
不同成熟度鲜食嫩南瓜种子活力及生理生化特性研究
2024-04-02 11:41赵夏云瞿飞杨静马关鹏周麟笔蒋芳芳文林宏
中国瓜菜 2024年3期
赵夏云 瞿飞 杨静 马关鹏 周麟笔 蒋芳芳 文林宏


摘 要:为了探究鲜食嫩南瓜种子成熟过程中种子活力及生理生化特性的变化趋势,以自育品种为试验材料,对不同成熟度采收的种子活力指标和生理生化指标进行测定,分析他们之间的相关性。结果表明,随着种子的成熟,种子逐渐发育饱满,种皮木质化明显;授粉后45 d时种子已具有萌发能力,授粉后70 d种子活力最高;贮藏物质在授粉后35~50 d基本完成积累,脂肪、蛋白质、淀粉大量形成,而可溶性总糖和可溶性还原糖含量同期持续减少。吸胀8 h的南瓜种子中SOD、CAT、POD活性受MDA含量变化影响,随着成熟度的增加,呈先升高后降低再升高的变化趋势;各内源激素含量变化波动不同,其中SA与BR含量整体上呈显著增长趋势且高于其他激素,GA3与ABA含量变化趋势相反,ETH含量授粉后35 d最高。相关性分析表明,种子活力指标与SOD、POD活性以及淀粉、粗脂肪、CTK、GA3、IAA、BR、SA含量呈正相关,而与可溶性总糖、可溶性还原糖、蛋白质、MDA、ABA、ETH含量呈负相关。综上,不同成熟度对鲜食嫩南瓜种子质量和种子活力有显著影响。鲜食嫩南瓜种子发育过程中的生理生化过程复杂,伴随着贮藏物质的大量积累,抗氧化酶活性与MDA、内源激素含量不断变化。
关键词:鲜食嫩南瓜;种子活力;生理生化特性;成熟度
中圖分类号:S642.1 文献标志码:A 文章编号:1673-2871(2024)03-103-09
Study on seed vigor and physio-biochemical characteristics of fresh-Cucurbita moschata seeds of different maturity
ZHAO Xiayun1, QU Fei1, YANG Jing1, MA Guanpeng1, ZHOU Linbi1, JIANG Fangfang2, WEN Linhong1
(1. Institute of Horticulture, Guizhou Academy of Agricultural Sciences,Guiyang 550025, Guizhou, China; 2. Majiang Economic Crop and Technology Promotion Station, Majiang 557600, Guizhou, China)
Abstract: To explore the changes of seed vigor and physio-biochemical characteristics of fresh-Cucurbita moschata seeds during the maturation period, a self-breeding variety was used as materials to analyze the relationship of the vigor and physio-biochemical indexes of seeds with different maturity. The results showed that the seeds gradually developed plump morphology with the maturity, and the seed coat was obviously lignified. After 45 days post-anthesis (dpa), the seeds began to appear germinating ability and reached a peak at 70 dpa. During 35-50 dpa, the accumulation of stored substances was basically completed, and large quantities of fat, protein and starch were formed, while the content of total soluble sugar and soluble reducing sugar continued to decrease. With increased seed maturity, the activities of SOD, CAT and POD in seeds after 8 h imbibition were affected by MDA content, with a trend of first increasing and then decreased. The content of endogenous hormones varied, with of SA and BR showing a significant increasing trend overall and higher than other hormones, while GA3 and ABA showed an opposite trend, and ETH was the highest at 35 dpa. Correlation analysis indicated that seed vigor indexes were positively correlated with SOD activity, POD activity, the contents of starch, crude fat, CTK, GA3, IAA, BR and SA, but negatively correlated with the contents of total soluble sugar, soluble reducing sugar, protein, MDA, ABA and ETH. In conclusion, the development of fresh-Cucurbita moschata seeds was accompanied by the accumulation of storage substances, the increase and decrease of antioxidant enzyme activities and MDA content, the change of endogenous hormone content and the complex physio-biochemical process.
