
低磷胁迫对山核桃幼苗根系形态和生理特征的影响
2010-12-05 02:49李永夫金松恒叶正钱黄坚钦姜培坤
浙江农林大学学报 2010年2期
李永夫,金松恒,叶正钱,黄坚钦,姜培坤
(1.浙江林学院 环境科技学院,浙江 临安 311300;2.浙江林学院-中国科学院南京土壤研究所森林土壤与环境联合实验室,浙江 临安 311300;3.浙江林学院 浙江省现代森林培育技术重点实验室,浙江 临安 311300)
缺磷是植物生长的主要限制因素之一[1]。以往的不少研究结果表明,植物在低磷胁迫条件下会呈现出一系列的适应性反应。例如,植物根变长、根毛数量和密度增大、形成排根等[2]以及根构型发生改变,从而有利于植物对土壤磷的吸收[3]。缺磷还会导致光合作用产物分配,从而使根冠比增加[2]。另外,叶片酸性磷酸酶活性和根系分泌酸性磷酸酶活性增加也是很多植物对低磷胁迫的反应之一[4]。因此,如何挖掘植物自身抵抗低磷胁迫的能力,为缓解土壤磷素生物有效性低下的问题提供了新的途径[5]。而深入系统研究植物对低磷胁迫的适应性机制对于能否有效应用上述新途径具有非常重要的意义。山核桃Carya cathayensis是中国特有的名优干果和木本油料作物,具有非常高的营养、保健、美容及药用价值[6]。近几年来,浙江省临安市山核桃生产迅速发展,也为当地农民的收入增加起到了重要的作用。初步调查发现,品质好的山核桃大部分都生长于石灰性母质发育的土壤上。而这种类型的土壤,养分(特别是磷养分)一般都比较贫瘠,并且养分的有效性也低。然而,在野生状态下,山核桃树营养状况正常。因此,我们可以推测,山核桃在适应低磷胁迫方面具有主动性适应机制。就目前调查的资料而言,关于山核桃根系形态和生理特性对低磷胁迫的反应方面的研究在国内外还鲜有报道。笔者以1年生山核桃幼苗为试验材料,通过溶液培养试验,研究低磷胁迫处理对山核桃根性形态、叶片光合作用特性以及叶片和根系分泌酸性磷酸酶活性的影响,为提高山核桃磷养分的利用效率提供依据。
1 材料与方法
1.1 供试材料
本研究植物材料为1年生山核桃苗。
1.2 试验设计和方法
水培试验于2007年3-6月在浙江林学院人工智能气候室内进行。营养液按照霍格兰营养液作一定调整后配制。营养液配方如下:Ca(NO3)2·4H2O 2.00 mmol·L-1,KH2PO40.10 mmol·L-1,MgSO4·7H2O 0.50 mmol·L-1,KCl 0.10 mmol·L-1,K2SO40.70 mmol·L-1,H3BO310.00·mol·L-1,MnSO4·H2O 0.50 μmol·L-1,ZnSO4·7H2O 1.00 μmol·L-1,CuSO4·5H2O 0.20 μmol·L-1,(NH4)6Mo7O240.01 μmol·L-1,Fe-EDTA 100 μmol·L-1。供试山核桃植株取自山核桃栽培基地,运回实验室后用自来水冲洗干净,选择大小均一的山核桃幼苗移植到含有36 L正常培养液的塑料培养槽中。培养2周后(山核桃达1片真叶时)按照以下处理继续培养:试验设 2 个供磷水平:0.02 mmol·L-1(低磷处理),0.10 mmol·L-1(正常供磷处理)。磷以KH2PO4形态供应。试验设3个重复。在处理后的培养过程中每天用pH计测定营养液pH值,并用 0.1 mol·L-1氢氧化钠或 0.1 mol·L-1氯化氢调节营养液至 pH 5.8,保持 24 h连续通气,6 d换1次营养液。连续处理36 d后收获。
