
旱作区农田土壤AM 真菌对长期有机无机施肥的响应*
2024-02-20 08:07田霞,王媛,张雨,郭蓉,马琨
中国生态农业学报(中英文) 2024年2期
田 霞,王 媛,张 雨,郭 蓉,马 琨
(宁夏大学西北退化生态系统恢复与重建教育部重点实验室/宁夏大学西北土地退化与生态恢复国家重点实验室培育基地 银川 750021)
丛枝菌根(AM)真菌可与自然界中绝大部分植物形成共生关系[1],在促进土壤物质循环、改善土壤结构[2]、提升土壤质量[3]等方面发挥重要作用。然而,AM 真菌对土壤质量的影响却取决于其群落组成[4]。因此,了解AM 真菌群落动态变化对于长期有效的土壤生态系统管理非常重要。
施肥在保持和改善土壤肥力的同时影响了土壤AM 真菌群落组成和多样性。已有研究报道了无机肥施用对土壤AM 真菌群落的影响。如,Chen 等[5]、Zeng 等[6]研究认为,施用化学氮、磷肥对土壤AM真菌群落多样性有负面影响;氮肥施用对AM 真菌群落的多样性(Shannon 指数)产生了负效应,受无机氮肥施用影响,土壤pH、硝态氮和速效磷的变化是驱动AM 真菌群落变化的主要因素[6]。Liu 等[7]也发现,施用无机肥会降低高寒草甸生态系统下垂穗披碱草(Elymus nutansGriseb.)草地土壤AM 真菌群落多样性并改变群落组成;长期施用无机肥会导致黑土中AM 真菌群落组成减少[8]。综合比较发现,长期施用无机氮、磷肥会对地上植物群落的物种丰富度产生负面影响,也会导致寄主植物对AM 真菌的碳分配减少,进而降低AM 真菌群落多样性。但Cheng 等[9]却发现,长期或短期施用无机肥会显著增加AM 真菌多样性。分析认为,这主要是由于该试验中氮和磷施用一方面显著促进了植物根系发育,使AM 真菌获得更多的碳源,但另一方面导致土壤pH 下降,刺激AM 真菌的菌丝分枝、囊泡形成和定殖,有利于AM 真菌多样性提高。此外,土壤AM 真菌多样性的提高也与90 多年的轮作模式有关。有机肥作为微生物的主要底物和能量来源,也会影响AM 真菌的群落组成[10]。Oehl 等[11]认为,有机肥的施用对AM 真菌多样性有积极影响。长期有机肥施用增加了AM 真菌多样性,但降低了AM 真菌的丰富度[12]。然而,Wang 等[13]却发现,黄淮海平原长期施用有机肥对土壤AM 真菌群落组成和多样性有显著的负面影响。但Qin 等[14]认为,施用有机肥有利于土壤AM 真菌的生长,长期有机肥与无机肥配施却会抑制土壤AM 真菌群落多样性。Ma 等[4]利用中国东北黑土地37 年的长期研究证实,AM 真菌群落多样性降低和升高是由长期无机肥和有机肥配施导致的土壤理化性质变化所驱动。Luo 等[15]针对15年不同有机无机肥配施后玉米(Zea maysL.)-小麦(Triticum aestivumL.)轮作系统中土壤AM 真菌群落研究得出,施肥通过改变表层和底土中速效磷和土壤有机碳水平而导致AM 真菌群落组成产生差异;施肥情况下,作物种类、覆盖作物、农业管理活动和土壤环境都会影响AM 真菌群落结构。因此,无机肥、有机肥以及有机无机肥混施导致土壤AM 真菌多样性升高、降低或不变等不一致的结果说明,AM 真菌对土壤环境变化的响应,应该是施肥类型、施肥方式、土地利用方式、土壤性质、气候条件以及AM 真菌特性等因素综合作用的结果,其对土壤AM 真菌群落结构组成及多样性变化的驱动机制仍需要进一步研究。
宁夏中部旱作区水资源匮乏、干旱严重、土壤贫瘠肥力较低,土壤培肥对提升土壤生态系统生产力的作用显著。但有关长期有机无机肥配施下,旱作区土壤AM 真菌群落对肥料类型、培肥过程的响应以及有机无机肥配施驱动AM 真菌变化的作用机制仍不明确。因此,为揭示长期有机无机施肥对AM 真菌群落组成和多样性的影响机制,在宁夏中部干旱区进行了为期11 年的施肥研究,旨在阐明旱作区土壤AM 真菌群落变化与土壤环境因子间的相互关系,揭示AM 真菌多样性及组成对培肥管理的响应规律,以期为宁夏中部旱作区合理施肥管理提供理论依据。
1 材料与方法
1.1 研究区概况