Key words: Fresh-Cucurbita moschata; Seed vigor; Physio-biochemical characteristics; Maturity
鲜食嫩南瓜是指中国南瓜类型中以嫩瓜作为主要食用器官的品种,属于葫芦科南瓜属一年生草本蔬菜,在贵州、湖南、广东等地均有大面积种植。鲜食嫩南瓜果实鲜嫩甘甜,果型小,营养丰富,深受消费者的喜爱,有很好的市场前景。在南瓜生产中,种子至关重要,种子质量直接关系到南瓜植株的生长、果实品质和产量。种子成熟度则是种子质量的重要指标,也是南瓜集约化生产和优质高产的必要质量指标。
在种子发育过程中,种子活力随着种子成熟度变化而变化。随着种子的成熟,贮藏物质不断累积,种子活力逐渐提高,为种子萌发和幼苗生长打下基础。种子中含有的主要贮藏物质包括脂肪、蛋白质、淀粉等,在种子萌发和幼苗生长过程中被分解和利用[1-2]。种子活力检测的方法有很多,标准发芽试验与生理生化测定结合,能够综合测定鲜食嫩南瓜种子活力[3]。
种子萌发还涉及相关酶和激素活化及其生理生化过程。丙二醛(malondialdehyde,MDA)是膜脂过氧化的最终分解产物,在种子萌发过程中过量积累会损伤细胞膜,影响种子活力[4-5]。超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)和过氧化氢酶(catalase,CAT)是植物中的抗氧化酶,通过调节酶活性,减少活性氧积累造成的损伤,维持种子正常萌发[6-8]。脱落酸(abscisic acid,ABA)具有诱导和维持种子休眠,抑制种子萌发的作用[9]。赤霉素(gibberellin,GA3)则打破休眠、促进种子萌发和拮抗ABA[10-11]。乙烯(ethylene, ETH)在种子萌发过程中能够促进胚轴伸长和根毛分化[12-13]。生长素(indole-3-acetic acid,IAA)与ABA协同作用调控种子休眠,抑制种子萌发[14]。油菜素内酯(brassinolide,BR)通过不同信号途径促进种子萌发[15]。水杨酸(salicylic acid,SA)能通过下调GA3诱导的α淀粉酶活性从而抑制种子萌发,也能在高盐胁迫下通过其他途径促进萌发[16]。细胞分裂素(cytokinin,CTK)能够逆转ABA对种子萌发的抑制,与GA3、IAA和ETH等激素协同影响种子萌发[17]。
目前,关于鲜食嫩南瓜种子发育过程中种子活力与生理生化变化相关性的研究较为缺乏。笔者以自育杂交组合为材料,在测定不同成熟期种子性状、种子贮藏物质的基础上,进一步研究不同成熟度种子活力与萌动种子中的抗氧化酶活性以及MDA、内源激素含量等生理生化特性之间的相关性,以期为解析鲜食嫩南瓜种子活力形成机制和确定杂交制种的合适采收时间提供依据。
1 材料与方法
1.1 材料
试验于2022年5-12月在贵州省农业科学院园艺研究所进行。供试材料为自育鲜食嫩南瓜(Cucurbita moschata)杂交组合GG307(G340×G477),母本、父本分别是韩国玉玲珑、贵阳小青瓜经连续8代自交选育而成的自交系。选取饱满种子,温汤浸种后,播种到32孔穴盘中,2叶1心时定植到田间。试验地面积667 m2,采用高畦覆膜栽培,行距0.7 m,株距0.6 m,呈三角形定植。
選择长势基本一致的植株,分别将雌花、雄花套袋,在第二天早上完成全部人工授粉,每株1个种瓜。完全随机设计试验,授粉后35 d开始,每隔5 d采收种瓜一次,直至70 d,共取样8个时期。每个时期3次重复,5个种瓜的种子混合为1次重复,不经后熟,直接取种自然晾干后用于种子活力及生理生化指标测定。
1.2 方法