1.3 样品处理和测定方法
1.3.1 气体交换参数测定 山核桃叶片的气体交换参数用Li-6400红蓝光源 6400-02(Li-Cor公司,USA)测定。该仪器可以原位测定叶片的光合速率、胞间二氧化碳摩尔分数、气孔导度等参数。测定的叶片为山核桃的第1片完全展开叶。时间为不同磷处理后第28天上午8∶30-10∶00。叶室设定参数如下:温度为 28 °C,二氧化碳摩尔分数为 390 μmol·mol-1,光合有效辐射(PAR)为 1 000 μmol·m-2·s-1。
1.3.2 根系形态 收获植株后,先用自来水冲洗干净,然后立即将根系样品保存于体积分数为25%的乙醇溶液中。采用 Win-Rhizo(Regent Instruments,Canada)根系分析仪对根系根长、根表面积和根体积进行定量分析。
1.3.3 酸性磷酸酶活性 山核桃幼苗叶片酸性磷酸酶活性的测定参照 McLachlan等[7]的方法略作改动。取山核桃幼苗的第一完全展开叶,去掉叶基和叶尖部分,称取一定质量的鲜样(0.1~0.2 g)后,快速用液氮冷冻。样品加入5.00 mL浓度为0.20 mol·L-1的冰醋酸-醋酸钠缓冲液(pH 5.8),在冰浴中研磨匀浆,将匀浆溶液转入10.00 mL离心管,27 000×g离心10 min(4°C)。上清液为酶测定液。测定反应如下:0.05 mL酶液 +0.45 mL缓冲液 +4.50 mL 5 mmol·L-1对硝基苯酚磷酸盐,30°C黑暗中保温30 min后,加2.00 mL 2.00 mol·L-1氢氧化钠中止反应。在405 nm处测定吸光度值,以无酶反应作空白调0。同时作对硝基苯酚的标准曲线,根据标准曲线算出反应体系中生成的对硝基苯酚的量。山核桃幼苗叶片酸性磷酸酶活性以单位时间单位鲜质量所生成的对硝基苯酚的微摩尔数来表示。山核桃幼苗根系分泌酸性磷酸酶活性的测定参考 Ni等[8]的方法。取正常供磷处理和低磷处理的山核桃幼苗各1株,用去离子水将山核桃根系洗干净,然后将山核桃植株移至盛有500.00 mL营养液的三角瓶中(正常供磷处理和低磷处理的山核桃植株分别移至正常营养液和低磷营养液中,营养液中对硝基苯酚磷酸二钠的浓度均为1.00 mmol·L-1)。三角瓶外面用2层黑色塑料薄膜包住。以不培养植株作为对照。正常光温条件下培养2 h,吸取1.00 mL反应液加入到含5.00 mL 1.00 mol·L-1氢氧化钠的试管中,摇匀后在405 nm处测定吸光度值,以无酶反应作空白调0。试验结束后,剪取根系,称其鲜质量。山核桃幼苗根系分泌酸性磷酸酶活性以单位时间单位根鲜质量所生成的对硝基苯酚微摩尔数来表示。1.3.4 根系干质量和磷质量分数 地上部和地下部样品用去离子水冲洗几遍。将鲜样先在105℃下杀青30 min,再在70℃下烘48 h(至恒量),测定其干物质量。再用万能不锈钢粉碎机磨细,过60目筛,放在封口袋里,供矿质元素分析测定用。植物样品用浓硫酸-过氧化氢消煮,采用钼兰比色法测植株全磷[9]。
1.4 数据统计
