
湖南八大公山桃叶珊瑚果实与种子的形态发育动态
2024-01-27 10:38卢小蓉刘江涛陈锦华谷志容田向荣
亚热带植物科学 2023年5期
卢小蓉,刘江涛,余 萍,李 龙,陈锦华,谷志容,何 钢,田向荣*
(1. 吉首大学生物资源与环境科学学院,湖南 吉首 416000;2. (武陵山区)植物资源保护与利用湖南省高校重点实验室,湖南 吉首416000;3. 湖南省八大公山国家级自然保护区管理处,湖南 张家界 427000;4. 湘西土家族苗族自治州林业科学研究所,湖南 吉首 416000)
在分类上,桃叶珊瑚Aucubachinensis原来归属于山茱萸科Cornaceae,后来经向秋云等学者修订后,被列入东亚特有的桃叶珊瑚科Aucubaceae[1—4]。桃叶珊瑚科植物均为分布于亚热带常绿阔叶林下的典型灌木,可作为亚热带常绿阔叶林灌木种群研究的重要材料。同时,由于其叶色常绿、叶形优美、花果颜色鲜艳,因而常用作园林植物[1];而且其所富含的次生代谢产物桃叶珊瑚苷是一些药物的中间体和抗氧化剂。因此,桃叶珊瑚是一种多用途植物。
目前,对桃叶珊瑚的研究主要集中在分类系统位置[5—6]、次生代谢产物[7—10]及其药理作用[11—13]等方面,对其园林应用和天然产物开发相关的种苗繁育研究还非常有限[14—16]。为了满足园林应用和天然产物开发的需要,桃叶珊瑚种苗繁育的瓶颈问题亟待解决。
果实与种子的发育特征是与种苗繁育有关的核心问题之一,桃叶珊瑚果实与种子发育特征的相关研究对其园林应用有重要意义。然而,桃叶珊瑚果实与种子发育的特征及影响其发育的生态限制因子尚不清楚。《中国植物志》上关于桃叶珊瑚的描述表明其花期为1~2 月,果熟期为翌年2 月[17],说明桃叶珊瑚果实与种子发育缓慢,可能有花果同期、果期重叠现象。
本研究以湖南桑植八大公山国家级自然保护区内常绿阔叶林下的桃叶珊瑚为研究材料,通过实地调查、解剖观察和相关性分析等方法对不同发育时期桃叶珊瑚果实外部形态特征、胚与胚乳的形态特征变化进行研究,主要探讨:(1) 桃叶珊瑚果实与种子发育的形态变化规律;(2) 果实/种子形态变化与温度和降雨量等生态因子的相关性;(3) 发育缓慢导致花果同期、果期重叠的原因等问题。通过对果实与种子的发育及其对生态因子响应规律的阐释,为桃叶珊瑚的保护和利用提供理论依据。
1 材料与方法
1.1 材料
2021 年4 月~2022 年4 月(13 个月),每月定期在湖南省桑植县八大公山国家级自然保护区天平山林区内(29°78′ N、110°09′ E,海拔1316.1~1462.4 m)采集桃叶珊瑚果实,选择野外坐果率最高的10 株为定点观测和采样植株(图1A、B),花冠凋落后开始采样测定,每月测定1 次,直至果实完全自然掉落,带回实验室观测。

图1 八大公山国家级自然保护区天平山林区内的桃叶珊瑚Fig. 1 Aucuba chinensis plants in Tianpingshan forest area of Badagongshan National Nature Reserve
1.2 方法
1.2.1 果实形态测量
每次从植株不同方位随机摘取发育正常、无病虫害的果实30 个,用体视显微镜(Leica M205C)对果实长度、宽度、果皮厚度进行测量与拍照。发育时间以花冠凋落后的月份数(Months after withering,MAW)表示,果形指数以果实平均长度和果实平均宽度的比表示。
1.2.2 果实颜色测定
参考梁森苗等[18]测定果实颜色的方法,用国际标准色卡(CMYK RGB)与果皮颜色对照,鉴定每个月的果实颜色,并记录其对应的红绿蓝(RGB)数值。