1.2.1 种瓜性状及萌发特性测定 种瓜取种前测量单瓜质量,种子晾晒干燥后统计千粒重。取不同成熟期的种子各100粒,3次重复,浸种后进行室内种子发芽试验。将种子置于恒温培养箱29 ℃条件下,每天定点观察统计正常发芽粒数,3 d统计发芽势,7 d统计发芽率,7 d后用1/1000天平称量各处理中正常幼苗单株鲜质量,分别计算发芽指数和活力指数。
发芽势/%=3 d正常发芽种子粒数/供试种子粒数×100; (1)
发芽率/%=7 d正常发芽种子粒数/供试种子粒数×100; (2)
发芽指数(GI)=Σ(Gt/Dt); (3)
活力指数(VI)=GI×S。 (4)
公式中,Dt:发芽日数,Gt:与Dt相对应的每天发芽种子数,S:发芽试验结束时单株幼苗鲜质量。
1.2.2 贮藏物质含量测定 取不同成熟期自然干燥的混收种子,磨碎后进行贮藏物质含量测定,3次重复。采用3,5-二硝基水杨酸法测定种子可溶性还原糖含量[18];采用蒽酮比色法检测种子可溶性总糖及淀粉含量[18];采用BCA(bicinchoninic acid assay,二辛可宁酸)法测定蛋白含量[19];采用脂肪测定仪法测定粗脂肪含量[18]。
1.2.3 酶活性测定 种子消毒后用蒸馏水浸泡8 h,在29 ℃下萌发3 h,去除种皮后测定酶活性,3次重复。采用硫代巴比妥酸法测定MDA含量[20];采用水溶性四唑盐法测定SOD活性[20];采用愈创木酚法测定POD活性[20];采用紫外分光度法测定CAT活性[20]。
1.2.4 内源激素含量测定 按照上海酶联公司试剂盒(ml077222、ml077231、ml750790、ml780845、ml803924、ml077224、ml970662)方法,分别对吸胀8 h后的不同成熟度鲜食嫩南瓜种子中的ABA、IAA、GA3、CTK、BR、SA、ETH含量进行检测,3次重复。
1.3 数据处理与分析
使用Microsoft Excel 2019和IBM SPSS Statistics26.0进行差异显著性统计分析。使用Photoshop CC 2018软件绘制处理图片。使用R语言进行相关性分析。
2 结果与分析
2.1 不同成熟度鲜食嫩南瓜种瓜及种子性状变化
随着鲜食嫩南瓜种子的成熟,种皮边缘颜色逐渐加深,棱齿突出,且种皮木质化程度增加,变为黄白色。整个过程中,种子体积呈先迅速增大后平缓缩小的变化趋势,种子也由干瘪逐渐发育饱满(图1-A)。授粉后35~50 d为种子干物质快速积累期,干种子的千粒重从69.70 g增加到104.70 g,显著提高了50.22%,平均每天增加2.33 g,子叶厚度也明显增加。达到生理成熟后,种子干物质积累速度变缓,授粉后55~70 d,千粒重差异不显著。种瓜单瓜质量的变化趋势与干种子千粒重基本一致,授粉后35~50 d迅速增加,之后缓慢增加,65 d时单瓜质量最高,为2 797.93 g,比35 d时显著提高了837.53 g,70 d时小幅减少,为2 683.05 g(图1-B)。
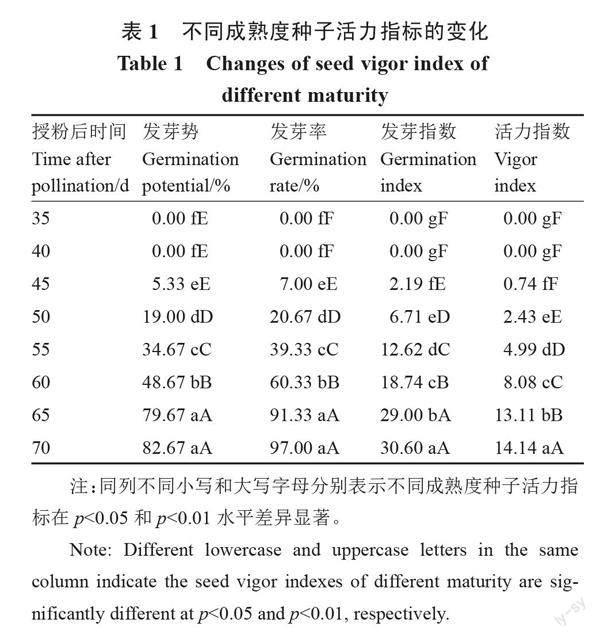
2.2 不同成熟度鲜食嫩南瓜种子活力的变化
鲜食嫩南瓜种瓜不同成熟度对其种子活力指标有显著影响。由表1可以看出,在授粉后45 d,种子具有发芽能力,但其发芽势、发芽率、发芽指数和活力指数均极低。授粉后45~60 d,种子相应活力指标随着成熟度的增加逐渐升高,但发芽率均未达到61%。与授粉后60 d相比,授粉后65、70 d的种子发芽率均显著提高,分别为91.33%、97.00%,所有指标均显著提高了50%以上。
2.3 不同成熟度鲜食嫩南瓜种子主要贮藏物质的变化