数据进行方差分析,并用新复极差法进行多重比较。数据的统计分析均在Microsoft Excel和SAS 8.0软件上进行。
2 结果与分析
2.1 低磷胁迫对山核桃植株干质量和根冠比的影响
从图1可以看出,与正常供磷相比,低磷胁迫处理显著降低山核桃植株的干重(P<0.05)。低磷胁迫条件下的根干质量为正常供磷条件下的74.7%(图1-A);低磷胁迫下的植株地上部干质量为正常供磷的53.2%(图1-B)。低磷胁迫显著增加山核桃植株的根冠比(P<0.05)(图1-C)。
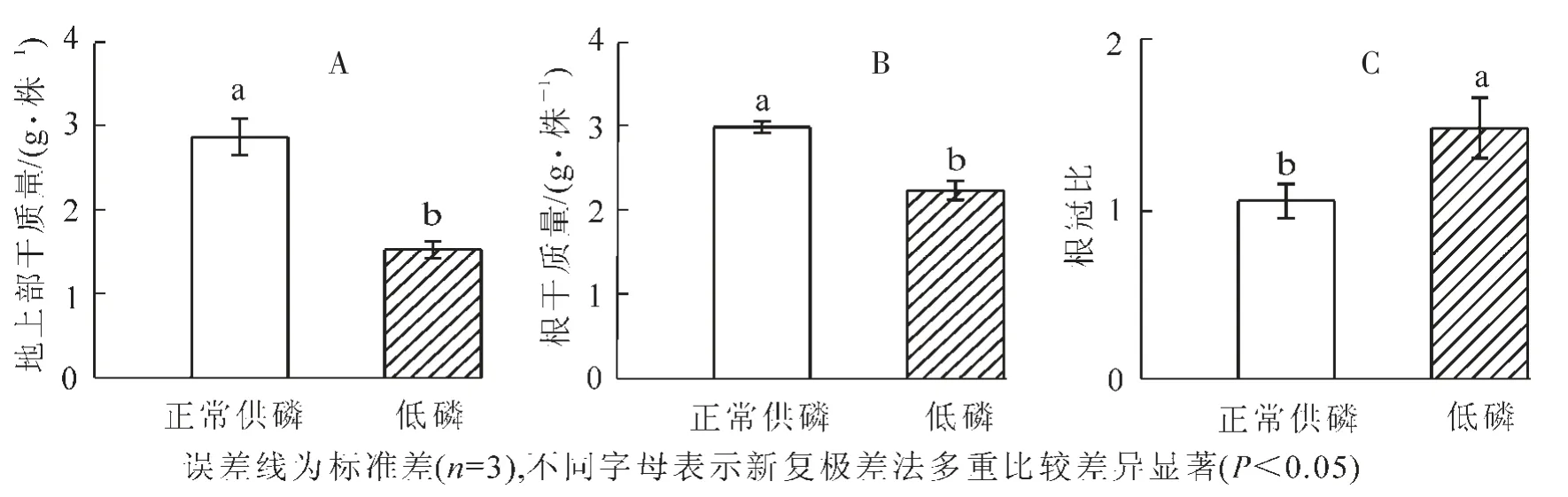
图1 低磷处理对山核桃干物质量(A,B)和根冠比(C)的影响Figure 1 Effect of low P treatment on the dry weight and root/shoot dry weight ratio of hickory
2.2 低磷胁迫对山核桃磷营养特性的影响
从表1可见,低磷处理显著降低山核桃植物的磷质量分数和磷积累量。与正常供磷相比,低磷处理条件下山核桃地上部和根系磷质量分数分别下降81.1%和79.4%;与正常供磷相比,低磷处理条件下山核桃地上部和根系磷积累量分别下降89.1%和85.9%。与正常供磷条件相比,低磷处理显著增加磷在根部的分布(P<0.05)(表1)。
2.3 低磷胁迫对山核桃根系形态特性的影响
从图2可见,低磷胁迫对于山核桃的根系形态具有非常显著的影响。与正常供磷相比,低磷处理显著降低山核桃的根长、根表面积和根体积(P<0.01)。低磷处理条件下山核桃根长、根表面积和根体积分别为正常供磷条件下的66.1%,66.0%和72.7%。

表1 低磷处理对山核桃地上部和根系的磷质量分数和磷积累量的影响Table 1 Effect of low P treatment on the concentration and accumulation of P in shoots and roots of hickory

图2 低磷处理对山核桃根长(A),根表面积(B)和根体积(C)的影响Figure 2 Effect of low P treatment on root length(A),root surface area(B),and root volume(C)of hickory seedlings
2.4 低磷胁迫对山核桃叶片光合作用特性的影响
从图3可见,低磷处理显著降低山核桃叶片的光合速率和气孔导度(P<0.05)。与正常供磷的植株相比,低磷处理使山核桃叶片光合速率下降30.3%(图3-B)。但是对叶片胞间二氧化碳摩尔分数没有显著的影响(图3-C)。

图3 低磷处理对山核桃叶片光合速率(A),气孔导度(B)和胞间二氧化碳(C)的影响Figure 3 Effect of low P treatment on photosynthetic rate(A),stomatal conductance(B),intercellular CO2concentration(C)in the leaf of hickory
2.5 低磷胁迫对山核桃叶片和根系分泌酸性磷酸酶活性的影响
