
塔里木河中、下游胡杨林种内竞争对比研究*
2024-01-30 01:46刘建莹玉米提哈力克何熙祥木扎帕尔米吉提
新疆大学学报(自然科学版)(中英文) 2024年1期
刘建莹,玉米提·哈力克,何熙祥,木扎帕尔·米吉提
(新疆大学生态与环境学院绿洲生态教育部重点实验室,新疆 乌鲁木齐 830017)
0 引言
物种竞争一直是生态学的核心问题,研究植物间的竞争对了解植物群落组成以及演替具有重要影响,可进一步指导并促进森林生态系统功能的完善[1].树木因对资源的争夺而产生的相互作用关系称为林木竞争[2].树木间的竞争是森林生态系统中的普遍现象,其结果是一个有机体影响另一个有机体的生长和发育.
竞争在形式上表现为树木个体生长与生存空间的关系,而实质上是反映树木对环境资源所承受的竞争压力.随着树木的生长发育,其对空间以及资源的需求逐渐增大,树木之间开始竞争,使得部分树木逐渐死亡,其余部分树木则占据更大的生存空间[3].树木本身生长状态(如胸径、树高、冠幅等)、立地条件及相邻木状态均对树木竞争强度有显著影响[4].一般认为,当树木树冠或根系发生重叠时树木之间才会产生竞争,竞争压力主要来源于周围一定距离内的树木.当树木距离较远时,树木的竞争作用减弱或消失[5].
胡杨是杨柳科、杨属的一种落叶乔木,是塔里木河流域荒漠河岸植被主要优势种[6].胡杨对区域生态系统的良性发展十分关健,如水土保持、水源涵养和生物多样性保育等[7-8].胡杨的种群分布格局决定着塔里木河流域植物群落结构[9].前人对树木种内竞争等方面进行了较多研究.潘磊磊等认为沙地樟子松竞争半径为8 m,且胸径达到20 cm以上时,对象木竞争强度趋于稳定并维持在较低水平[10].周禧琳等认为生境的改变是影响胡杨种内竞争强度变化的主要外在因素[11].目前,关于胡杨竞争强度及其对区域水条件响应的研究还比较欠缺,不同水分梯度下胡杨种内竞争机制方面缺乏深入认识.另外,胡杨种内竞争研究主要集中在单一区域,不同区域胡杨种内竞争对比研究较少.鉴于此,本文对比分析塔里木河中、下游不同区域胡杨种内竞争随水分梯度的变化规律,以期为塔里木河下游胡杨种群的更新、复壮提供理论参考.
1 研究区概况
研究区位于塔里木河中游英巴扎至恰拉之间的河段(41°03′~41°18′N,84°42′~84°42′E),以及塔里木河下游库鲁克沙漠与塔克拉玛干沙漠之间的冲积平原——阿拉干(40°08′N,88°21′E).研究区气候干旱、降雨稀少、蒸发量大且风沙浮沉天气频发,属于典型的大陆性荒漠气候.特殊的气候及地理条件决定了当地以耐高温、耐盐碱的植被居多,沿河道发育有胡杨、黑刺(Lycium ruthenicum)、多枝柽柳(Tamarix ramosissima)、罗布麻(Poacynum hendersonii)、花花柴(Karelinia caspia)、铃铛刺(Halimodendron halodendron)、芦苇(Phragmites communis)等[12-14],地表流沙活动频繁,植被盖度较低.
2 研究方法
2.1 数据获取
2022年6月至7月,在塔里木河下游阿拉干断面垂直于河道选取3条样带,命名为A、B、C,塔里木河中游轮台县境内垂直于河道选取3条样带,命名为D、E、F,塔里木河下游每条样带内依据水分梯度划分3块50 m×50 m样方,塔里木河中游胡杨林分每条样带内依据水分梯度划分3块30 m×30 m样方,共18个样方.利用手持测高仪、胸径尺、皮尺对样方内所有胡杨进行编号并测定其胸径、树高、枝下高、冠幅等参数,同时人工建立坐标系,测定所有林木的空间位置坐标,胡杨林分基本特征见表1.