试验地位于宁夏同心县王团镇旱作节水示范区(36°51′N,105°59′E),该区域地处宁夏中部干旱带,属典型大陆性气候。年均降水量272.6 mm,蒸发量2740 mm,年均气温8.7 ℃。土壤质地为灰钙土,试验地为2010 年秋季新推耕地,试验前0~20 cm 土壤基础理化性状为: 全氮0.18 g∙kg−1、全磷0.65 g∙kg−1、有机质4.78 g∙kg−1、碱解氮9.13 mg∙kg−1、速效磷9.32 mg∙kg−1、速效钾97.08 mg∙kg−1、pH 9.20。
1.2 试验设计与样品采集
试验开始于2011 年3 月,2011—2016 年种植模式为马铃薯(Solanum tuberosumL.)—油葵(Helianthus annuusL.)—马铃薯—油葵—油葵—玉米轮作,2017—2021 年均种植玉米。试验采用单因素随机区组设计,4 个处理,3 次重复,小区面积90 m2(6 m×15 m)。处理为不施肥(T0)、单施化肥(T1)、化肥+牛粪有机肥(T2)、化肥+羊粪有机肥(T3)。牛粪有机肥的年施用量为37 500 kg∙hm−2,牛粪(以风干基计)含有机质36.09%、全氮19.09 g∙kg−1、全磷11.06 g∙kg−1、全钾12.28 g∙kg−1、全盐23.12 g∙kg−1,pH 7.62;羊粪施用量为37 500 kg∙hm−2,羊粪(以风干基计)含有机质41.15%、全氮21.93 g∙kg−1、全磷14.33 g∙kg−1、全钾7.72 g∙kg−1、全盐4.69 g∙kg−1,pH 7.62。长期定位试验中,马铃薯和玉米的氮(N)、磷(P2O5)、钾(K2O)肥配比为12∶6∶7,施用量分别为尿素 391.35 kg∙hm−2、重过磷酸钙195.6 kg∙hm−2、硫酸钾210 kg∙hm−2;油葵氮(N)、磷(P2O5)、钾(K2O)肥配比为12∶6∶4,施用量分别为尿素 391.35 kg∙hm−2、重过磷酸钙 195.6 kg∙hm−2、硫酸钾 120 kg∙hm−2。全部的重过磷酸钙、硫酸钾以及30%尿素在春季整地时基施,剩余70%尿素追施。牛粪、羊粪有机肥在秋季土地翻耕整理时做基肥一次性施入。玉米采用宽窄行种植(宽行行距为70 cm,窄行行距40 cm,株距20 cm),栽培管理措施一致。玉米采用滴管方式,各年间补充灌水定额均为1800 m3∙hm−2。年际间的滴灌时间基本相似,分别在5 月23 日、6 月23 日、7 月12 日、8 月2 日左右,玉米主要生育期滴灌补水4 次,每次补水均为450 m3∙hm−2。
2018 年和2021 年,玉米收获期均在10 月3 日,收获前利用5 点采样法采集试验各处理试验小区0~20 cm 耕层土壤样本。为减少处理间的误差,各试验小区均采集独立样本2 个。样品置于4 ℃冰盒保存带回实验室。一部分样品自然风干,用于土壤理化性质测定。另一部分土壤样品过2 mm 筛后,保存在−80 ℃冰箱中,用于土壤AM 真菌多样性测定。
1.3 测定方法
1.3.1 土壤理化性质测定
土壤全氮采用半微量凯式法测定,土壤碱解氮采用碱解扩散法测定,土壤全磷采用HClO4-H2SO4钼锑钪比色法测定,土壤速效磷采用Olsen 法测定,土壤速效钾采用NH4OAc 浸提火焰光度法测定,土壤有机质采用重铬酸钾容量-外加热法测定,土壤pH采用1∶5 水土比测定[16]。
1.3.2 DNA 提取、PCR 扩增和测序