正常供磷和低磷处理条件下山核桃叶片酸性磷酸酶活性和根系分泌酸性磷酸酶活性如图4所示。低磷胁迫使山核桃叶片酸性磷酸酶活性和根系分泌酸性磷酸酶活性显著增加(P<0.05),分别为正常供磷处理下的135%和159%。
3 讨论
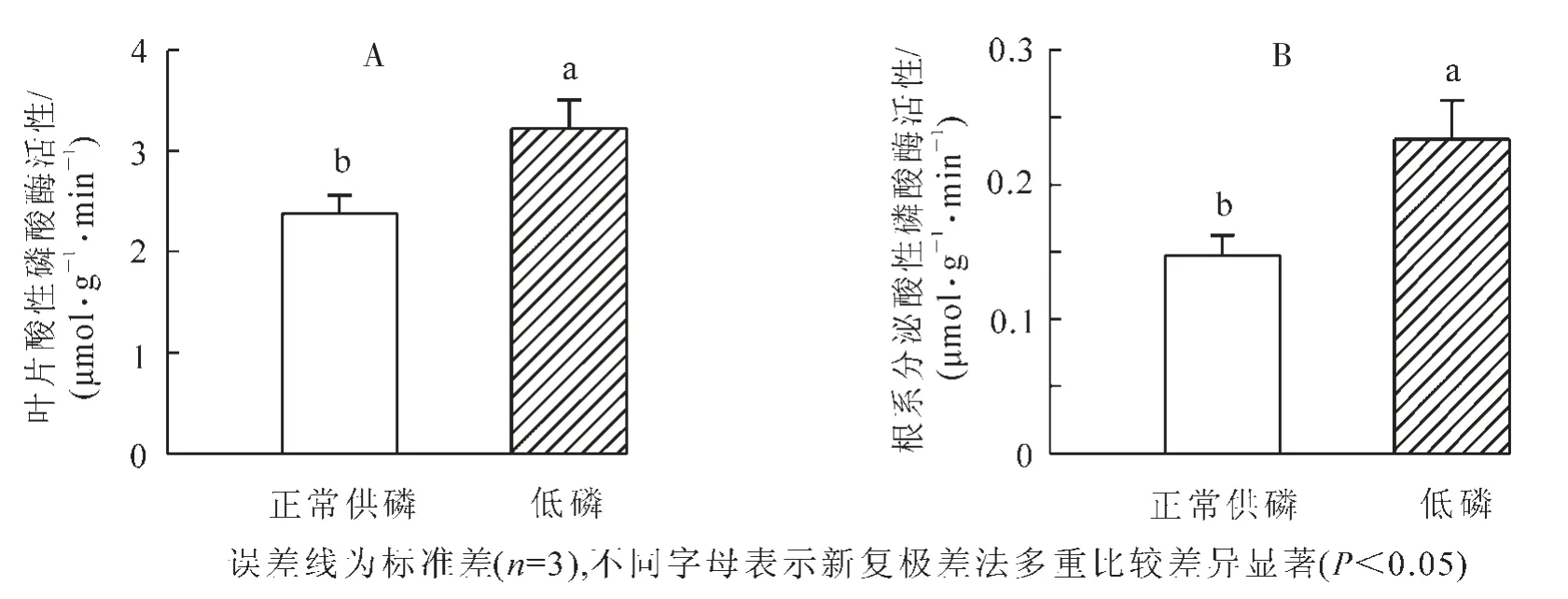
图4 低磷处理对山核桃叶片酸性磷酸酶活性(A)和根系分泌酸性磷酸酶活性(B)的影响Figure 4 Effect of low P treatment on leaf acid phosphotase activity(A)and root secreted acid phosphotase activity(B)in hickory
以往的大量结果表明,植物的生长发育以及产量会在很大程度上受到磷供应水平的影响[2]。本研究结果表明,低磷处理显著降低山核桃植株幼苗地上部和根系的干质量,但是两者降低的幅度不同。低磷处理对地上部植株干质量的影响要显著高于对根系干质量的影响。另外,低磷处理显著增加山核桃植株的根冠比。这跟其他植物如水稻 Oryza sativa[10],大豆 Glycine max[11],马尾松 Pinus massoniana[12],水曲柳Fraxinus mandshurica[13]等的报道相似。上述结果表明,低磷处理条件下,尽管地上部和根系干质量都显著下降,但是植物受到低磷胁迫时,会首先保证根系的生长。也有研究报道,低磷处理条件下地上部干物质质量显著降低,而根系干质量非但没有降低甚至还有少许增加[14]。从本研究根冠比的结果也可以说明,低磷处理条件下,山核桃会分配更多的干物质到根系,从而优先保证根系的生长,使根系能够尽可能地吸收养分,从而减少植物生长受到低磷胁迫的影响。
大量研究结果表明,植物根系形态特征和磷素养分的吸收有着非常密切关系[15]。例如,Newman等[16]报道小麦Triticum aestivum的吸磷量与根系总长度呈显著正相关。李海波等[2]的研究结果表明低磷条件下,根系总表面积、侧根总长度以及侧根数量均与水稻的吸磷量存在显著正相关。Borkert等[17]的研究结果表明,大豆吸磷量只与根系总表面积呈显著正相关,而与根长、平均根轴和根干质量等都没有显著相关性。这表明,植物的根系形态会与植物吸收磷素养分具有非常密切关系。同时,以往不少研究还发现,低磷胁迫会诱导根系形态发生变化(如根变长、根毛数量和密度增大等)[15]。上述这些变化无疑会促进植物对磷素的吸收。而在本研究结果中,我们并没有发现低磷处理增加山核桃根长或者根表面积的现象。这可能与低磷胁迫的程度以及植物种类有关。一般来讲,当植株受到轻度低磷胁迫的时候,植物会出现根长变长,侧根增加的现象。然而,随着低磷胁迫程度的加深,植株整个根系的生长可能会受到影响,从而使植株根系的各个形态特征(如根长、根表面积和根体积)都有显著下降。因此,从本研究结果来看,山核桃的根系形态反应不是山核桃适应低磷胁迫的主要机制之一。