2.2 竞争分析
诸多竞争指数中,Hegyi[15]的竞争模型可同时反映种群个体生长与生存空间的关系,以及树木个体对环境资源的需求与真实生境下树木对环境资源占有量之间的关系.且Hegyi的单木竞争模型简单易行,所获得的数据准确性高,已经被大量用于树木竞争关系研究[16-19].因此,本文利用Hegyi竞争指数,探究塔里木河中、下游胡杨种内竞争规律,其公式为:
式中:CI为竞争指数,为平均竞争指数;DBHi为对象木i的胸径,DBHj为竞争木j的胸径;Lij为对象木i与竞争木j之间的距离;N为竞争木的株数,n为对象木株数.CI、值越大,表明对象木承受的竞争压力越大.
式中:CIA为林分平均竞争指数,其值越大,则表明样地内树木受到的竞争压力越大,树木之间的竞争越激烈.
一般认为生态位发生重叠时,由于资源和空间的有限性,进而树木之间发生竞争,所以对象木所受到的竞争压力主要来自周围树木.因此,本文采用固定样圆半径法确定塔里木河中、下游胡杨竞争范围.随半径增加,当平均竞争指数变化不显著时,确定胡杨的竞争范围Lij.
2.3 统计分析
基础数据分析采用Excel 2019和SPSS 26.0完成,Hegyi的单木竞争模型通过软件R4.1.2的spatstat包完成计算,并采用Origin 2022进行绘图.
3 结果与分析
3.1 竞争范围的确定
3.1.1 塔里木河下游胡杨竞争范围
为计算对象木的竞争半径,本文以2 m为样圆半径、2 m为半径增量,分别计算2~20 m范围内对象木的平均竞争指数.如图1所示,A、B、C 3条样带各样方内胡杨平均竞争指数随半径的增加而减小,或随竞争半径增加呈现波动趋势.根据竞争强度变化量可知,随竞争范围的增加,竞争指数变化量整体呈现减小的趋势,其中样方A3、C3中胡杨竞争指数随竞争范围的增加呈现上下波动.当竞争半径为10 m时,竞争指数与竞争指数变化量保持稳定且处于较低水平,表明距离越近竞争木对对象木的影响越明显.故本文认为在塔里木河下游10 m是研究胡杨种内竞争的最适范围.
3.1.2 塔里木河中游胡杨竞争范围
为确定塔里木河中游胡杨竞争范围,本文以2 m为样圆半径、2 m为半径增量,分别计算2~14 m范围内对象木的平均竞争指数(图2).D、E、F 3条样带各样方内胡杨平均竞争指数随半径的增加而逐渐减小,随竞争范围的增加,竞争强度变化量整体呈现减小的趋势.当竞争半径为8 m时,竞争指数与竞争指数变化量保持稳定且处于较低水平,表明距离越近竞争木对对象木的影响越明显.故本文认为在塔里木河中游8 m是研究胡杨种内竞争的最适范围.