1.2.3 种子与胚形态测量
参考郑国琦等[19]测定种胚形态的方法,用解剖刀将果实沿着背缝线对半切开,然后置于体视显微镜下测量果实、子房壁与果皮厚度,胚乳、胚的大小,胚率为胚长度与种子长度的比值。
1.3 数据获取与统计分析
果实与种子形态数据均为平均值±标准差(mean±SD)。使用SPSS22.0 中One Way ANOVA 方法进行差异显著性分析(P<0.05)。
气温与降水等气象数据来自于采集地八大公山气象站58 年(1961~2018 年)年间的连续记录数据。将果形指数、胚比分别与月平均气温和降水进行Pearson 相关性分析,以确定果实/种子发育受生态因子影响的关联度。使用Origin 2018 绘图。
2 结果与分析
2.1 桃叶珊瑚的花期与果期
八大公山国家级自然保护区内野生桃叶珊瑚的花期为3~4 月,其中雄花花期为3 月(图1 C),雌花花期为4 月(图1 D);果熟期至翌年4 月(图1 E、F),从4 月坐果到次年4 月完成自然掉落共经历13个月,由于果期较长,花和果实发育期在3~4 月重叠,因此出现了花果同期(图1 E)和果期重叠(图1 F)现象。据实地调查42株桃叶珊瑚植株结果量发现,其果实产量较高,平均结果量为45±40 个,其中最少结果量为7 个,最多的可达173 个。
2.2 发育期果实外部形态与颜色的变化
通过对果实长度、宽度与果皮厚度进行测量,发现果实的生长发育呈现“慢-快-慢-快”的趋势,依据生长曲线可将果实形态发育过程大致分为4 个阶段(图2、图3)。

图2 桃叶珊瑚果实长度、宽度(A)与果形指数、果皮厚度(B)生长发育动态变化Fig. 2 Dynamic changes of fruit length, width (A) and fruit shape index, ovary wall and pericarp thickness (B) of Aucuba chinensis

图3 桃叶珊瑚果实不同发育时期颜色与基色色度值变化Fig. 3 Changes of color and primary chromaticity value of Aucuba chinensis fruits at different developmental stages
启动期(1~4 MAW):果实长度、宽度以及果皮厚度的平均月增长量分别为0.80 mm、0.22 mm、0.04 mm;净增量占掉落时成熟果实长度的14.83%、宽度的6.86%以及果皮厚度的8.18%;此阶段果实长度的增长显著大于宽度,果形指数保持在2.49~2.63之间,果实整体呈长圆柱形。结合图3 可知,果实颜色为青绿色,蓝色色值缓慢增加。该阶段果实发育处于蓄势待发的启动状态。
生长期 Ⅰ(4 ~8 MAW):果实长度、宽度以及果皮厚度的平均月增长量分别为1.1 mm、1.24 mm、0.11 mm;净增量占掉落时果实长度的27.30%、宽度的51.66%、果皮厚度的27.04%;果实宽度的生长速率大于长度,果实迅速膨大,果形指数迅速减小至1.57,果实由长圆柱形变至近卵圆形。果实仍然为青绿色,蓝色色值一直处于增加状态,红色和绿色色值波动较大(图3)。整体而言,果实发育进入第一个迅速增长高峰期。
停滞期(8~10 MAW):该阶段果实生长极为缓慢,果实长度、宽度、果皮厚度的平均月增长量分别为0.34 mm、0.2 mm、0.04 mm;净增量占掉落时果实长度的4.22%、宽度的4.16%、果皮厚度的4.40%;果形指数一直维持在1.56~1.57 之间,果实保持近卵圆形。该阶段果实颜色由青绿色转为黄色。