土壤AM 真菌的DNA 提取、扩增参照马琨等[17]的步骤。采用Illumina 公司的TruSeq Nano DNA LT Library Prep Kit 制备测序文库,由上海派森诺生物科技有限公司对AM 真菌群落DNA 片段进行双端测序,首先调用qiimecutadapt trim-paired 切除序列的引物片段,弃去未匹配引物的序列;然后通过qiime dada2 denoise-paired 调用DADA2 进行质控、去噪、拼接、去嵌合体。对于AM 真菌序列选用UNITE 数据库,采用QIIME2 的classify-sklearn 算法,对于每个OTU 的代表序列,在QIIME2 软件中使用默认参数,使用预先训练好的Naive Bayes 分类器进物种注释,获得每个样本中的AM 真菌群落在各分类水平的具体组成。使用QIIME2 软件中的多样性插件,对每个样品的α 指标(包括Chao1 丰富度指数,Shannon 多样性指数)进行估计。在OTU 层面,计算各样本的距离矩阵,并通过多种非监督的排序、聚类手段,结合相应统计学检验方法,衡量不同样本(组)间的β 多样性差异及差异显著性。各处理组间分类学组成的差异分析使用Mothur 软件,调用Metastats 的统计学算法,对基于属水平的各分类单元序列量差异进行比较检验。
1.4 数据处理与统计分析
利用Excel 2019 进行数据整理,采用DPS 7.05软件进行单因素方差分析(One-way ANOVA)和最小显著性差异(LSD 检验,P<0.05)。采用Origin 2021绘制AM 真菌属水平相对丰度分布图。非度量多维尺度(NMDS)用Vegan 和ggplot2 包在R 软件进行可视化。采用Canoco 5.0 软件进行AM 真菌多样性、丰富度及基于属水平上AM 真菌群落组成相对丰度与土壤环境要素间的冗余分析(RDA),采用蒙特卡洛999 次随机无限排列检验,确定土壤因子的显著性影响(P<0.05)。
2 结果与分析
2.1 有机无机肥配施对土壤理化性质的影响
由表1 可见,在长期有机无机肥配施下,土壤全氮、有机质和速效磷含量均显著增加(P<0.05)。与T0 处理相比,培肥7 年后(2018 年),T2、T3 处理的土壤全氮含量显著提高64.00%、56.00%,有机质显著提高58.27%、41.06%,有效磷含量显著提高124.31%、113.90%;培肥10 年后(2021 年),与T0 处理相比,T2、T3 处理的土壤全氮显著增加78.05%、85.37%,有机质显著增加85.86%、100.25%,有效磷含量显著提高117.17%、85.49%。但长期有机无机结合的培肥措施显著降低了土壤pH (P<0.05),较T0处理而言,2018 年T2、T3 处理显著降低2.51%、2.05% (P<0.05),2021 年显著下降1.09%、0.97%。
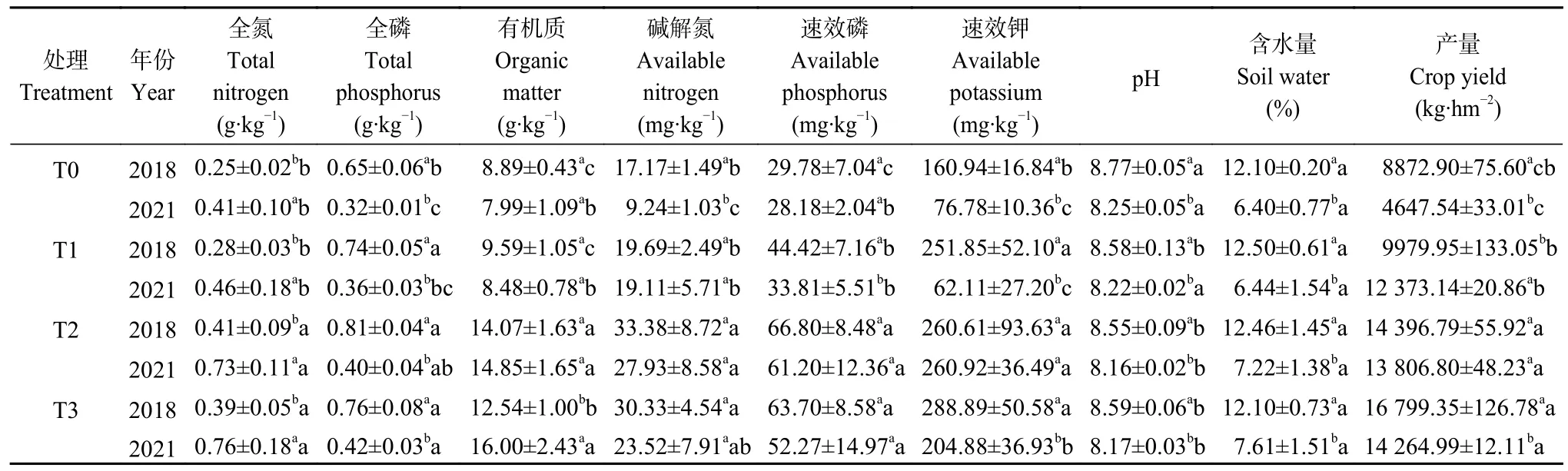
表1 2018 年和2021 年不同有机无机肥配施处理下的土壤理化性质和作物产量Table 1 Soil physicochemical properties and crop yields under different treatments of long-term combined application of organic and inorganic fertilizers in 2018 and 2021
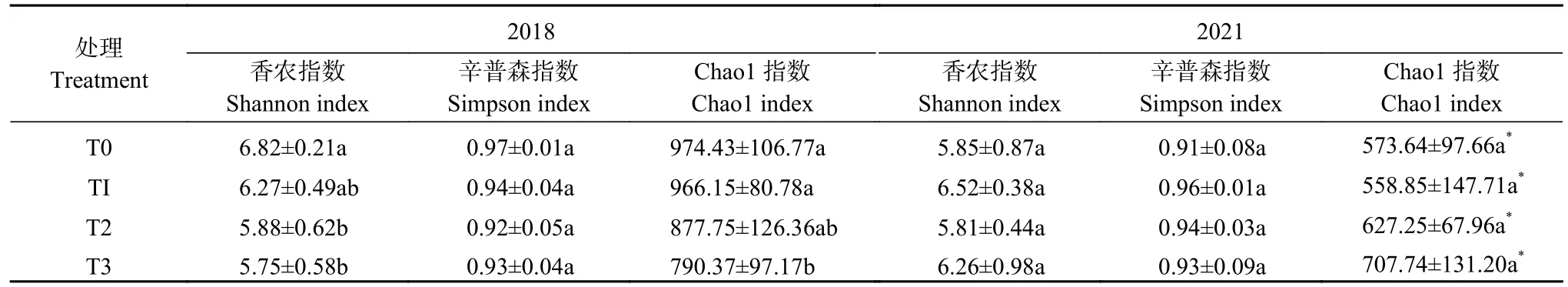
表2 2018 年和2021 年长期有机无机肥配施对土壤丛枝菌根(AM)真菌群落α 多样性的影响Table 2 Effect of long-term combined application of organic and inorganic fertilizers on α diversity of soil arbuscular mycorrhiza(AM) fungal communities in 2018 and 2021
2021 年,化肥配施牛粪有机肥和配施羊粪有机肥处理间对土壤理化性状的影响整体上无明显差异。与2018 年相比,2021 年T0、T1 和T3 处理下土壤全磷、速效钾含量均呈显著下降趋势。其中,土壤全磷含量显著下降50.77%、51.35%和44.77% (P<0.05);速效钾含量显著下降52.29%、75.34%和29.08%(P<0.05)。
