
干旱胁迫对翅茎白粉藤光合作用及叶绿体超微结构的影响
2024-01-09 05:35李进华麦生威吴国文张广照
广西林业科学 2023年6期
李进华,麦生威,李 冰,赵 伟,吴国文,张广照,唐 庆
(1.广西壮族自治区林业科学研究院,广西南宁 530002;2.广西新祥高速公路有限公司,广西崇左 532200;3.广西交通投资集团崇左高速公路运营有限公司,广西崇左 532200)
翅茎白粉藤(Cissushexangularis)为葡萄科(Vitaceae)白藤属木质藤本,为热带及亚热带地区特色药用植物,分布于广东、海南和广西等地,生于山地疏林中;其能祛风活络、散瘀活血,可用于缓解风湿痹痛、腰肌劳损、跌打损伤等[1-2]。其根系发达,固土能力强,是边坡绿化中优选的植物;其还可以涵养水源,在一定程度上预防和减少水土流失、山体滑坡等现象。
近年来,全球气候变暖,导致季节性降水分布不平衡,全国各地出现异常气候现象,干旱现象频繁发生[3]。干旱对植物的生长、光合作用等产生不良影响,影响植物的生长发育[4]。光合作用是植物生长和发育的关键过程,受光照、温度、水分和热量等多种因素影响。植物光合速率日变化与植物的生长和环境适应密切相关[5-6]。植物光合色素包括叶绿素和类胡萝卜素,其中叶绿素主要负责吸收、传递和转化光能,类胡萝卜素负责光能捕获和光破坏防御,两者在很大程度上决定植物光合能力的强弱[7-8]。叶绿体对干旱胁迫较敏感,表现为形态结构、大小和数目均会随植物所处环境的变化而变化[9]。目前,关于翅茎白粉藤的研究主要集中在有效成分与药理学[10-15]、苗木繁殖[16]等方面,在干旱胁迫方面的研究鲜见报道。本研究以翅茎白粉藤为试验材料,采用盆栽控水法,探讨干旱胁迫对翅茎白粉藤光合作用和叶绿体超微结构的影响,为其栽培提供参考。
1 材料与方法
1.1 试验地概况
试验地位于广西壮族自治区林业科学研究院园林花卉研究所(108°21′E,22°56′N),属南亚热带季风气候,年均气温20~21 ℃,≥10 ℃年均积温7 206 ℃,年均降水量1 350 mm,一般年份有2~4天轻霜;5—9月为雨季,月均降水量为100 mm 以上;10月—翌年4月雨量较少,月均降水量为80 mm以下[17]。
1.2 试验材料
供试材料为2年生翅茎白粉藤,选择长势一致、具有9~10张叶片、根系发达、无病虫害且株高145.0~148.0 cm的优良植株。
1.3 试验方法
2022年6月10日—8月10日,在广西林科院园林花卉研究所基地的高透光遮雨棚下,采用盆栽法进行试验。每处理选择30盆翅茎白粉藤,在胁迫前充分浇透水肥,使土壤含水量及营养基本保持一致,待土壤自然晾干后,进行干旱胁迫。参照Hsiao[18]的方法,设正常供水(CK)、中度干旱(MD)和重度干旱(SD)3 个水分处理,即土壤含水量分别控制在田间持水量的(70 ± 5)%、(50 ± 5)%和(35 ±5)%。每天早上8:00、中午12:00 和下午18:00,采用WET-2 土壤水分温度电导率速测仪(Delta-T,北京博伦经纬科技发展有限公司)测量土壤含水量,水分不足时用有刻度的烧杯进行补水,并记录加水量。处理60天后,分别测定形态指标、光合指标,观察叶片叶绿体超微结构。
1.4 测定指标及方法
1.4.1 形态指标
2022年6月10日和8月10日分别测定株高、地径、叶片数、叶长和叶宽及根系生物量(鲜重)和地上部分生物量(鲜重)等指标。