由图2可知,在不同成熟度的鲜食嫩南瓜种子中,随种子成熟度增高,可溶性总糖和可溶性还原糖含量均呈现先显著降低后趋于稳定的变化趋势,蛋白质含量呈先增加后降低的变化趋势,粗脂肪和淀粉含量则呈先显著增加后趋于稳定的变化趋势。授粉后35 d,种子中可溶性总糖和可溶性还原糖含量(w,后同)最高,分别为110.72、33.55 mg·g-1,50 d时分别为85.91、24.44 mg·g-1,分别显著降低了22.41%、27.15%,55~70 d可溶性总糖和可溶性还原糖含量波动较小,可溶性还原糖含量差异不显著。蛋白质含量在授粉后55 d达到最高,为293.74 mg·g-1,45~55 d间蛋白质含量差异不显著,60 d时蛋白质含量极显著降低,70 d降低至248.94 mg·g-1。粗脂肪含量在授粉后35~50 d迅速积累,50 d粗脂肪含量达到最高,为382.88 mg·g-1,比35 d时显著升高了149.30%,平均每天积累15.29 mg·g-1,之后小幅波动。淀粉含量与粗脂肪含量变化趋势相似,35~50 d时显著提高,50~70 d趋于稳定。
2.4 不同成熟度鲜食嫩南瓜种子抗氧化酶活性及MDA含量的变化
由图3可知,在吸胀8 h的种子中,MDA含量(b)在授粉后35~55 d呈显著下降的趋势,55 d时降至最低,为21.17 nmol·g-1,60~70 d快速增加后小幅减少。CAT、SOD和POD活性在授粉后35~55 d呈上升趋势,且均在35~45 d活性上升最快,均在55 d时达到最大值,分别为2 413.24 U·g-1,460.76 U·g-1和16 646.35 U·g-1,此后活性呈先降低后升高的波动但均保持较高水平。
2.5 不同成熟度鲜食嫩南瓜种子内源激素含量的变化
由图4可知,吸胀8 h的不同成熟度种子中的7种内源激素含量随着种子成熟度的增加呈现出不同的变化趋势。其中,ABA呈降-升-降-升-降的变化趋势,授粉后35~45 d含量变化不显著,含量最高点出现在授粉后55 d。随着种子成熟度的增加,GA3含量呈先极显著降低后极显著升高的动态变化,与ABA相反,授粉后55 d时含量最低。SA与BR含量整体上呈增长趋势,但均在55 d时突然极显著降低。ETH含量在授粉后35 d的萌动种子中最高,极显著降低后趋于平稳。在不同成熟度的萌动南瓜种子中,CTK和IAA含量变化不规律,CTK含量总体呈现出较低的水平,而SA和BR含量整体水平明显高于其他激素。
2.6 萌发特性与生理特性的相关性分析
对不同成熟度鲜食嫩南瓜种子各项指标进行相关性分析,结果显示(图5),发芽率、发芽势、发芽指数、活力指数4个种子活力指标均与SOD、POD活性及淀粉、粗脂肪、CTK、GA3、IAA、BR和SA含量呈正相關,而与可溶性总糖、可溶性还原糖、蛋白质、MDA、ABA和ETH含量呈负相关。另外,发芽势与BR和SA含量呈极显著正相关,而发芽率、发芽指数和活力指数与淀粉、SA含量呈显著正相关,与BR含量呈极显著正相关。可溶性总糖和可溶性还原糖含量与除MDA、GA3、IAA、ETH含量以外的其余指标呈负相关。推测,可溶性总糖和可溶性还原糖对鲜食嫩南瓜种子萌发有一定的抑制作用。
3 討论与结论
伴随着种子体积增大、种皮边缘颜色加深、种皮木质化程度加剧等形态变化,鲜食嫩南瓜种子先后达到生理成熟和形态成熟状态。在鲜食嫩南瓜种子发育过程中,千粒重变化明显,发育初期快速增加,生理成熟后趋于平稳。
鉴定种子活力最直观有效的方式就是萌发试验。冯志红等[21]采用发芽试验对授粉后30~55 d的西葫芦杂交种子活力进行检测,发现授粉后不足40 d的种子质量极差,授粉后45 d以上的种子质量较好。在本研究中,过早采收会显著影响到种子的千粒重、发芽率、发芽势等活力指标,随着种瓜成熟度增加,种子千粒重趋于稳定,达到完熟后,发芽率显著提高。孙守如等[22]对西葫芦收种指标研究发现,种瓜成熟度越高,种子活力越高,且随着后熟时间增加,不同瓜龄的种子活力也显著提高。后熟对早期采收的甜瓜种子的发育同样非常重要,成熟度越低,后熟作用越显著,但种瓜后熟作用并不能完全取代植株对种子的影响[23]。在本研究中,未经后熟的鲜食嫩南瓜种子,直至授粉65 d之后,发芽率才达到90%以上。推测,鲜食嫩南瓜种子在发育后期经过质的变化,才达到真正的成熟,而后熟处理可以促使早采收的种子完熟,具备发芽能力,但相关的生理生化过程还有待于进一步研究。