有机无机肥配施对作物产量影响较大,但玉米产量对不同施肥措施有不同的响应。整体上,不同施肥处理下2018 年产量高于2021 年产量。在2018年,T3 处理玉米产量最高,其次是T2 处理,与T0 处理相比,T2 和T3 处理分别显著提高62.26%、89.33%(P<0.05),但两处理间无显著差异。2021 年,玉米产量顺序为T3>T2>T1>T0,T1、T2 和T3 较T0 处理分别显著提高166.23%、197.08%和206.94% (P<0.05)。
2.2 有机无机肥配施对土壤AM 真菌群落组成的影响
长期有机无机肥配施下,2018 年和2021 年,土壤AM 真菌由12 属下降到7 属,土壤AM 真菌减少5 属(图1)。其中,无梗囊霉属(Acaulospora)、内养囊霉属(Entrophospora)、巨孢霉属(Gigaspora)、耳孢囊霉属(Otospora)和有隔球囊霉属(Septoglomus)的AM 真菌在2021 年均未发现。
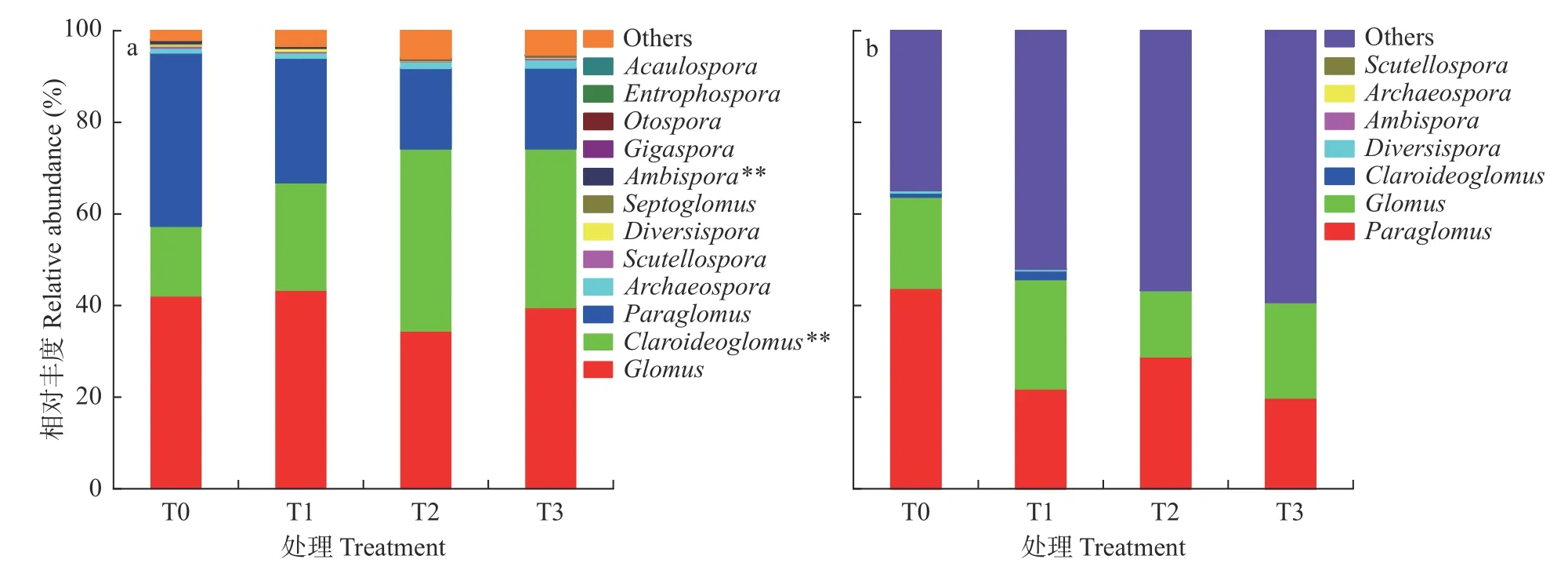
图1 2018 年(a)和2021 年(b)长期有机无机肥配施处理下土壤丛枝菌根(AM)真菌属水平群落组成Fig.1 Community composition of arbuscular mycorrhiza (AM) fungi at genus level in 2018 (a) and 2021 (b) under different treatments of long-term combined application of organic and inorganic fertilizers
连续有机无机肥配施7 年后(2018 年),土壤AM 真菌优势属为球囊霉属(Glomus)、近明球囊霉属(Claroideoglomus)和类球囊霉属(Paraglomus),平均相对丰度分别占总序列的39.83%、28.38%、25.82%。连续有机无机肥配施10 年后(2021 年),AM 真菌优势属仍为球囊霉属和类球囊霉属,平均相对丰度分别达19.78%和28.58%。尽管球囊霉属和类球囊霉属AM 真菌均为长期培肥下的优势属,但2021 年土壤近明球囊霉属的相对丰度从2018 年的28.38%降低为1.00%,为非优势属。可见,在长期培肥条件下,与球囊霉属和类球囊霉属相比,近明球囊霉属更容易受长期培肥影响而发生显著改变。
2018 年,与T0 相比,T2 和T3 处理下AM 真菌两性球囊霉属(Ambispora)的相对丰度分别下降85.55%和81.05%,处理间两性球囊霉属AM 真菌相对丰度有显著性差异(P<0.05)。但随培肥时间延长,2021 年不同处理间土壤两性球囊霉属物种相对丰度差异消失。与2018 年相比,2021 年AM 真菌群落属水平相对丰度下降,其中两性球囊霉属、原囊霉属(Archaeospora)、盾巨囊霉属(Scutellospora)、近明球囊霉属和球囊霉属分别降低85.74%、99.91%、97.74%、97.52%和50.35%。
2.3 有机无机肥配施对土壤AM 真菌群落α 多样性的影响
由表2 可知,与T0 相比,2018 年T2 和T3 处理下土壤AM 真菌群落的多样性指数(Shannon 指数)和丰富度指数(Chao1 指数)下降。其中,T2 和T3 处理下的Shannon 指数分别显著下降13.78%和15.69%(P<0.05);Chao1 指数分别下降9.92%和18.89%,T3处理达显著水平;但T2 和T3 处理间Shannon 指数和 Chao1 指数无显著差异。有机无机肥配施对Simpson 指数没有显著影响。
2021 年,AM 真菌多样性指数(Shannon 指数和Simpson 指数)和丰富度指数(Chao1 指数)在不同施肥处理间均无显著差异。随着培肥时间变化,2021年各处理AM 真菌群落Chao1 指数显著降低,与2018 年相比,T0、T1、T2 和T3 处理的Chao1 指数分别下降41.13%、42.16%、28.54%和10.45%。