1.4.2 光合参数测定
2022年8月10日,在天气晴朗的条件下进行测定。测定时间为8:30~18:30,每隔2 h 测定1 次,每处理测定3 盆,每盆按东、南、西和北方向随机选取3~5 片成熟叶测定,采用Li-6400XT 便携式光合仪(LI-COR)测定叶片的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)及蒸腾速率(Tr)等生理指标,计算水分利用效率(WUE),计算公式为:
1.4.3 叶绿素和类胡萝卜素含量测定
2022年8月10日,取0.3 g 新鲜样品,放入研钵中,加入少量石英砂和碳酸钙粉及95%乙醇2~3 mL,研磨成匀浆,再加95%乙醇5 mL 继续研磨至组织变白,静置3~5 min 后,上清转移定容至25 mL,备用。将提取液稀释至合适浓度后,倒入1 cm 光径比色皿中,以95%乙醇为空白,在波长665、649 和470 nm 下测定吸光度。计算叶绿素a(Ca,mg/L)、叶绿素b(Cb,mg/L)和类胡萝卜素(Cx.c,mg/L)浓度;计算公式[19]为:
式中,Vt为提取液总体积(mL);FW 为鲜重(g);n为稀释倍数。
1.4.4 叶绿体超微结构观察
2022年8月10日,以测定光合特性的叶片为材料,按要求截取2 mm×2 mm 叶片样品,置于2.5%戊二醛中固定过夜,采用0.1 mol/L 磷酸漂洗液漂洗3 次,每次15 min,将样品放入1%锇酸溶液,在4 ℃冰箱中固定2 h。在0~4 ℃环境下,采用50%、70%和90%乙醇逐级脱水,每次脱水时间为15 min;采用100%乙醇和100%环氧丙烷分别脱水5 min,3 次重复;采用环氧丙烷+包埋液(V∶V=2∶1)包埋2 h,再采用环氧丙烷+ 包埋液(V∶V=2∶1)包埋3 h,纯包埋液浸泡过夜,最后采用纯包埋液在室温下浸泡3~4 h。将样品块置于包埋模板内,加入纯包埋剂后放入恒温烘箱内聚合,第1 次烘烤温度和时间为37 ℃和12 h,第2次烘烤温度和时间为45 ℃和12 h,第3 次烘烤温度和时间为60 ℃和48 h,半薄(1 μm)定位,采用超薄切片(70 nm)机切片,采用3%醋酸铀-枸橼酸铅双染色,采用透射电镜观察,拍片。
1.5 数据处理
采用Excel 软件进行数据整理和图表绘制;采用SPSS 7.05 软件进行单因素方差分析(One-way ANOVA)和多重比较分析。
2 结果与分析
2.1 干旱胁迫对翅茎白粉藤生物量的影响
MD 和SD 处理下,胁迫后,株高增加,SD 处理株高最高(187.20 cm),比胁迫前增加38.40 cm;地上部分鲜重、地下部分鲜重均增加,MD 处理的地上部分鲜重和地下部分鲜重均最重(262.00 和43.70 g),比胁迫前分别增加191.70 和25.00 g;叶片数增加,MD处理的叶片数最多(64张),比胁迫前增加18张;地径、叶长、叶宽和根冠比均降低(表1)。随干旱胁迫程度增加,株高增量显著增加(P<0.05);地上部分鲜重和地下部分鲜重增量均呈先增后降的趋势,

表1 翅茎白粉藤干旱胁迫前后的相关生物量变化Tab.1 Biomass changes of C.hexangularis before and after drought stress
MD 处理均显著高于其他处理(P<0.05);地径、叶长、叶宽和叶片数增量均呈降低趋势;根冠比增量呈增加趋势。