生长期 Ⅱ(10 ~13 MAW):果实长度、宽度、果皮厚度的平均月增长量分别为0.99 mm、0.42 mm、0.18 mm;净增量占掉落时果实长度的18.36%、宽度的13.20%、果皮厚度的33.3%;果形指数由1.57增长至1.70,果实长度增长快于宽度,果实向长卵圆形变化。该时期果实颜色迅速转变为红色,主要是在红色色值稳定的情况下,绿色和蓝色色值迅速下降所致。
2.3 胚乳形态变化
胚乳生长发育过程与果实形态变化规律基本相似(图4~图5),也可分为4 个阶段。
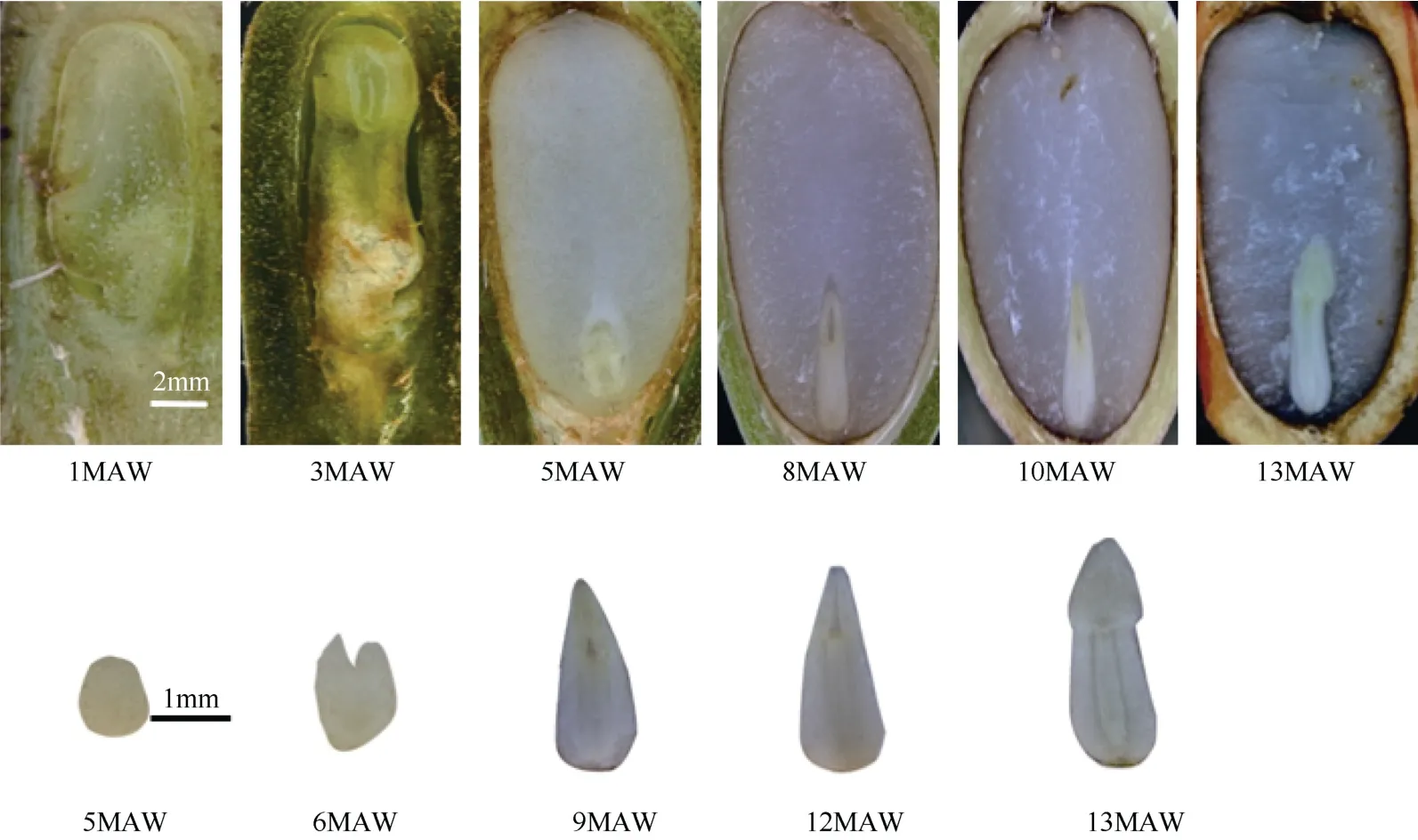
图4 不同发育时期的桃叶珊瑚果实与胚形态变化Fig. 4 Morphological changes of fruits and embryos of Aucuba chinensis at different development periods

图5 桃叶珊瑚果实在发育过程中的胚乳长度、宽度(A)与胚乳长度/宽度比值变化(B)Fig. 5 Changes of endosperm length and width (A) and endosperm length / width ratio (B) at different developmental stages of Aucuba chinensis fruits
启动期(1~4 MAW):此时的种子以胚珠的形式存在,胚珠和果皮之间有一个较大的空隙,大多数的果实只是在胚珠的中心对应胚乳的部位有一个小型的凹陷,尚未发育成胚乳。在体视显微镜下未能观察到胚乳。
生长期 Ⅰ(4 ~8 MAW):胚乳长度的增长略大于宽度,长度和宽度的平均月增长量分别为1.96 mm、1.67 mm,净增量占掉落时胚乳长度的45.29%、宽度的71.71%;这一阶段,胚乳长与宽的比值从4 MAW 的2.59 下降至8 MAW 的1.51。
停滞期(8~10 MAW):胚乳长度和宽度的平均月增长量分别为0.49 mm、0.24 mm;净增量占掉落时胚乳长度的11.27%、宽度的10.29%;胚乳长与宽的比值维持在1.5 左右。
生长期 Ⅱ(10 ~13 MAW):胚乳长度和宽度的平均月增长量分别为1.01 mm、0.11 mm,净增量占掉落时胚乳长度的23.30%、宽度的4.57%;胚乳长与宽的比值由1.50 增长至1.71。
2.4 发育期胚形态变化
2.4.1 胚生长与胚率变化
依据桃叶珊瑚种子的胚长度、宽度的变化曲线(图6),也可将其生长发育过程分为4 个阶段。

图6 桃叶珊瑚种子不同发育时期胚长度、宽度(A)以及胚率、胚长/宽比值(B)变化Fig. 6 Embryo length and width (A), embryo rate and ratio of embryo length to width (B) at different developmental stages of Aucuba chinensis seeds
启动期(1~5 MAW):体视显微镜下未能观察到种胚。
生长期Ⅰ(5~8 MAW):胚长度和宽度平均月增长量为0.57 mm 和0.16 mm;净增量占掉落时胚长度的40.67%、宽度的39.34%;由于长度增长明显高于宽度,胚的长宽比也从1.83 增至2.56;胚率由0.17增至0.27;该阶段与果实长度和宽度的生长期基本同步。