2.4 长期有机无机肥配施对土壤AM 真菌群落β 多样性的影响
基于Bray-Curties 距离的NMDS 分析显示(图2),长期不同培肥阶段(2018 年、2021 年)不同施肥处理的土壤AM 真菌群落组成差异极显著(P=0.001,R2=0.35;P=0.001,R2=0.26)。T0 与T1、T0 与T2 和T3 间在不同年NMDS 的排序图上均明显分开(图2a,b);其中,T0 与T2、T3 间的距离明显大于其与T1 间的距离。这表明,T0 与T2、T3 处理间土壤AM 真菌群落组成的差异较T0 与T1 处理间的差异更大;而T2 和T3 处理的土壤AM 真菌群落较为相似,两处理间无明显差异。可见,采用化肥+牛粪有机肥或化肥+羊粪有机肥的培肥方式能够较好地维持AM 真菌群落结构的相似性。相似的土壤培肥方式更有利于形成相似的土壤AM 真菌群落组成。
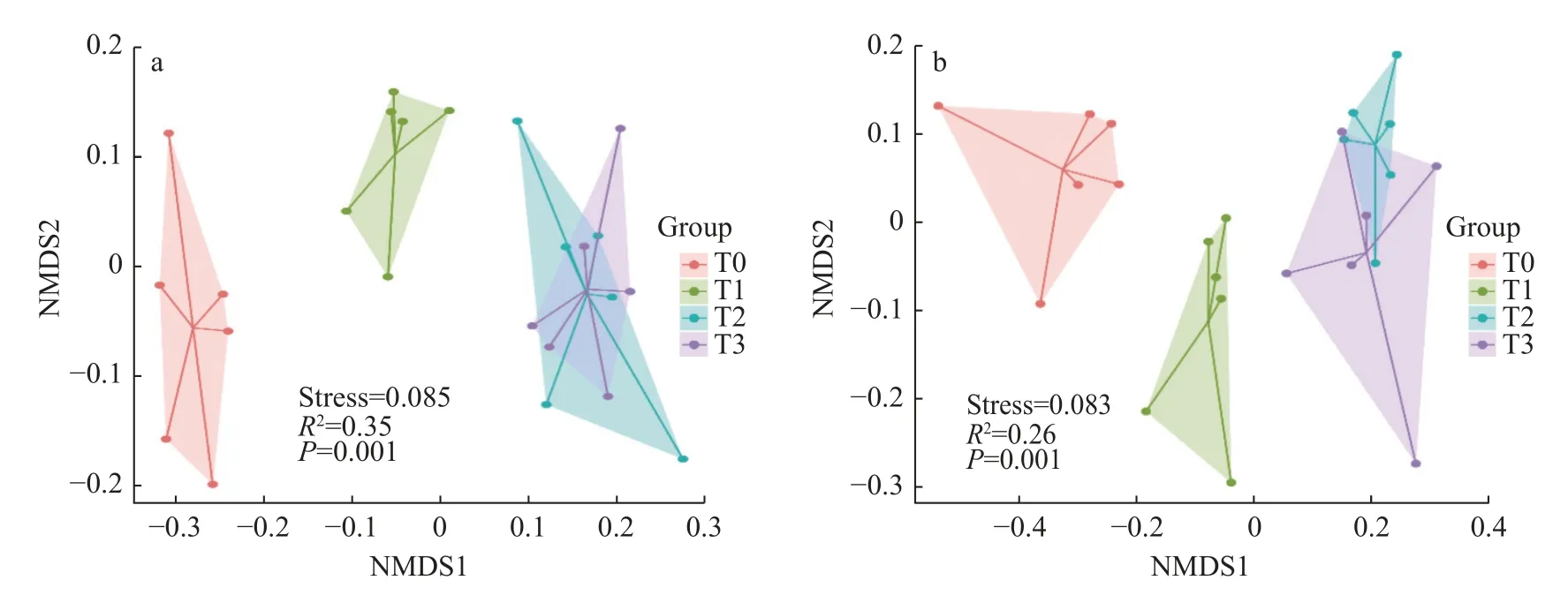
图2 2018 年(a)和2021 年(b)长期不同施肥处理土壤丛枝菌根(AM)真菌属水平群落结构的非度量多维尺度(NMDS)分析Fig.2 Non-metric multidimensional scaling (NMDS) analysis on a Bray-Curtise dissimilarity matrix of soil arbuscular mycorrhiza(AM) community composition at genus level in 2018 (a) and 2021 (b) under different long-term fertilization treatments
2.5 土壤AM 真菌群落组成、多样性与土壤理化性质的RDA 分析
冗余分析(RDA)显示: 2018 年,两个排序轴对AM 真菌群落的解释度分别为35.34%和19.83%;2021 年,第1 和第2 排序轴能够解释15.38%和10.95%的方差(图3),AM 真菌群落组成对长期有机无机肥配施处理产生了不同响应。其中,2018 年,AM 真菌丰富度指数(Chao1)和多样性指数(Simpson指数、Shannon 指数)与土壤全氮、碱解氮、有机质、速效磷呈负相关关系,与pH 呈正相关关系。2021年,AM 真菌丰富度指数(Chao1 指数)与土壤全磷、有机质、速效钾、速效磷、全氮正相关,与pH 负相关;但AM 真菌Shannon 指数和Simpson 指数与pH正相关,和其他因子负相关,与AM 真菌丰富度呈相反的趋势。属水平分类学组成中,AM 真菌均不同程度地受到土壤全氮、全磷、有机质、速效磷和pH等的影响。2018 年,球囊霉属和类球囊霉属相对丰度的改变与pH 呈正相关,而2021 年球囊霉属丰度与pH 正相关,类球囊霉属与pH 负相关;两个年度的AM 真菌球囊霉属和类球囊霉属相对丰度均与全氮、全磷、有机质等呈负相关。蒙特卡洛前向选择检验表明: 2018 年,AM 真菌群落组成和多样性仅与全氮(F=9,P=0.001)和pH (F=2.9,P=0.039)呈极显著或显著相关,与其他土壤环境因子的相关性均不显著。土壤全氮、pH 是AM 真菌群落变化的主要驱动因素,其贡献率为59.20%。2021 年,AM 真菌群落结构和多样性的主要驱动因素只有土壤全磷,贡献率为26.10% (F=2.3,P=0.035)。可见,随培肥时间的变化,受培肥方式的影响,驱动土壤AM 真菌群落结构和多样性变化的土壤因子会发生显著改变。
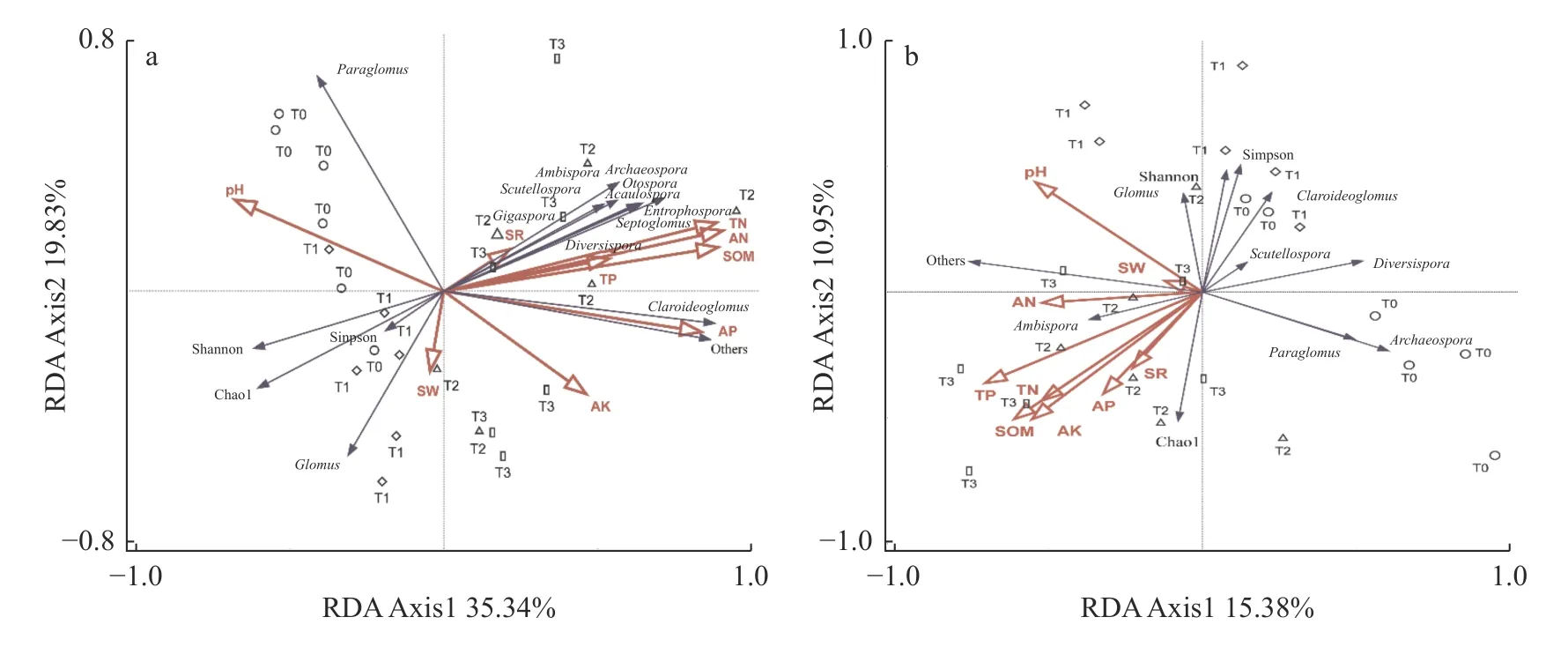
图3 2018 年(a)和2021 年(b)土壤丛枝菌根(AM)真菌群落结构、多样性与土壤环境因子的RDA 分析Fig.3 Redundancy analysis (RDA) of soil arbuscular mycorrhiza (AM) community structure,diversity and soil environmental factors in 2018 (a) and 2021 (b)