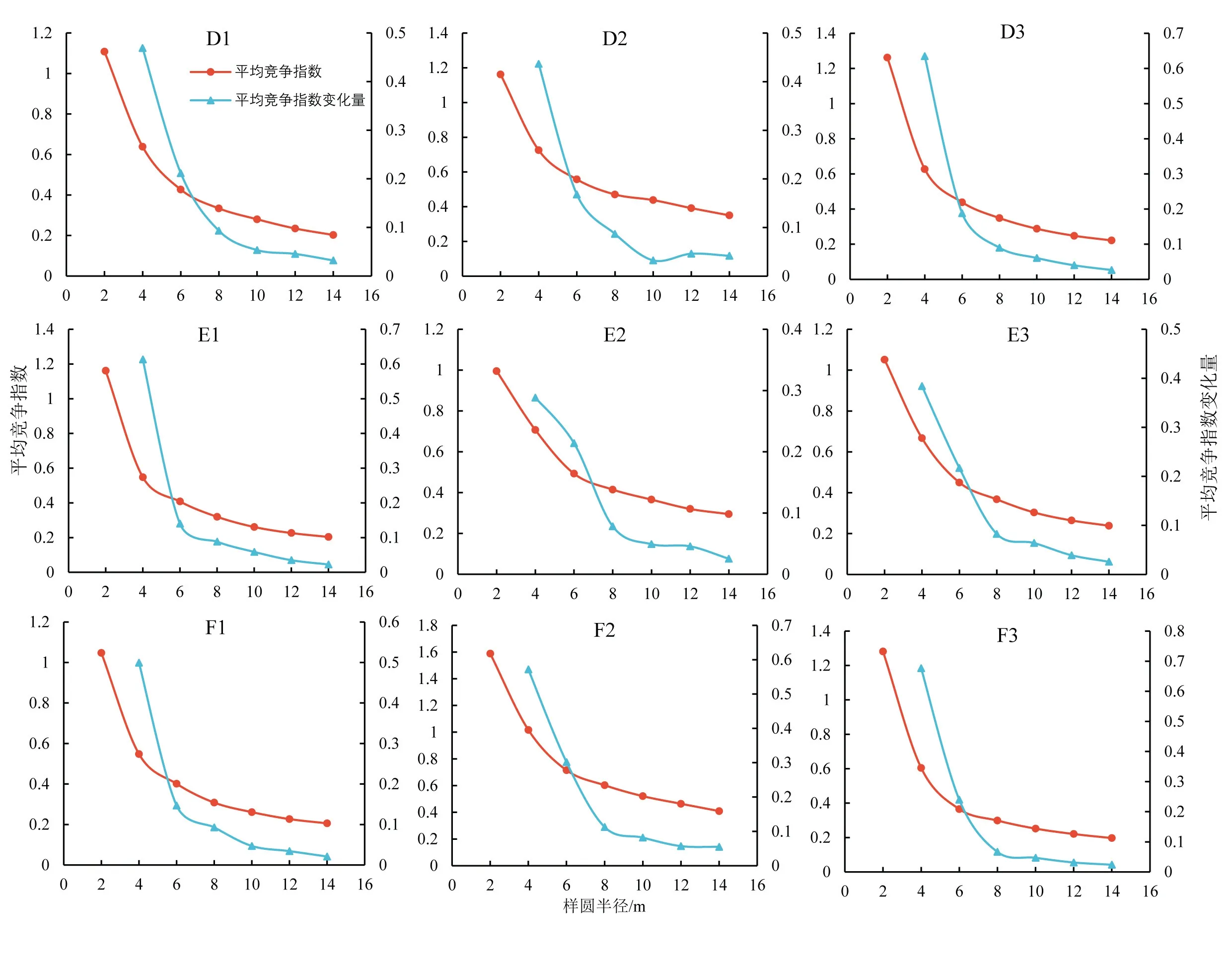
图2 塔里木河中游胡杨竞争强度与样圆半径的关系
3.2 胡杨竞争强度与胸径的关系
本文对塔里木河下游及中游不同水分梯度的胡杨进行回归分析,发现胡杨竞争指数与胸径很好服从幂函数关系.由图3可知,塔里木河流域胡杨整体表现为胸径越大、受到的竞争压力越小,塔里木河下游胡杨胸径达到20 cm以上、中游胡杨胸径达到30 cm以上时,胡杨的竞争指数逐渐稳定,并维持在较低水平.相比之下,塔里木河下游胡杨胸径对竞争指数解释度高于中游胡杨(图3、图4).这表明在塔里木河下游,单木尺度上胡杨胸径更能体现其竞争压力的大小.在塔里木河下游,随着水分梯度的降低,胡杨胸径对竞争指数的解释度逐渐降低;与之相反,在塔里木河中游,随着水分梯度的降低,胡杨胸径对竞争指数的解释度逐渐增加.这可能是塔里木河中、下游水环境差异所导致的.