2.2 干旱胁迫对翅茎白粉藤光合参数日变化和日均值的影响
CK、MD 和SD 处理的净光合速率和气孔导度日变化曲线均为双峰形,均出现光合“午休”现象;CK的净光合速率和气孔导度日变化曲线峰值均出现在10:30和16:30,MD和SD处理的峰值均出现在12:30和16:30(图1a~b)。MD和SD处理的胞间CO2浓度日变化曲线均为双峰形,峰值均出现在10:30和16:30;CK日变化曲线为单峰形,峰值出现在16:30(图1c)。CK 的蒸腾速率日变化曲线为单峰形,峰值出现在10:30;MD 处理为单峰形,峰值出现在14:30;SD 处理没有明显的变化(图1d);CK 和SD 处理的水分利用效率日变化曲线均为单峰形,峰值出现在16:30;MD 处理为双峰形,峰值出现在12:30 和16:30(图1e)。
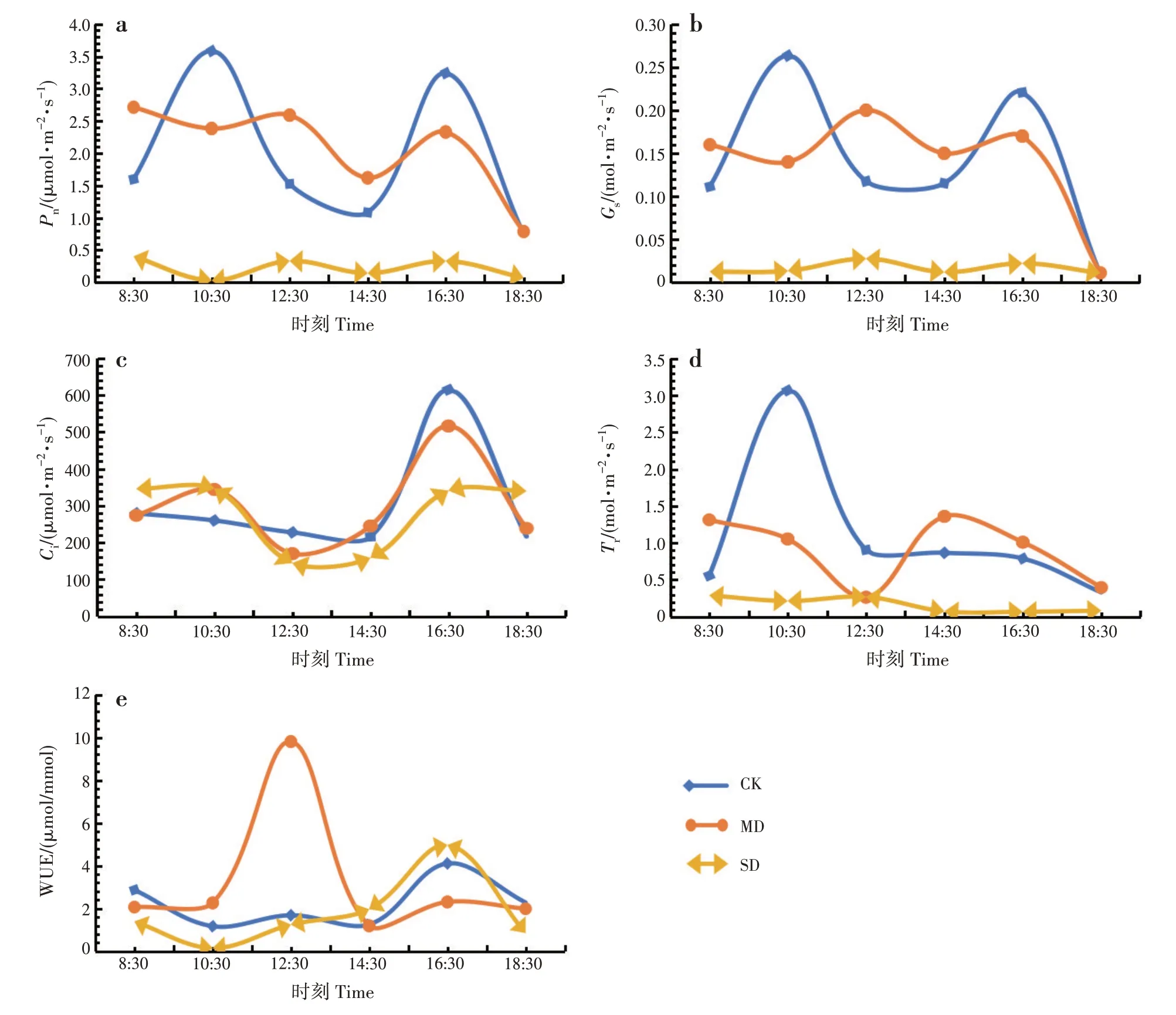
图1 干旱胁迫对翅茎白粉藤叶片光合参数日变化的影响Fig.1 Effects of drought stress on diurnal variations of photosynthetic parameters of C.hexangularis leaves
净光合速率、气孔导度、胞间CO2浓度、蒸腾速率和水分利用效率日均值均差异显著(P<0.05)(表2)。随干旱胁迫程度增加,净光合速率和水分利用率日均值均呈先升后降的趋势,均表现为MD处理>CK>SD 处理;气孔导度、胞间CO2浓度和蒸腾速率日均值均减小,均表现为CK>MD 处理>SD处理。

表2 干旱胁迫对翅茎白粉藤叶片光合作用参数日均值的影响Tab.2 Effects of drought stress on daily mean values of photosynthetic parameters of C.hexangularis leaves
2.3 干旱胁迫对翅茎白粉藤叶片光合色素的影响
叶绿素a 含量、叶绿素b 含量、类胡萝卜素含量和叶绿素总量均差异显著(P<0.05)(表3)。随干旱胁迫程度增加,叶绿素a含量、叶绿素b含量、类胡萝卜素含量和叶绿素总量均呈先增后降的趋势,均在MD 处理下最高,分别为0.722、0.344、0.140 和1.066 mg/g。

表3 干旱胁迫对翅茎白粉藤叶片光合色素的影响Tab.3 Effects of drought stress on photosynthetic pigments of C.hexangularis leaves (mg/g)
2.4 干旱胁迫对翅茎白粉藤叶片叶绿体超微结构的影响
CK 的叶绿体呈长椭圆形,球体比较饱满,类囊体片层整齐、层数多、排列紧密、淀粉粒饱满及叶绿体被膜清晰;MD 处理的叶绿体质壁分离,类囊体结构模糊,排列不整齐,线粒体肿胀;SD处理的叶绿体外膜部分破裂,基粒扭曲变形,基质排列紊乱(图2a~c)。不同干旱胁迫程度均对叶绿体的结构造成一定程度损坏,受损程度随干旱胁迫增加而增加。

图2 干旱胁迫对翅茎白粉藤叶绿体超微结构的影响Fig.2 Effects of drought stress on chloroplast ultrastructures of C.hexangularis
3 讨论与结论
