
[7] GUPTA N P, JAIN V, JOSEPH M, et al. A review on micropropagation culture method[J]. Asian Journal of Pharmaceutical Research and Development, 2020,8(1): 86-93.
[8] WHITE T J, BRUNS T, LEE S, et al. Amplification and irect sequencing of fungal ribosomal RNA genes for phylogenetics[M]//PCR Protocols: A guide to methods and applications. New York, America: Academic Press Inc, 1990: 315-322.
[9] LEIFERT C, RITCHIE J Y, WAITES W M. Contaminants of plant-tissue and cell cultures[J]. World Journal of Microbiology and Biotechnology, 1991, 7(4): 452-469.
[10] 谢颖苑,廖飞扬,李天卓,等.桉树组培厂季节性污染及防控[J].绿色科技,2015(1): 34-35.
[11] LAU H T, FARYNA J, TRIPLETT E W. Aquitalea magnusonii gen. nov., sp. nov., a novel Gram-negative bacterium isolated from a humic lake[J]. International Journal of Systematic and Evolutionary Microbiology, 2006, 56(4): 867-871.
[12] MEZZATESTA M L, GONA F, STEFANI S. Enterobacter cloacae complex: clinical impact and emerging antibiotic resistance[J]. Future Microbiology, 2012, 7(7): 887-902.
[13] KUMAR P, KAUSAR M A, SINGH A B, et al. Biological contaminants in the indoor air environment and their impacts on human health[J]. Air Quality, Atmosphere and Health, 2021, 14(11): 1723-1736.
[14] LUCAS J A. Plant pathology and plant pathogens[M]. 4th edition.Wiley-Blackwell, 2020.
Investigation and Analysis of Contaminant Microorganisms in Eucalypt Tissue Culture Seedling Production
CAI Huajie1, HUANG Xiangdong1, MA Tianchi1, YI Runhua2, HE Pulin1*
(1. Leizhou Forestry Bureau Co., Ltd of China Forestry Group, Suixi 524348, Guangdong, China;2. College of Coastal Agriculture Sciences, Guangdong Ocean University, Zhanjiang 524088, Guangdong, China)
Species of contaminated bacteria found in eucalypt tissue cultures were investigated by morphological and molecular biology methods in order to potentially reduce the incidence of deleterious contamination of such cultures. In total 50 species representing 25 genera of microorganism were identified from tissue culture bottles, including 7 species from 6 genera of bacteria along with 43 species from 19 genera of fungi, equating to 14.0% of contaminants being bacteria and 86.0% being fungi. Eucalypt subculture seedlings were mainly contaminated byalong withspp. andspp. each with 5 species. Later after transplanting, it was found that eucalypt explants were contaminated by 21 species representing 11 genera of fungi and 1 species of bacteria.
eucalypt tissue culture; pollutant bacteria; tissue culture seedlings; morphological; amplicon
10.13987/j.cnki.askj.2023.04.005
Q93-331
A
蔡华杰(1972— ),男,政工师,从事森林资源培育。E-mail: 2259787650@qq.com
何普林(1974— ),男,大学本科,林业高级工程师,从事森林资源培育。E-mail: honglin923@sohu.com
猜你喜欢
中国森林病虫(2021年6期)2021-12-20
高师理科学刊(2020年1期)2020-11-26
中国森林病虫(2018年4期)2018-09-19
现代园艺(2018年3期)2018-02-10
现代园艺(2017年13期)2018-01-19
山东林业科技(2017年5期)2017-04-05
西南农业学报(2016年6期)2016-04-16
塔里木大学学报(2015年1期)2015-04-25
大豆科技(2014年5期)2014-03-23
新疆农垦科技(2014年6期)2014-02-28