图3 塔里木河下游胡杨竞争指数与胸径的散点图

图4 塔里木河中游胡杨竞争指数与胸径的散点图
3.3 水分梯度与竞争强度的关系
在塔里木河下游,随着水分梯度的降低,胡杨竞争强度表现出下降的趋势(图5).样带A中,胡杨竞争强度随着水分梯度的降低,呈现先升高后降低的趋势,且存在显著差异(P<0.05).样带B中,从B1到B2胡杨竞争强度下降但差异不显著(P>0.05),至样方B3胡杨竞争指数出现显著下降(P<0.05).样带C中,从C1到C2胡杨竞争强度下降但差异不显著(P>0.05),至样方C3胡杨竞争指数出现显著下降(P<0.05).
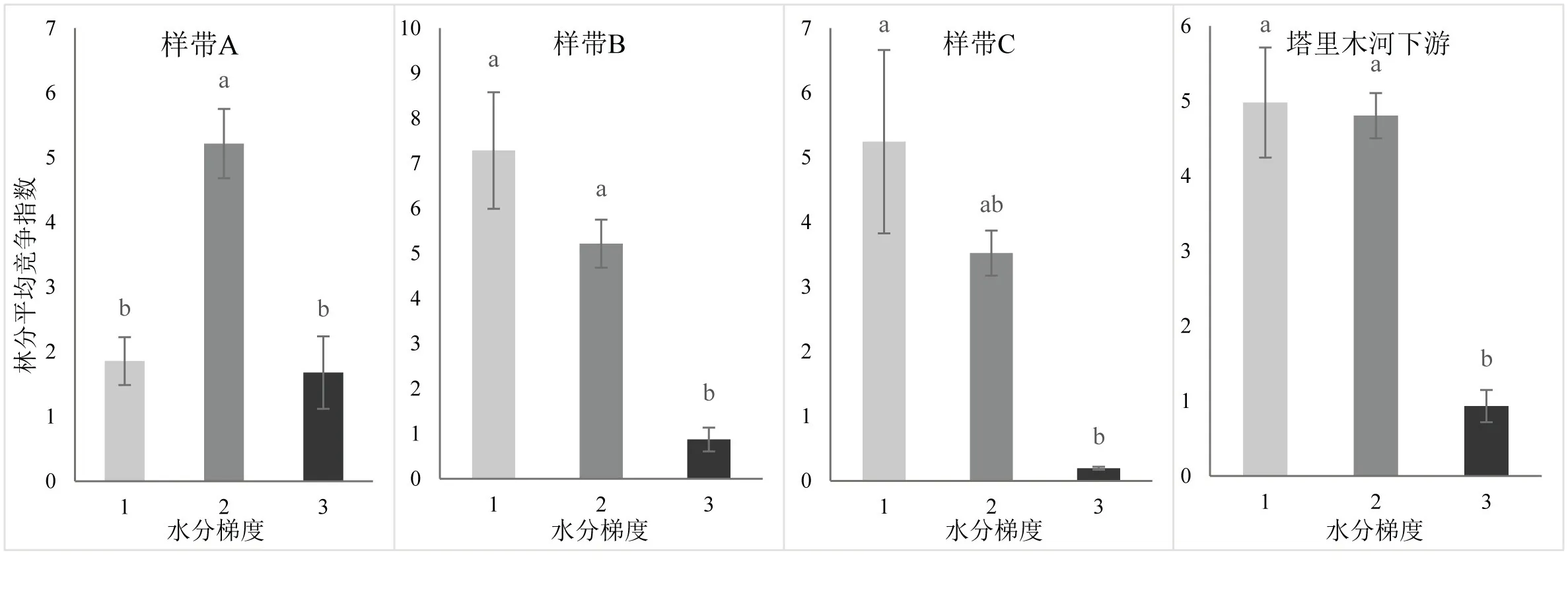
图5 塔里木河下游水分梯度与竞争指数条形图
通过分析塔里木河中游水分梯度与胡杨竞争指数,发现随着水分梯度的降低,胡杨竞争强度表现出整体下降的趋势,但差异不显著(P>0.05)(图6).样带D中,随着水分梯度的降低,胡杨竞争强度整体表现为先降低后升高的趋势,但差异不显著(P>0.05).样带E中,胡杨竞争强度随水分梯度的降低整体呈现下降的趋势,但差异不显著(P>0.05).样带F中,从F1到F2胡杨竞争强度下降,但差异不显著(P>0.05).至样方F3胡杨竞争强度出现显著下降(P<0.05).