低磷处理显著降低植株叶片光合速率在C3和C4植物上均有报道[18-19]。本研究结果表明,低磷处理导致山核桃植株的叶片光合速率下降33.3%。一般来讲,导致光合速率下降主要有2个因素:①气孔因素,主要受气孔数量、气孔孔径和气孔开度等的影响[20]。②非气孔因素,主要受内部的酶活力和光合组分控制。而在以往的研究中,低磷处理影响植物叶片光合速率的机制既包括气孔因素又包括非气孔因素[18-19]。本研究结果表明,低磷处理显著降低山核桃植株的气孔导度,但对山核桃叶片胞间二氧化碳摩尔分数并没有显著的影响。这表明低磷胁迫对山核桃叶片光合速率的抑制作用主要是由于非气孔因素引起的。Fredeen等[18]的研究结果也表明,磷缺乏引起菜豆Phaseolus vulgaris光合速率的下降主要是由非气孔因素引起的。由于磷缺乏造成叶片光合速率下降的影响因子较多,因此,还需在以后的试验中作深入研究。
低磷胁迫下植物根系分泌酸性磷酸酶活性的增加已经被许多研究学者认为是植物适应低磷胁迫的机制之一[4]。以往的研究结果表明,玉米 Zea mays[21]和羽扇豆 Lupinus micranthus[22]受到低磷胁迫时,根系分泌酸性磷酸酶活性显著增加。本研究的结果表明,与正常供磷的植株相比,低磷胁迫使山核桃根系分泌酸性磷酸酶活性增加了59%。由于有机磷是土壤磷素的重要组成部分[23],因此,根系分泌磷酸酶活性的增加将促进土壤中有机磷的水解,从而提高土壤中磷的生物有效性。如Asmar等[24]的研究结果表明5个大麦Hordeum vulgare基因型根际NaHCO3-Po的亏缺程度与根系分泌的酸性磷酸酶活性呈显著的正相关。另外,从本研究的结果表明,低磷处理显著增加了山核桃叶片酸性磷酸酶活性,这与以往在其他植物如玉米[21,25]、 小麦[26]、 菜豆[27]和大豆[28]上的研究结果基本一致。植物叶片酸性磷酸酶活性的提高,可以促进植物利用有机磷,从而提高植物的磷利用效率。因此,低磷处理条件下,根系分泌酸性磷酸酶和叶片酸性磷酸酶活性的提高可能是山核桃适应低磷胁迫的机制之一。
[1]HINSINGER P.Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes:a review[J].Plant Soil,2001,237:173-195.
[2]李海波,夏铭,吴平.低磷胁迫对水稻苗期侧根生长及养分吸收的影响[J].植物学报,2001,43(11):1154-1160.LI Haibo,XIA Min,WU Ping.Effect of phosphorus deficiency stress on rice lateral root growth and nutrient absorption[J].Acta Bot Sin,2001,43(11):1154-1160.
[3]LIAO Hong,YAN Xiaolong.Adaptive changes and genotypic variation for root architecture of common bean in response to phosphorus deficiency[J].Acta Bot Sin,2000,42(2):158-163.
[4]VANCE C P,UHDE-STONE C,ALLAN D L.Phosphorus acquisition and use:critical adaptations by plants for securing a nonrenewable resource[J].New Phytol,2003,157:423-447.