受干旱胁迫时,植物会产生抗旱性,生长发育特征会发生变化,表现为根系发达而深扎,根冠比变大,叶片细胞变小,叶脉致密,单位面积气孔数目增多[20]。本研究中,中度和重度干旱胁迫下,胁迫后株高增加,地径、叶长和叶宽降低,可能是植物通过调节叶肉细胞水分来抵抗干旱胁迫造成的损伤[21]。胁迫后,地上部分鲜重、地下部分鲜重和叶片数均增加;随干旱胁迫程度增加,地上部分鲜重和地下部分鲜重增量呈先增后降的趋势,叶片数增量呈减少趋势,根冠比增量呈增加趋势。表明中度干旱胁迫时,翅茎白粉藤通过扩大水分吸收范围,增加地上部分鲜重、地下部分鲜重和叶片数;重度干旱胁迫时,植物利用有限的水分维持生命活动,降低生物量,使叶片水分含量与土壤可利用水分平衡。
本研究中,CK、中度和重度干旱胁迫下,翅茎白粉藤叶片净光合速率、气孔导度日变化曲线均为双峰形,出现光合“午休”现象。植物光合作用的抑制因素包括气孔限制和非气孔限制,分别由气孔关闭和光合器官自身活性下降引起。干旱胁迫下,净光合速率上升,气孔导度降低,胞间CO2浓度降低,说明气孔和非气孔因素共同限制净光合速率;净光合速率降低,气孔导度降低,胞间CO2浓度降低,说明气孔因素限制净光合速率;净光合速率降低,气孔导度降低,胞间CO2浓度升高,说明非气孔因素限制净光合速率[22]。本研究中,中度干旱胁迫时,净光合速率和水分利用率均上升,气孔导度、胞间CO2浓度和蒸腾速率均降低,说明气孔和非气孔因素共同限制净光合速率;重度干旱胁迫时,净光合速率、气孔导度、胞间CO2浓度、蒸腾速率和水分利用率均降低,说明气孔因素限制净光合速率。
光合色素含量直接影响叶片的光合能力[23]。本研究中,翅茎白粉藤的叶绿素和类胡萝卜素含量均呈先升后降的趋势;中度干旱胁迫下,叶绿素和类胡萝卜素含量均达到最大值,且显著高于对照和重度干旱胁迫。说明中度干旱胁迫有利于翅茎白粉藤的光合作用。
植物细胞的超微结构变化规律是判断其抗逆能力的重要指标之一[24]。受干旱胁迫时,植物的叶绿体会遭受不同程度的损伤[23]。本研究中,中度和重度干旱胁迫下,叶绿体的结构遭到一定程度破坏,导致光合作用减弱,原因可能是干旱胁迫导致内膜不饱和脂肪酸减少,引起膜蛋白的不稳定和膜结构功能的丧失。
综上所述,翅茎白粉藤在中度干旱胁迫下,净光合速率和水分利用率较高,虽然对叶绿体的结构造成一定程度的损坏,但未见植物死亡,可见翅茎白粉藤具有一定的抗旱特征,这一研究结果可为矿山修复、高速公路石质边坡绿化及屋顶绿化等恶劣的环境选择植物提供科学依据。本研究中,翅茎白粉藤的种植基质只采用了1 种混合基质,为了使翅茎白粉藤能在不同地区生长,下一步将开展不同基质、不同光照条件下的胁迫研究。
利益冲突:所有作者声明无利益冲突。
作者贡献声明:李进华负责试验设计和论文撰写;麦生威、李冰和赵伟负责野外种源收集;吴国文负责数据测定与整理;张光照负责数据测定;唐庆负责文献检索和论文校对。
猜你喜欢
今日农业(2021年19期)2022-01-12
湖北农业科学(2021年13期)2021-08-04
新疆农业科技(2021年2期)2021-07-03
中学生物学(2020年12期)2020-03-30
新农业(2017年2期)2017-11-06
广西林业科学(2016年1期)2016-03-20
生物灾害科学(2015年3期)2015-03-11
生物技术通报(2014年10期)2014-03-21
食品科学(2013年6期)2013-03-11
中国烟草学报(2012年2期)2012-04-09