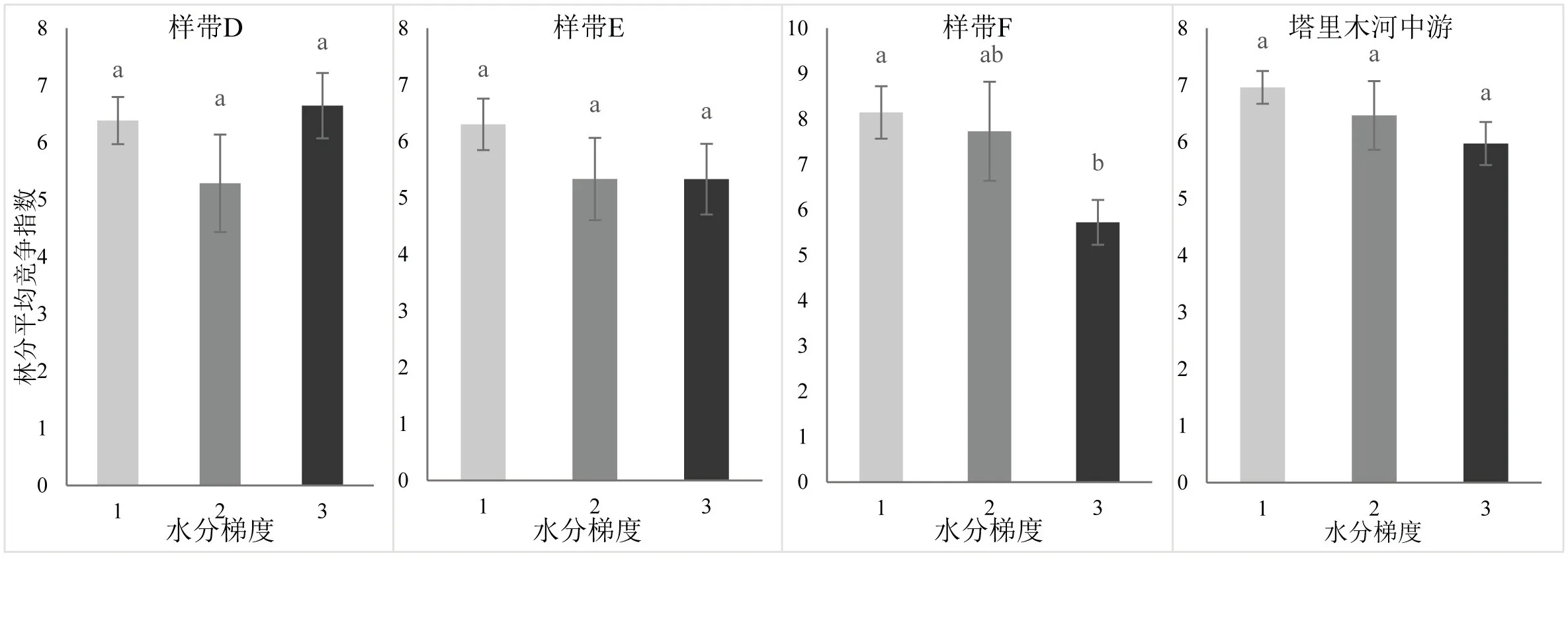
图6 塔里木河中游水分梯度与竞争指数条形图
通过独立样本T检验对比塔里木河中游及下游胡杨竞争强度,结果表明:不同水分梯度下塔里木河中游胡杨竞争指数均高于下游,达到显著水平(P<0.05)(图7),且随着水分梯度的降低,显著性逐渐加强.这可能与不同地域水环境差异及其林分密度有关.