[5]戴开杰,沈有信,周文君,等.在控制条件下云南松幼苗根系对低磷胁迫的相应[J].生态学报,2005,25(9):2423-2426.DAI Kaijie,SHEN Youxin,ZHOU Wenjun,et al.Mechanism of Pinus yunnanensis seedlings root response to phosphorus deficiency under controlled conditions[J].Acta Ecol Sin,2005,25(9):2423-2426.
[6]钱新标,徐温新,张圆圆,等.山核桃果仁微量元素分析初报[J].浙江林学院学报,2009,26(4):511-515.QIAN Xinbiao,XU Wenxin,ZHANG Yuanyuan,et al.Trace elements in kernels of Chinese hickory(Carya cathayensis)grown in limestone and noh-limestone soils[J].J Zhejiang For Coll,2009,26(4):511-515.
[7]MCLACHLAN K D,ELLIOTT D E,De MARCO D G,et al.Leaf acid phosphatase isozymes in the diagnosis of phosphorus status in field-grown wheat[J].Aust J Agr Res,1987,38:1-13.
[8]NI Junjian,WU Ping,LUO Ancheng,et al.Low phosphorus effects of the metabolism on rice seedlings[J].Commun Soil Sci Plan Anal,1996,27:3073-3084.
[9]鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,1999:312-314.
[10]李永夫,罗安程,黄继德,等.不同磷效率水稻基因型根系形态和生理特性的研究[J].浙江大学学报:农业与生命科学版,2006,32(6):658-664.LI Yongfu,LUO Ancheng,HUANG Jide,et al.Study on the morphological and physiological characteristics of roots in rice genotypes with contrasting P efficiency[J].J Zhejiang Univ Agric & Life Sci,2006,32(6):658-664.
[11]CAKMAK I,HENGELER C,MARSCHNER H.Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus,potassium and magnesium deficiency[J].J Exp Bot,1994,45:1245-1250.