停滞期(8~12 MAW):胚长度和宽度的平均月增长量分别为0.06 mm、0.02 mm;净增量占掉落时胚长度的5.98%、宽度的5.74%;胚长与宽的比值从2.56 增长至2.93;胚率一直稳定在0.27~0.30,基本与果实发育同步,但这种缓慢生长持续到花冠凋落后的第12 MAW。
生长期Ⅱ(12~13 MAW):胚的发育进入第二个生长高峰期,虽然胚宽度变化不明显,但胚长度迅速增长,胚长度和宽度的平均月增长量分别为1.2 mm、0.14 mm;净增量占掉落时胚长度的28.71%、宽度的11.48%;胚长与宽的比值从2.93增长至3.84,形成较为明显的长条形。胚率再一次小幅提升,由0.30 增加到0.34;该阶段与果实发育期有部分重叠,但胚率的增长时间较短,然后果实掉落。
2.4.2 胚形态变化
桃叶珊瑚的胚胎发育属双子叶植物普遍的发育形式,都经历球形胚→心形胚→鱼雷形胚→子叶胚阶段(图4)。但不同形态胚的出现时间和不同胚的比例呈现出多态性(图7),鱼雷型胚持续时间长达9 个月,说明种胚发育缓慢。如图7 所示,生长期Ⅰ(5~8 MAW)期间,鱼雷型胚数量逐渐增多,占比由53%增至97%。停滞期(8~10 MAW),子叶胚自8 MAW开始出现后,球形胚与心形胚已消失,鱼雷型胚占比减少,但仍达90%以上。生长期Ⅱ(10~13 MAW),子叶胚占比迅速上升,但只达到23%,同期鱼雷形胚的占比则降为77%,说明桃叶珊瑚果实自然掉落时(13 MAW),种胚发育不完全。

图7 桃叶珊瑚果实发育过程不同种胚形态比例的变化Fig. 7 Changes in the proportion of embryo morphology of different species during fruit and seed development of Aucuba chinensis
2.5 环境因子与果实/种子形态变化的相关性
相关性分析表明,果形指数与月平均气温、平均降雨量均呈现显著正相关,相关系数分别为0.58和0.89 (图8 A、B),说明温度越高、降雨量越丰富越有利于桃叶珊瑚果实生长。其中降雨量对果实生长的影响达极显著水平(P<0.01),说明果实生长显著受降雨量的影响。胚率与平均温度、降雨量呈显著正相关(图8 C、D),说明胚的生长也要求较高的温度和降雨量。综上,桃叶珊瑚的果实和种子生长发育受水热条件影响较强烈。

图8 八大公山国家级自然保护区58 年(1961~2018 年)的月平均温度、降雨量与果实/种子生长发育的相关性分析Fig. 8 Correlation analysis of 58 years (1961-2018) monthly average temperature, rainfall and fruit / seed growth and development in Badagongshan National Nature Reserve
3 结论与讨论
3.1 桃叶珊瑚的花期与果期
研究表明,生长于八大公山的桃叶珊瑚果实发育缓慢,从花冠凋落到果实掉落持续了13 个月,在此期间出现明显的花果同期和果期重叠现象。另外,瘿椒树Tapisciasinensis[20]、海州常山Clerodendrum trichotomum[21]和油茶Camelliaoleifera[22]均具有该现象。麻力[20]对瘿椒树果期重叠现象的研究表明其功能是保障雌性功能,以及果实和种子的正常发育,但并未与生态因子关联起来。