3 讨论
3.1 长期有机无机肥配施对土壤的影响
本研究中,长期培肥使土壤有机质、全氮含量增加,pH 下降,有机无机肥配施处理效果最显著,这与马桂秀等[18]的研究一致。多年的种植、培肥改善了土壤结构,作物植株的枯落物和根系分泌物为土壤微生物生长提供了有利条件,促进土壤腐殖质的形成,增加了土壤有机质含量。牛粪等有机肥的施入直接为土壤提供丰富的有机物质,当有机质被分解后会产生大量的有机酸和腐殖酸等物质,能够释放土壤中原本不易分解和转化的矿质养分,使土壤中的养分含量大幅度提高[19]。有机无机配施培肥下土壤有机质含量高于单施化肥,这与牛粪、羊粪本身含有较高有机碳,连续施用改善了土壤养分的供给特征有关。以往的研究表明,土壤有机质和全氮之间呈显著正相关,有机质含量的提高促进了全氮的积累[20],这与本研究结果类似。
本试验中,长期有机无机肥配施下土壤pH 显著下降。李娟等[21]的研究表明,长期施肥处理下土壤pH 存在一定程度的下降,且有机无机肥配施的土壤pH 比单施化肥更低。方凯等[22]的研究也表明,单施化肥和秸秆还田配施化肥均会使土壤pH 下降,并且秸秆还田配施化肥使pH 下降更加显著。以上研究均支持了本研究的试验结果。相关研究认为,有机粪肥中富含微生物代谢所需要的营养物质,能够促进微生物代谢产生有机酸[23]。此外,有机无机肥配合施用会增加土壤硝酸盐含量[24],土壤中NH4+-N 转化成NO3−-N 时,也会释放大量H+,加速土壤酸化[25]。刘灿等[26]进一步证实,在施用化肥基础上增施有机肥其酸化速率大于单施化肥;有机肥施入后土壤阳离子交换量下降幅度比单施化肥处理更大,也是导致土壤pH 降低的主要原因。另外,本研究还发现随着种植年限的增加,不施肥处理土壤pH 也在下降,这是因为本研究试验地是秋季新推耕地,属于新垦原生土。旱作区原生土碱性较强,土壤有效性差,尤其腐殖质高度匮乏[27]。新垦生土有机质含量很低,在熟化过程中,由于多年连续施用有机肥以及轮作作物残茬和残留根系的影响,有机质含量不断增加,被分解后产生大量有机酸和腐殖酸等物质,从而降低土壤pH。
与2018 年相比,2021 年后T0、T1 和T3 处理土壤全磷、速效钾显著下降。分析认为,这是由于作物收获带走的土壤磷钾量大于土壤中输入的磷钾量。樊红柱等[28]发现,连续施肥33 年后土壤磷含量降低,作物吸收携出磷量为输入土壤磷总量的6 倍,土壤磷表现出亏缺状况。相关研究表明,在轮作种植模式中作物需钾量较大,小麦-玉米-棉花(Gossypium hirsutumL.)[29]、早稻(Oryza sativaL.)晚稻和单季稻轮作土壤钾素均表现亏缺[30]。本试验中玉米和马铃薯都属于喜磷、喜钾作物,对磷钾需求量比较大,作物根系具有一定溶磷钾能力,对土壤中难溶性磷钾吸收利用效果较好[31],在持续种植下导致土壤磷、钾含量下降。另一方面,有机肥在腐化过程中产生有机酸根离子活化了土壤磷素,降低了磷素的吸附,使得磷素更易于移动,全磷、有效磷含量随施肥年限的延长都表现出下降趋势,导致磷素大量损失[32],进而降低了表层土壤全磷和有效磷的含量。由此可见,为保证土壤养分均衡,在旱作区长期有机无机肥配施过程中,应适当提高磷肥、钾肥的投入比例,避免土壤磷、钾的匮缺,而导致土壤磷、钾含量持续下降。
3.2 长期有机无机肥配施对土壤AM 真菌群落组成及多样性的影响
Huo 等[33]的研究表明,土壤AM 真菌群落结构组成显著受施肥制度影响。球囊霉属和原囊霉属AM 真菌的相对丰度在磷肥和羊粪配施处理下较高;而化肥处理下,AM 真菌近明球囊霉属、两性球囊霉属、盾巨囊霉属的相对丰度较高,无肥处理下的类球囊霉属和原囊霉属相对丰度更高,每种施肥制度下都有独特的AM 真菌菌属,AM 真菌属是受长期施肥制度驯化影响的。但试验中,土壤AM 真菌球囊霉属和类球囊霉属相对丰度并未受到施肥管理差异的影响,仍为优势属。相关研究证实,土壤球囊霉属和类球囊霉属AM 真菌具有特定的繁殖方式,不易受外界施肥的干扰,能够在各种极端环境下生存。其中,球囊霉属AM 真菌可通过菌丝体侵染根段来定殖新根,其产孢率高,增长速度快,有较强的生态适应性,更容易生存和繁殖[34];而类球囊霉属与球囊霉属属于同一科,孢子特征以及丛枝和内生菌丝的形态与球囊霉属相似,从而具有相似的特性[35]。此外,球囊霉属和类球囊霉属AM 真菌在土壤中发挥着促进作物对氮磷吸收的功能,球囊霉属还可以通过分泌球囊霉素提高土壤有机质含量,改善土壤排水通气状况[36]。因此,作物会倾向于选择球囊霉属和类球囊霉属作为优势类群从而有利于自身生长。
