
垫状锦鸡儿的繁殖方式及其在适应干旱荒漠环境中的生态意义
2011-01-05 08:15张建华马成仓李清芳乌云其其格高玉葆
天津师范大学学报(自然科学版) 2011年4期
张建华,马成仓,李清芳,乌云其其格,高玉葆,2
(1.天津师范大学a.生命科学学院,b.天津市细胞遗传与分子调控重点实验室,天津 300387;2.南开大学 生命科学学院,天津300071;3.锡林浩特市草原工作站,内蒙古 锡林浩特 026000)
垫状锦鸡儿的繁殖方式及其在适应干旱荒漠环境中的生态意义
张建华1,马成仓1,李清芳1,乌云其其格3,高玉葆1,2
(1.天津师范大学a.生命科学学院,b.天津市细胞遗传与分子调控重点实验室,天津 300387;2.南开大学 生命科学学院,天津300071;3.锡林浩特市草原工作站,内蒙古 锡林浩特 026000)
研究内蒙古荒漠区锦鸡儿属优势种垫状锦鸡儿灌丛结构、分株特征、繁殖倾向和克隆生长特性.结果表明:在荒漠区,垫状锦鸡儿的营养繁殖占主导地位,有性繁殖为辅;其营养繁殖由枝条克隆产生无性系分株,克隆生长构型为密集型.较高的无性繁殖比例、密集型的克隆构型和发达的根系使垫状锦鸡儿能很好地适应荒漠区的干旱环境,并成为荒漠区的优势种.
垫状锦鸡儿;无性繁殖;干旱荒漠区;适应性
锦鸡儿属(Caragana)植物属于豆科,全世界有100余种[1],主要分布于亚洲和欧洲的干旱和半干旱地区[2],为落叶灌木,以其强抗旱性而著称[3].垫状锦鸡儿是锦鸡儿属的重要物种,分布于我国的内蒙古、宁夏、甘肃、青海、四川、西藏和蒙古国等地[4].由于它对荒漠气候的适应使其成为荒漠区的优势种,是荒漠区防风固沙、保持水土的优良植物.另外,它还具有饲用、绿肥、薪炭、蜜源和入药等资源价值.文献已经报道了垫状锦鸡儿对干旱荒漠环境的形态[5]和生理适应对策[6-7],研究表明:垫状锦鸡儿通过叶片的形态结构、高水分利用效率、强保水能力和维持稳定的水分等方式来适应荒漠环境.繁殖是植物生活史的重要部分,成功繁殖是植物在特定环境定居的标志,许多研究都表明植物通过繁殖对策的改变而适应不同环境[8-12],可见,植物的繁殖对策对环境的适应具有重要意义.关于垫状锦鸡儿繁殖方式尚未见报道.为此,针对内蒙古干旱荒漠区垫状锦鸡儿集群的灌丛结构、繁殖倾向和克隆生长特性进行调查,旨在揭示垫状锦鸡儿的繁殖特点及其繁殖对策在适应荒漠环境中的生态意义,为有效利用该植物资源,发挥其在防风固沙、保持水土中的作用提供科学依据.
1 研究地区与研究方法
1.1 研究地区概况
研究地点位于内蒙古高原西部的阿拉善左旗,地 理 位 置 为 东 经 105.66°,北 纬 38.84°,海 拔1 561.0 m.其年降水量110 mm,平均气温7.80℃,≥10℃年积温3 250℃,日照时间3 200 h,总辐射量625.0 kJ·cm-2,土壤含水质量分数为1.73%(30 cm,1.66%;60 cm,1.55%;100 cm,1.98%)[5].境内植被稀疏,植物种类贫乏,盖度低,一般只有1%~20%,以旱生、超旱生和盐生的灌木、半灌木、小灌木为主要建群植物[13].垫状锦鸡儿是该区的优势种之一.
1.2 实验方法
2009年7月在研究地区随机选取5个垫状锦鸡儿集群,先用卷尺测量各集群的面积和高度,接着用挖掘工具先小心地去掉沙包,再由地表向下挖掘一壕沟,采用分层剥离土壤的办法,将植株的地上和地下部分暴露出来,观察植株地上部分的形态并测量植株的高度;仔细观察枝条被沙埋后不定根的生长情况和每个单株的无性系分株情况;统计集群中的实生苗(单株)和无性系分株的数目并以此计算有性繁殖和无性繁殖的比例;观察根的空间走向,测量母株侧根和分株不定根分布的深度范围和直径;绘集群中单株分布、克隆分株的不定根根系和主根根系平面图(本文详细报告3个典型的集群).