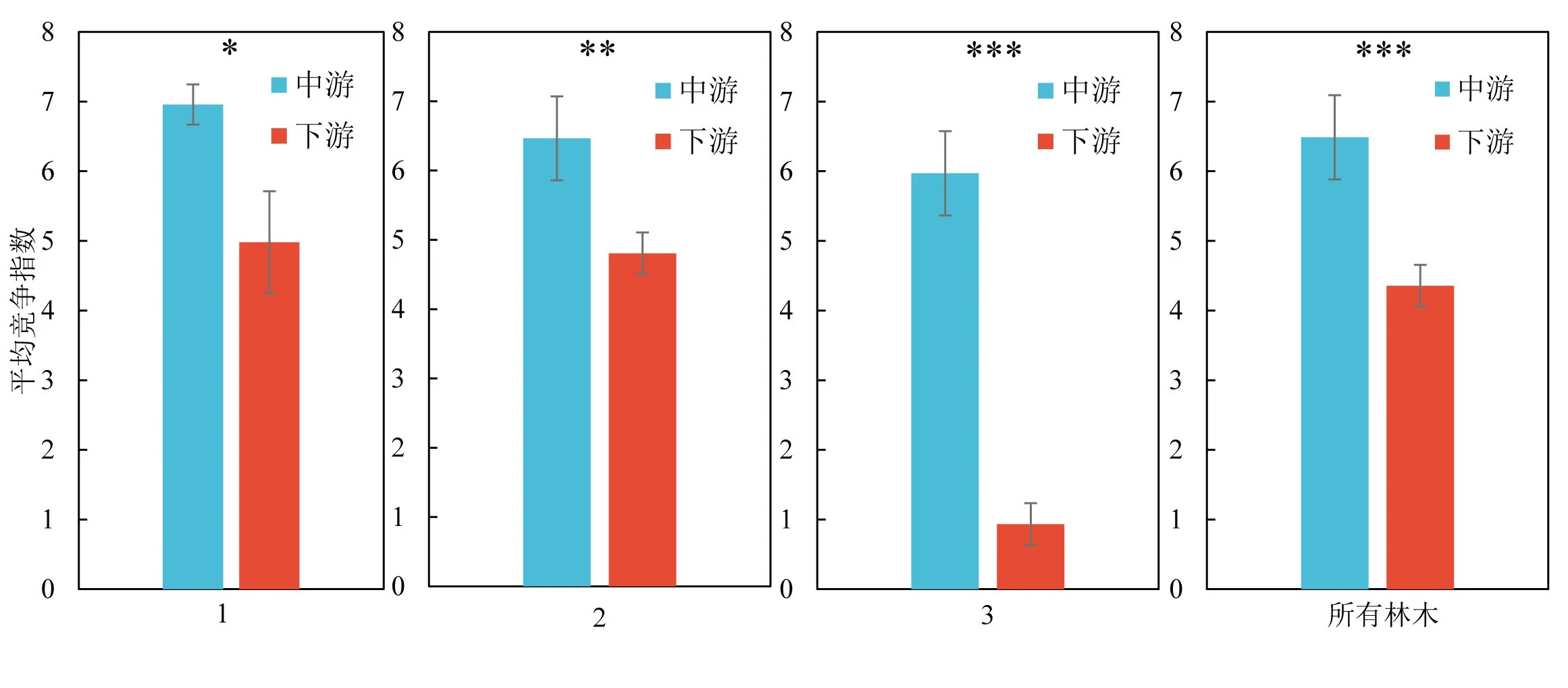
图7 塔里木河中、下游水分梯度与竞争指数条形图
4 讨论
有研究表明树木个体之间的竞争主要发生在相邻树木个体之间[20].因此,本文采用固定样圆法确定塔里木河下游及中游胡杨竞争半径[21],发现塔里木河下游竞争范围介于2~10 m,竞争指数整体呈下降趋势,当超过10 m时胡杨竞争指数保持在稳定范围.塔里木河中游竞争范围介于2~8 m,竞争指数整体呈下降趋势,当超过8 m时胡杨竞争指数保持在稳定范围.由此得出塔里木河下游胡杨竞争范围为10 m、中游胡杨竞争范围为8 m,出现差异的原因可能与研究区域胡杨所处生境有关.相较塔里木河中游,区域水资源与养分资源更为匮乏,胡杨根系延伸范围相较中游更大,需要更大的面积以获取生长、生存所需资源,故而塔里木河下游胡杨竞争范围大于中游.