本研究中,有机无机肥配施(T2、T3)处理下,2018—2021 年土壤明球囊霉属AM 真菌的相对丰度显著下降,从优势属变为非优势属。有研究表明,明球囊霉属相对丰度与有机质、全氮呈显著负相关[37],与pH、全磷呈正相关[38]。分析认为,本试验长期培肥下土壤有机质、全氮含量增加,pH 和全磷下降是导致明球囊霉属丰度下降的重要因素。此外,Sheng等[39]认为,明球囊霉属能形成大量的丛枝和泡囊,而丛枝和泡囊与土壤有效钾含量呈正相关。综合分析认为,长期施肥制度下,随培肥时间变化,不同培肥措施下土壤钾含量均呈显著下降趋势,受土壤钾等肥力因子差异的影响AM 真菌出现不同的驯化趋势,这可能是导致土壤近明球囊霉属AM 真菌相对丰度显著下降的原因。
随培肥时间延长,试验中土壤AM 真菌α 多样性显著下降,AM 真菌也从12 属下降到7 属,有5 属相对丰度下降到极低水平或未检测出,这可能与土壤环境变化有关。分析认为,一方面,长期有机无机肥配施显著提高了土壤全氮、有机质等养分含量,植物不需与AM 真菌形成特别多的共生关系也能获取足够的养分;相应的AM 真菌对作物根系的侵染受到抑制,造成AM 真菌各属的丰度下降[40]。另一方面,土壤磷[41]钾[42]含量直接影响到AM 真菌的生长发育,试验中土壤全磷、速效钾含量下降,AM 真菌难以获取足量的磷钾,生长发育受到抑制进而导致丰度下降。此外,不同AM 真菌在生理和生态特性方面存在差异,pH 降低可能也会导致个别喜好高pH 的AM 真菌属丰度下降[43],一些耐受性不强的AM 真菌类群变得稀少甚至消失。
本研究中长期有机无机肥配施处理(T2、T3)下,2018 年AM 真菌群落的α 多样性指数(Shannon指数、Chao1 指数)较不施肥对照(T0)显著下降,这与Wang 等[44]的报道结果一致。一方面,有机物料的施入使得某些特定真菌类群产生“偏好”效应,消耗大量营养物质,从而抑制了其他类群的生长[45];另一方面,有机粪肥持久性氮的缓慢释放,导致土壤氮含量升高,生态系统的孢子丰度和根的定殖明显减少,AM真菌物种丰富度和多样性下降[46]。2021 年,土壤AM 真菌Shannon 指数和Chao1 指数在不同处理下无明显变化。随着培肥年限增加,AM 真菌α 多样性随着时间推移差异性消失。Herrmann 等[47]的结果表明,AM 真菌群落丰富度随时间变化并未表现出显著性差异,这可能与肥料施入后土壤肥力变化以及部分土壤养分含量下降有关。本研究下AM 真菌群落随时间变化差异性消失,可能是因为大量的AM 真菌物种已经适应了旱作区长期施用有机无机肥料类型和肥料负荷的结果。
NMDS 结果表明,随着培肥时间变化,土壤AM真菌群落的β 多样性差异依然存在,但不同有机无机肥配施方式(T2、T3)间土壤AM 真菌群落结构没有明显差异,这与Verbruggen 等[48]的研究吻合,即与传统管理下的土壤相比,施有机肥的土壤中AM 真菌群落更相似,这可能是由于有机无机肥配施处理(T2、T3)的土壤理化性质(如pH、有机质、全氮、全磷)较为接近[13],为AM 真菌的发育和繁殖提供了较为相似的生存环境,更易于驯化相似的AM 真菌菌属群落,其对AM 真菌类群的选择效应也相似[4]。
3.3 AM 真菌群落组成和多样性与土壤环境因子间的相互联系