种子发育中还涉及贮藏物质的积累和存储,他们为种子萌发和活力激发提供有效的动力和养分,其含量对种子活力有制约作用。本试验结果表明,淀粉、粗脂肪含量在授粉后35~50 d快速升高,这也是种子千粒重同期迅速增加的原因,相关性分析也表明了他们的积累与鲜食嫩南瓜种子发育指标呈正相关。而可溶性总糖和可溶性还原糖则在种子成熟的过程中逐渐转化为其他贮藏物质,其含量呈先快速降低后趋于平稳的变化趋势。值得注意的是,蛋白质在授粉后35~50 d积累速度比较缓慢,可能是由于蛋白质的积累要先于淀粉和脂肪。笔者在研究中还发现,贮藏物质中,脂肪、蛋白质的含量较高,这与前人关于南瓜种子营养成分的研究相吻合[24-25]。
种子在萌发过程中新陈代谢旺盛,利用自身营养物质供种子萌发时会产生很多活性氧,从而诱发抗氧化酶活性的增强,以消除过氧化物[6,26-27]。在本研究中发现,在授粉后35~55 d的萌动种子中,SOD、POD和CAT酶活性均呈升高趋势,而MDA含量显著降低;授粉后60~70 d,SOD、CAT抗氧化酶活性先降低后升高,POD活性逐渐升高,而MDA含量随之降低。这说明随着种子成熟度的增加,种子萌发时新陈代谢旺盛,抗逆能力增强,有利于萌发;成熟度过高,抗逆能力有所减弱,但还是具有较强的保护及修复能力。刘欢等[28]在米槁种子发育的研究中同样发现,MDA含量与抗氧化酶活性呈此消彼长的变化趋势。不同作物种子在萌动过程中,各抗氧化酶被激发的活性程度是不同的[5,7-8,29]。本研究中,POD和CAT酶活性整体更强,鲜食嫩南瓜在种子萌动过程中更多参与了活性氧的代谢。
种子萌发过程非常复杂,需要多种激素的协同精细调控。在种子萌发过程中,各内源激素含量因种子成熟度的不同而发生变化,进而影响种子萌发。鲜食嫩南瓜种子在萌动过程中,GA3在成熟度很低和很高的种子中大量合成,GA3与ABA含量基本上呈相反的变化趋势,表明他们之间存在拮抗作用。在种子萌发过程中,BR和SA通过激活抗氧化酶系统、调节α-淀粉酶活性等来增强种子抗逆性[30-31]。在本研究中,不同成熟度种子中BR和SA的相对含量最高且与种子活力呈显著或极显著正相关,表明BR和SA在南瓜种子萌发的起始阶段发挥重要作用。有研究表明,CTK在种子萌发过程中具有剂量效应[32-33],本研究中CTK整体含量很低,但与种子活力指标呈正相关。推测在种子萌发阶段,CTK以低含量促进南瓜种子萌发。
种子发育过程中的贮藏物质累积、相关酶及激素的调节与代谢等一系列特定生理生化变化间存在相互作用。本研究中,在种子发育初期,种子活力随种子成熟度增加而升高,但整体活力很低。在这一阶段,伴随着淀粉、蛋白质、脂肪的累积,种子抗氧化酶活性不断增强,MDA、GA3和ETH含量持续降低,SA、BR和ABA含量则持续增加。在种子发育成熟阶段,贮藏物质完成积累,种子抗氧化酶活性有所降低,但整体维持在较高活性水平,SA和BR含量大幅增加,种子活力大幅提升,表明这两种激素与种子活力有更为重要的关系。种子发育过程极其复杂,各生理生化指标间如何相互作用还有待深入研究。
综合鲜食嫩南瓜种子成熟过程中种子性状和生理生化变化的特性,笔者认为不同成熟度与鲜食嫩南瓜质量和种子活力密切相关。随着种子成熟度增加,种皮逐渐木质化,种子干物质大量积累,千粒重增加,种子活力提升。种子活力与BR含量呈极显著正相关,与淀粉、SA含量呈显著正相关。