2 结果与分析
2.1 集群的生长和分株特征
2.1.1 集群1
集群1所占据的空间为一个圆,周长为4.5 m,面积约1.6 m2,挖掘前从外形观察是一个枝条短而密、呈半球形、贴伏于地表的单株灌丛,高度为43 cm(包含沙包高度),沙包高约28 cm.其克隆生长构型见图1.
从图1可以看到,垫状锦鸡儿贴近地面的枝条或沙埋枝条可以产生不定根,形成无性系分株,可见垫状锦鸡儿属于枝源型克隆植物.垫状锦鸡儿母株的根系为轴根型,由主根和侧根构成,根的颜色为土黄色.主根上半部分粗壮(直径平均在4~6 cm),多弯曲,一直向下延伸,深度可达70 cm以上,它保证了垫状锦鸡儿在大气及土壤干旱的条件下吸收到充足的水分.母株的侧根多数由主根入土后10~20 cm处发出,分枝少,其中一级侧根较粗壮.垫状锦鸡儿根的入土深度比地上部分高度大2倍以上,根幅大于冠幅.
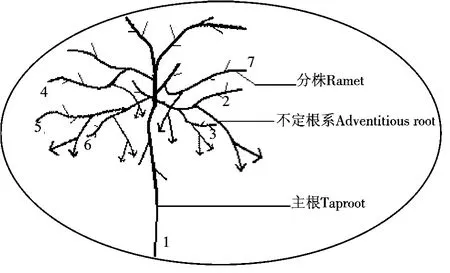
图1 垫状锦鸡儿集群1的生长构型Figure 1 Growth pattern of C.tibetica cluster 1
集群1中共产生6个子株,编号为2,3,4,5,6和7.新产生的分株与母株之间的间隔子长6~8 cm左右,它们紧密相连,交错生长在一起.此外,这些分株的地上枝条相互近于平行,角度约为0,数量较多.各级分株都具有发达的不定根,直径有0.8 cm,1.0 cm和1.5 cm不等.不定根分布幅度宽,入土深度多数在近地表至40 cm土层中,以水平和一定的锐角分布,最长可达80 cm.这些不定根在向四周延伸的过程中,会产生分枝,在土层中以一定的锐角分布.分株的不定根系深插入土壤中,有效地增加了垫状锦鸡儿地下部分的深度、广度和生物量,增强了垫状锦鸡儿根系吸收水分和养分的能力,从而引起地上部分的覆盖面积和生物量的增加.可见,这种分株系统的结构,在“搜寻”光能、吸收水分和养分、避免干旱方面有着非常强的适应性.集群1中未发现无性系分株与母株分离的现象.
2.1.2 集群2
此集群所占的空间为一个椭圆,周长为4.4 m,面积约1.54 m2,挖掘前从外貌看,是一个连续密集生长的灌丛.挖掘后发现,其内有2株垫状锦鸡儿,植株1高为42 cm,植株2的高度为41 cm,沙包高度约为27 cm,生长构型如图2所示,实物图如照片A.从照片A可以看到,植株1(树龄大)地上枝条较多,当年生枝条达50多根,其主根和各级侧根在延伸长度和粗度上均占有明显的优势.沙埋枝条产生的无性系分株较多,其不定根的数量较多,直径较粗.植株2树龄较小,地上枝条较少,产生的无性系分株少,不定根的数量少,直径小.植株1产生6个子株,编号为3,4,5,6,7和8.植株2产生1个分株,编号为9.在集群2中,尚未发现克隆分株与母株分离的现象.但是,观察到有些无性系分株间的结合部位出现部分断裂现象,断面(茎结构)中的髓部及其周围的木质部颜色发黑.