树木间的竞争受内在因素(个体大小、生长阶段等)与环境因素(地形、气候、水环境等)共同作用的影响[22].Hegyi的单木竞争模型是衡量树木个体竞争强度常用的竞争指标,能够间接反映植物个体对资源和环境的现实分配[23].树木大小对其竞争强弱存在较大的影响,本文通过拟合胡杨胸径与其竞争指数的关系,发现胡杨竞争指数随其胸径的增加而减小,符合物种竞争的一般规律[24].研究发现,塔里木河下游胡杨胸径低于20 cm时,其竞争指数随胸径变化显著;当胸径大于20 cm时,对象木竞争强度趋于稳定.塔里木河中游胡杨胸径低于30 cm时,胡杨竞争强度随胸径变化显著;当胸径大于30 cm时,对象木竞争强度趋于稳定.出现差异的原因可能与塔里木河中、下游区域不同的水环境有关.
树木之间的竞争意味着在有限的资源条件下、不足以维持两株或多株树木在同一空间范围内充分生长,故而竞争是影响其生长发育的重要因素,其主要表现为密度效应[25].树木之间的竞争主要发生在邻近木之间,包括对地上部分及地下部分的竞争[20].本文通过划分水分梯度,分析塔里木河中、下游不同水分梯度下胡杨竞争指数的变化,发现胡杨林平均竞争指数随着水分梯度的降低而逐渐减小,这可能是由于随土壤水分降低,水资源情况不足以支撑数量较多的胡杨同时生存,在自然稀疏规律的作用下,部分胡杨死亡,林分密度逐渐降低[26],从而导致胡杨林平均竞争指数降低.值得注意的是,并非所有样带胡杨竞争指数随水分梯度降低而降低,林分密度也并非随着水分梯度降低而降低,而是呈现林分密度随水分梯度降低先增加后降低或先降低后增加的结果(表1、图5、图6),造成其竞争强度变化的原因可能与小尺度下胡杨聚集程度相关.生境的改变是影响胡杨种内竞争强度变化的主要外在因素[11],本文发现不同水分梯度下塔里木河中游胡杨竞争指数均显著高于下游;另外,塔里木河中游胡杨竞争压力大于下游,且随着水分梯度的降低竞争强度下降幅度较小.可能是由于塔里木河中游水资源条件优于下游,故而导致中游胡杨林分密度更大,同时树木个体相对较大(表1).
5 结论
本文利用Hegyi单木竞争模型等分析方法,探究了塔里木河中、下游不同水分梯度下胡杨林种内竞争变化规律.结果表明:1)塔里木河下游胡杨的最适竞争范围为10 m,中游胡杨最适竞争范围为8 m.2)胡杨竞争指数与胸径服从幂函数关系(P<0.001),胡杨竞争指数随对象木胸径的增加而逐渐减小;塔里木河下游对象木胸径达到20 cm以上、中游达到30 cm以上时,其竞争指数逐渐稳定,并维持在较低水平.3)随着水分梯度的降低,胡杨林分竞争指数呈现下降趋势,且塔里木河中游胡杨竞争压力高于下游.未来在塔里木河下游胡杨更新、复壮过程中,应当充分考虑水分对胡杨种内竞争的影响,采取“线-面”结合的生态输水方式,以促进塔里木河流域胡杨的恢复与保护.
猜你喜欢
金桥(2021年11期)2021-11-20
北广人物(2020年47期)2020-12-09
意林绘阅读(2019年2期)2019-03-05
下一代英才(2018年12期)2018-02-20
创新作文(1-2年级)(2018年6期)2018-01-22
甘肃林业(2016年4期)2016-11-07
生态学报(2016年12期)2016-08-09
中国水土保持科学(2016年3期)2016-07-19
生态学报(2016年1期)2016-03-10
吐鲁番(2014年2期)2014-02-28