参考文献
[1] 王萍,赵清岩,王若菁,等.籽用南瓜种子成熟过程中主要营养成分的变化[J].园艺学报,2001,28(1):47-51.
[2] 崔竹梅,袁菲菲,韩耀平.南瓜种子萌发时贮藏蛋白及其酶活力的动态变化[J].安徽农业科学,2009,37(36):17937-17938.
[3] 蒋敏明.种子活力研究进展及展望[J].中国种业,2018(6):15-18.
[4] 陈思羽,刘鹏,朱末,等.大豆植株不同冠层种子活力及其萌发中抗氧化酶活性[J].植物学报,2016,51(1):24-30.
[5] 贺振,曲比伍合,蔡林,等.不同成熟度正红431种子抗逆指标及发芽率变化研究[J].四川农业大学学报,2022,40(1):36-41.
[6] DONG J A,WAN G W,LIANG Z S.Accumulation of salicylic acid--induced phenolic compounds and raised activities of secondary metabolic and antioxidative enzymes in Salvia miltiorrhiza cell culture[J].Journal of Biotechnology,2010,148(2/3):99-104.
[7] 谭萍,陈建桦,姚绍嫦,等.牛大力种子萌发过程中的生理生化变化[J].广西科学院学报,2020,36(2):177-183.
[8] 王娜,王佳茜,李国雷,等.栓皮栎种子萌发出苗特征与生理生化变化[J].林业科学,2022,58(4):1-10.
[9] 唐双,于安民,刘爱忠.脱落酸和赤霉素相互作用调控种子休眠或萌发的分子机理[J].分子植物育种,2022,20(20):6893-6900.
[10] NONOGAKI H.Seed dormancy and germination-emerging mechanisms and new hypotheses[J].Frontiers in Plant Science,2014,5,233.
[11] NONOGAKI H.Seed biology updates:Highlights and new discoveries in seed dormancy and germination research[J].Frontiers in Plant Science,2017,8,524.
[12] 杨荣超,张海军,王倩,等.植物激素对种子休眠和萌发调控机理的研究进展[J].草地学报,2012,20(1):1-9.
[13] 宋松泉,刘军,徐恒恒,等.乙烯的生物合成与信号及其对种子萌发和休眠的调控[J].作物学报,2019,45(7):969-981.
[14] LIU X D,ZHANG H,ZHAO Y,et al.Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis[J].Proceedings of the National Academy of Sciences of the United States of America,2013,110(38):15485-15490.
[15] 朱早兵,赖彩余,邹凯茜,等.油菜素内酯及其在种子萌发过程中的生理效应研究进展[J].热带农业科学,2015,35(5):13-18.
[16] 杨立文,刘双荣,林荣呈.光信号与激素调控种子休眠和萌发研究进展[J].植物学报,2019,54(5):569-581.
[17] 宋松泉,刘军,杨华,等.细胞分裂素调控种子发育、休眠与萌发的研究进展[J].植物学报,2021,56(2):218-231.