Wang 等[49]证实,单施氮肥显著降低了AM 真菌群落的丰富度和多样性;高无机氮磷供应率对AM真菌的定殖产生了负面影响[50],而适宜氮和磷肥配合施用则显著增加了AM 真菌的丰富度和多样性[49]。在旱作雨养农业生态系统中,长期氮和/或磷添加导致的土壤性质变化被认为是影响AM 真菌群落结构的主要因素。本试验中,驱动土壤AM 真菌群落组成和多样性变化的环境因子是土壤全氮、pH 和全磷。相关研究表明,土壤全氮、pH 和土壤全磷[51-52]是影响AM 真菌丰富度和多样性的主要变量。全氮与全磷之所以会影响AM 真菌群落,是因为土壤中的氮磷含量会直接影响植物对AM 真菌的碳供应水平,进而影响AM 真菌的生长发育。相关研究指出,当土壤中氮或磷足以促进植物生长时,植物向AM真菌输送较少的碳以换取养分;相反,如果土壤中氮或磷有限,植物向AM 真菌输送的碳增多,导致AM真菌丰度增加。土壤pH 则影响菌根的形成、AM真菌的产孢及种属分布[53],进而影响AM 真菌的群落组成和多样性。然而,Ma 等[4]认为,土壤有机质也是影响AM 真菌群落组成的最重要因素,这与本研究结果不一致,可能与研究区域的气候条件、耕作措施及土壤类型等因素有关。
有研究表明,AM 真菌多样性随土壤磷含量增加而上升,但是当土壤磷含量升高到一定程度时,AM真菌多样性呈下降趋势,其原因是土壤高磷含量会改变宿主植物根系分泌物,导致AM 真菌生存环境发生改变[46]。Yoneyama 等[54]研究发现,低氮能使高粱[Sorghum bicolor(L.) Moench]根系分泌一种促进AM 真菌菌丝生长的物质,但土壤中氮大量积累却降低了AM 真菌丰富度[55],说明氮过高对AM 真菌多样性水平不利。以上研究说明AM 真菌群落对土壤中氮磷含量比较敏感,氮磷含量高或者过低都会抑制菌根的生长发育。本研究中,随培肥时间的变化,驱动AM 真菌群落变化的主要环境因子从全氮、pH 变成全磷,可能是由于前期土壤全磷含量适中,较低水平的全氮对AM 真菌群落影响更大,而后期全磷含量低到较低水平,全氮升高到适宜含量,全磷成为驱动AM 群落变化的主要环境因子。
4 结论
连续培肥11 年显著增加了土壤全氮和有机质含量,显著降低了土壤pH,但长期有机无机肥配施并没有同步提升土壤全磷、速效钾等理化性状指标。长期连续有机无机肥配施影响了土壤AM 真菌群落组成及多样性,AM 真菌中球囊霉和类球囊霉等优势属并未受到有机无机肥配施影响,但与2018 年相比,2021 年后土壤近明球囊霉属AM 真菌由优势属变为非优势属,基于属水平的AM 真菌物种组成及相对丰度显著减少和降低。本研究认为,培肥通过改变土壤全氮、pH 和全磷等土壤理化性质,一方面直接影响AM 真菌对氮磷钾的吸收和种间竞争关系,另一方面会影响植物对AM 真菌的碳供应水平,从而影响AM 真菌的群落组成和多样性。AM 真菌群落受土壤环境因子影响会选择更敏感的驱动因素。2018 年驱动土壤AM 真菌群落结构和多样性变化的土壤因子为土壤全氮、pH,而持续培肥到2021 年驱动因子改变为土壤全磷。
在宁夏中部干旱区连续11 年的施肥研究,为该地区AM 真菌群落变化与土壤环境因子间相互关系研究积累了数据,阐明了培肥对AM 真菌群落组成和多样性的影响机制,为揭示宁夏旱作区不同施肥制度下AM 群体的生态位分化提供了有力的证据,对合理利用有机无机肥管理,平衡施肥,提供了重要的理论支撑。因此,在宁夏中部干旱区合理实施有机无机肥配施制度,最大限度地促进AM 真菌的生长和活动,提供植物所需的营养元素,改善土壤环境,这有助于提高农作物的产量和质量,优化农业生产系统。
猜你喜欢
中国民间疗法(2021年1期)2021-04-20
石材(2020年9期)2021-01-07
中国盐业(2018年20期)2019-01-14
实用临床医学(2016年8期)2016-06-07
现代农业(2016年3期)2016-04-14
山东青年(2016年2期)2016-02-28
山东医药(2015年14期)2016-01-12
土壤与作物(2015年3期)2015-12-08
微创泌尿外科杂志(2014年4期)2014-02-28
华东理工大学学报(自然科学版)(2014年5期)2014-02-27