在自然条件下,植物的花期、果期明显受到水热条件影响。邵周玲等[23]研究表明,四川米仓山区植物开花物候期随着水热条件的限制而明显推迟;三岛柴胡Bupleurumangustissimum[24]和新疆野杏Prunusarmeniaca[25]等的研究案例也表明水热条件是影响花期与果期的关键生态因子。基于此,我们对《中国植物志》记载的桃叶珊瑚属11 个种的花期、果期和原生地植被类型与气候特征进行整理(表1)[17]。从表1 可知,花期较早的是广西东兰的粗梗桃叶珊瑚A.robusta、香港的桃叶珊瑚和云南潞西、勐嘎的枇杷叶珊瑚A.eriobotryifolia,它们均处于水热条件更为丰沛的南亚热带常绿阔叶林下;花期较晚的是印度大吉岭的喜马拉雅珊瑚A.himalaica、浙江南部的青木A.japonica、湖北宜昌的倒心叶珊瑚A.obcordata、重庆南川金佛山的斑叶珊瑚A.albopunctifolia和湖南八大公山的桃叶珊瑚,均处于中亚热带常绿阔叶林下;同样果期最晚的也是处于中亚热带常绿阔叶林下的类群。这体现了桃叶珊瑚属植物开花结实的物候期随着水热条件亏缺而依次延迟的特性。的公开数据。

表1 桃叶珊瑚属植物的花、果期与生境特征Table 1 Floral, fruiting and habitat characteristics of Aucuba species
经比较发现,八大公山桃叶珊瑚的花期与果期较香港的桃叶珊瑚明显延迟,说明桃叶珊瑚开花结实的物候期随着纬度与海拔的升高而推迟,且受水热条件限制,进而形成了水热限制的时间窗口。这种水热窗口限制与板栗Castaneamollissima[26]、梨橙[27]等的相关研究结果相一致。
3.2 桃叶珊瑚果实与种子的生长发育特性
3.2.1 果实与种子发育的阶段性
不同植物的果实,其生长发育曲线类型不尽相同,但大体上可以分为两种,即单“S”型曲线和双“S”型曲线[19]。对桃叶珊瑚不同发育时期果实及种子长宽的测量结果表明,桃叶珊瑚果实的生长发育曲线属于典型的双“S”型,这与山茱萸Cornus officinalis[28]、杏Prunusarmeniaca[29]、李Prunus salicina[30]、油橄榄Canariumoleosum[31]等大多数核果类果实生长发育相似。虽然发育方式相同,但果实生长发育的最佳时期不同。桃叶珊瑚果实的2 个快速生长期分别为7~11 月(生长期Ⅰ)、翌年1 月~4 月(生长期Ⅱ)。而山茱萸果实的2 个快速生长期分别为4~6 月、7~8 月[28];杏为4~6 月初、6 月中旬~6 月下旬[29];杨梅果实为4~5 月、6 月[32]。由此可见,植物的果实发育不仅有其自身特性,也可能与生长区域及气候条件有关。桃叶珊瑚果实的第1 个快速生长期持续时间更长,且在该时期果实迅速膨大,因此说明生长期 Ⅰ(4 ~8 MAW)为果实生长发育的最佳时期。
桃叶珊瑚果实发育初期保持绿色,提示其叶绿素含量一直较高[33],但该过程蓝色色值的增加也提示类胡萝卜素的增长;果实成熟时变为红色,变红则主要是在红色色值稳定的情况下,绿色和蓝色色值迅速下降导致的,这似乎可归因于该时期叶绿素含量降至最低,而花青苷含量和类胡萝卜素含量显著增加[18]。
种胚生长与果实发育同步,生长期Ⅰ阶段仍是迅速生长的关键时期。同大多数双子叶植物一样,桃叶珊瑚种子的胚发育也经历球形、心形、鱼雷形和子叶型胚等阶段,但其果实成熟掉落时胚仍为鱼雷形胚(图7),有明显的发育迟滞特性。有研究发现,很多果实掉落前存在种胚形态发育迟滞的现象,比如箭叶淫羊藿Epimediumsagittatum种子胚发育长期停留在球形胚阶段[34],铁筷子Helleborus thibetanus[35]和羌活Hanseniaweberbaueriana[36]的种子也存在种胚形态发育不完全的现象。桃叶珊瑚种子掉落时的胚发育停滞必然导致其种子在萌发过程中具有明显的形态休眠,此后必然要经历形态后熟后才能在适宜的条件下萌发,这也与Carol 等[37]总结的具有形态休眠的科中包含桃叶珊瑚科的描述一致。