[18] 龙金飞,郑清岭,杨忠仁,等.不同年限对沙芥属植物种子寿命及贮藏物质的影响[J].种子,2017,36(10):15-20.
[19] WALKER J M.The bicinchoninic acid (BCA) assay for protein quantitation[J].Methodes in Molecular Biology,1994,32:5-8.
[20] 陈刚,李胜.植物生理学实验教程[M].北京:高等教育出版社,2016.
[21] 冯志红,李晓丽,宋晓飞,等.不同成熟度对西葫芦种子质量和子叶苗质量的影响[J].种子,2010,29(1):88-89.
[22] 孙守如,陈艳丽,赵香梅,等.不同瓜龄、后熟期对西葫芦采种指标的影响[J].华北农学报,2004,19(4):29-32.
[23] 李计红,雍山玉,吴玉梅,等.甜瓜种子成熟度对种子活力和幼苗生长的影响[J].甘肃农业大学学报,2006,41(1):39-42.
[24] MERU G,FU Y Q,LEYVA D,et al.Phenotypic relationships among oil,protein,fatty acid composition and seed size traits in Cucurbita pepo[J].Scientia Horticulturae,2018,233:47-53.
[25] MERU G,LEYVA D,MICHAEL V,et al.Genetic variation among Cucurbita pepo accessions varying in seed nutrition and seed size[J].American Journal of Plant Sciences,2019,10(9):1536-1547.
[26] KIBINZA S,BAZIN J,BAILLY C,et al.Catalase is a key enzyme in seed recovery from ageing during priming[J].Plant Science,2011,181(3):309-315.
[27] SANTOS F C,CAIXETA F,CLEMENTE A C S,et al.Gene expression of antioxidant enzymes and coffee seed quality during pre-and post-physiological maturity[J].Genetics and Molecular Research,2014,13(4):10983-10993.
[28] 刘欢,黄小龙,刘济明,等.米槁种子发育过程的形态和生理特性[J].东北林业大学学报,2022,50(1):21-26.
[29] 夏軍,时晓娟,郝先哲,等.低温对不同基因型棉种萌发过程中酶活性及激素含量的影响[J].植物生理学报,2019,55(9):1291-1305.
[30] 朱早兵,赖彩余,邹凯茜,等.油菜素内酯及其在种子萌发过程中的生理效应研究进展[J].热带农业科学,2015,35(5):13-18.
[31] 夏方山,毛培胜,闫慧芳,等.水杨酸对植物种子及幼苗抗逆性的影响[J].草业科学,2014,31(7):1367-1373.
[32] 邓志军,宋松泉.ABA对黑黄檀种子萌发的抑制作用以及其他植物激素对ABA的拮抗作用[J].云南植物研究,2008,30(4):440-446.
[33] ARA?JO S,PAGANO A,DONDI D,et al.Metabolic signatures of germination triggered by kinetin in Medicago truncatula[J].Scientific Repoter,2019,9:10466.
收稿日期:2023-04-21;修回日期:2023-06-29
基金项目:贵州省科技支撑项目(黔科合支撑[2021]一般212);贵州省高层次创新型人才项目(黔科合平台人才[2016]5671);贵州省科研机构创新能力建设专项资金项目(黔科合服企[2022]005);贵州省蔬菜现代农业产业技术体系项目(GZCYTX2021-01)
作者简介:赵夏云,女,副研究员,主要从事蔬菜育种及栽培技术研究。E-mail:yys_xiayun_zhao@163.com
通信作者:文林宏,男,研究员,主要从事蔬菜育种及栽培技术研究。E-mail:hong1521@163.com
猜你喜欢
新疆农业科学(2023年11期)2023-12-18
今日农业(2022年6期)2022-07-05
航天工业管理(2020年9期)2020-12-28
航天工业管理(2020年1期)2020-04-20
科学导报(2020年8期)2020-03-12
中国食品学报(2020年2期)2020-01-17
装甲兵工程学院学报(2018年4期)2018-10-19
种子(2018年9期)2018-10-15
学苑创造·B版(2018年12期)2018-03-04
装甲兵工程学院学报(2017年4期)2017-09-16