[12]徐向华,丁贵杰.马尾松适应低磷胁迫的生理生化响应[J].林业科学,2006,42(9):24-28.XU Xianghua,DING Guijie.Physiological and biochemical responses of Pinus massoniana to low phosphorus stress[J].Sci Silv Sin,2006,42(9):24-28.
[13]吴楚,范志强,王政权.氮磷供应状态对水曲柳幼苗氮磷吸收与生长的影响[J].林业科学,2005,41(5):196-200.WU Chu,FAN Zhiqiang,WANG Zhengquan.Influences of nitrogen and phosphorus supply on their absorption and growth of Fraxinus mandshurica seedlings[J].Sci Silv Sin,2005,41(5):196-200.
[14]MING Feng,MI Guohua,ZHANG Fusuo,et al.Differential response of rice plants to low-phophorus stress and its physiological adaptive mechanism[J].J Plant Nutr,2002,25:1213-1224.
[15]RAGHOTHAMA K G.Phosphate acquisition[J].Ann Rev Plant Physiol Plant Mol Biol,1999,50:665-693.
[16]NEWMAN E I,ANDREWA R E.Uptake of phosphorus and potassium in relation to root growth and root density[J].Plant Soil,1973,38:49-69.
[17]BORKERT C M,BARBER S A.Effect of supplying P to a portion of the soybean root system on root growth and P up-take kinetics[J].J Plant Nutr,1983,6:685-910.
[18]FREDEEN A L,RAO I M,TERRY N.Influence of phosphorus nutrition on growth and carbon partitioning in Glycine max[J].Plant Physiol,1989,89:225-230.
[19]JACOB J,LAWLOR D W.Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower,maize and wheat plants[J].J Exp Bot,1991,42:1003-1011.
[20]QUICK W P,CHAVES M M,WENDLER R,et al.The effect of water stress on photosynthetic carbon metabolism in four species grown under field conditions[J].Plant Cell Environ,1992,15:25-35.
[21]YUN S J,KAEPPLER S M.Induction of maize acid phosphatase activities under phosphorus starvation[J].Plant Soil,2001,237:109-115.
[22]WASAKI J,YAMAMURA T,SHINANO T,et al.Secreted acid phosphatase is expressed in cluster roots of lupin in response to phosphorus deficiency[J].Plant Soil,2003,248:129-136.
[23]DALAL R C.Soil organic phosphorus[J].Adv Agron,1997,29:83-117.
[24]ASMAR F,GAHOONIA T S,NIELSEN N E.Barley genotypes differ in activity of soluble extracellular phosphatase and depletion of organic phosphorus in the rhizosphere soil[J].Plant Soil,1995,172(1):117-122.
[25]GEORGE C E,LAUCHLI A.Evaluation of an acid phosphatase assay for detection of phosphorus deficiency in leaves of maize(Zea mays L.)[J].J Plant Nutr,1986,9:1469-1477.
[26]MCLACHLAN K D.Effects of drought,aging and phosphorus status on leaf acid phosphatase activity in wheat[J].Aust J Agric Res,1984,35:777-787.
[27]YAN Xiaolong,LIAO Hong,MELANIE C,et al.Induction of a major leaf acid phosphatase does not confer adaptation to low phosphorus availability in common bean[J].Plant Physiol,2001,125:1901-1911.
[28]TIAN Jiang,LIAO Hong,WANG Xiurong,et al.Phosphorus starvation-induced expression of leaf acid phosphatase isoforms in soybean[J].Acta Bot Sin,2003,45:1037-1042.
猜你喜欢
作物学报(2022年3期)2022-01-22
数学大王·中高年级(2021年3期)2021-03-10
分析化学(2018年7期)2018-09-17
阅读(中年级)(2018年11期)2018-01-19
中国组织化学与细胞化学杂志(2017年1期)2017-06-15
中国医学科学院学报(2013年6期)2013-03-11
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
中国新闻周刊(2004年11期)2004-04-07