桃叶珊瑚胚乳的发育与果实形态发育基本同步,且无明显的浆状期,提示其发育类型属于细胞型[38-39]。大多数双子叶植物的胚乳作为营养物质的临时贮存库,会随着胚的发育逐渐被吸收和消耗[40—41]。譬如,叶立新[42]发现,山茱萸种子的胚乳在发育中被消耗。但是,本研究表明桃叶珊瑚果实成熟时胚乳仍然占据整个种子的绝大部分,与箭叶淫羊藿[34]、黄连Coptischinensis[43]类似,其保留的大量胚乳是为种子后熟提供营养支持的。
3.3 影响桃叶珊瑚果实与种子生长发育的环境因素
不同植物果实生长对水热的要求不同。许越等[44]发现降水增加会抑制槜李Prunussalicinavar.salicina果实的生长,这说明该植物的果实对水分的要求较低。而何璇[45]的研究表明,冬枣Ziziphus jujuba生长期需要较多水分,以便能为果实膨大提供充足的水分。在本研究中,桃叶珊瑚果实的发育与降水的相关性较高,即雨量丰沛季节其果形指数明显增加。当然,温度对果实发育也有显著影响。低温环境会使苹果MaluspumilaMill 停止生长并进入休眠[46],低温维持时间与桃Prunuspersica的果形指数呈显著负相关[47];相反,高温促进柑桔citrus reticulata的生长,低于适宜温度会使温州蜜柑Citrus reticulata'Unshiu'的膨大受到抑制[48]。在本研究中,高温显著促进桃叶珊瑚果形指数的增加。但就环境因素的相关性而言,降雨量的影响要强于温度。换言之,降雨量对桃叶珊瑚的果实发育更为关键。
一般认为,胚生长对温度的要求较高,大麦Hordeumvulgare[49]、人参Panaxginseng[50]等的种胚发育均会因温度不足而被抑制甚至停滞。在本研究中,相关分析结果表明,桃叶珊瑚种胚发育明显受到温度的调控,且体现为明显的正相关。值得指出的是,其发育过程的高温期(4~6 MAW)会迅速地促进胚的生长,但这种温度促进仅能使大部分胚发育进入鱼雷形胚时期,而其真正的完熟可能还需要一个高温期的促进才能完成。这种温度促进效应的分段现象与前面提及的水热条件限制形成生态限制因子窗口应当有密切的关联。
总之,桃叶珊瑚果实的生长要求高温适湿的环境,这可能与其长期适应常绿阔叶林下环境形成的阴生特性有关。因此,我们认为水热窗口期足够长,其果实和种胚很可能就会具备正常的花、果期,从而不会出现花果同期甚至果期重叠的现象,即影响种子发育从而导致种苗繁育困难的可能主要是水热窗口时间的长度。
同时,果实掉落时胚尚未完成分化,必然导致其种子具有形态休眠,并在种子脱离母体后经历后熟过程而发育完全。因此,为了更好地理解桃叶珊瑚种子的萌发特性,尚需进一步研究其种子在后熟过程中的胚形态与生理生化变化,以及胚形态变化的生态效应。
猜你喜欢
小小说月刊(2021年3期)2021-03-25
辽宁农业科学(2021年1期)2021-03-17
中国农业大学学报(2020年8期)2020-07-22
饮食保健(2020年6期)2020-04-03
西藏农业科技(2019年1期)2019-07-25
小学阅读指南·高年级版(2016年9期)2016-10-31
科学大众·小诺贝尔(2016年6期)2016-08-17
石油化工应用(2014年2期)2014-03-11
无机化学学报(2014年8期)2014-02-28
东北师大学报(自然科学版)(2014年1期)2014-02-27