照片A
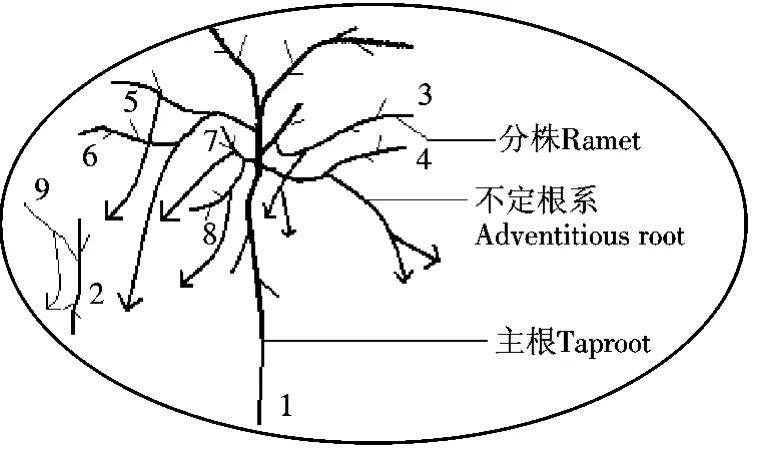
图2 垫状锦鸡儿集群2的生长构型Figure 2 Growth pattern of C.tibetica cluster 2
2.1.3 集群3
此集群所占的空间为一椭圆,其周长为6 m,面积约2.8 m2.挖掘前从外形观察,是一个连续紧密生长的灌丛.挖掘后发现,其内有3株垫状锦鸡儿.它们的生长构型如图3.植株1的株高为40 cm,树龄最大,地上部分和地下部分的生物量都最大,结构也最复杂.植株2和3的树龄较小,生物量较小,结构较简单.3植株都有一明显的主根,主根上半部分粗壮,多弯曲,深插入土壤中,侧根较少.3株垫状锦鸡儿通过沙埋枝条进行繁殖,植株1产生3个子株,编号为4,5和6.植株2和3分别产生1个子株,编号分别为7和8(带箭头的折线表示不定根系),但未发现分株与母株分离的现象.
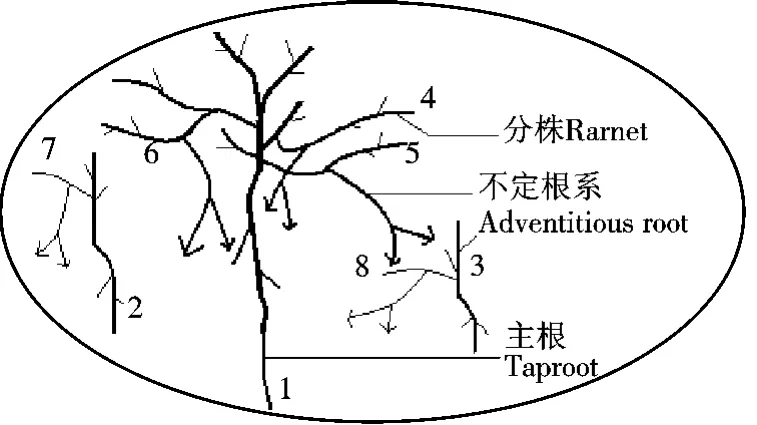
图3 垫状锦鸡儿集群3的生长构型Figure 3 Growth pattern of C.tibetica cluster 3
2.2 垫状锦鸡儿的繁殖方式
将5个集群中的实生苗和无性系分株分别进行统计,结果见表1.从表1看到:荒漠区垫状锦鸡儿既可以进行无性繁殖,又可以进行有性繁殖,且无性繁殖所占的比例(79.5%)明显高于有性繁殖(20.5%).
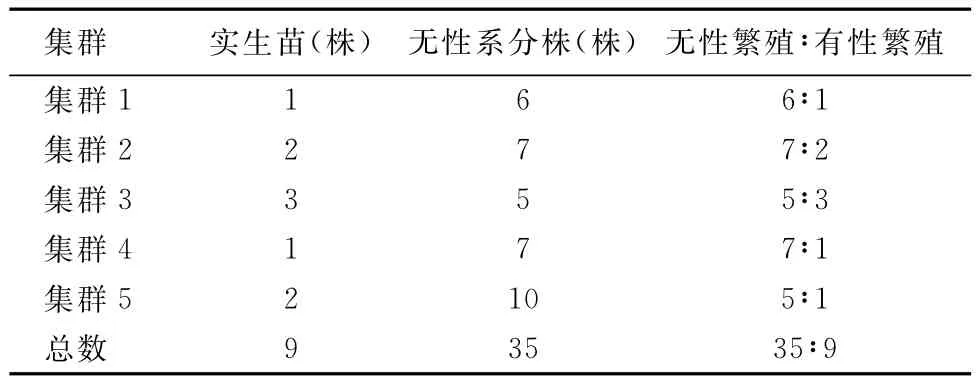
表1 垫状锦鸡儿集群的实生苗和无性系分株比例Table 1 Proportion of seedlings and ramets of C.tibetica clusters
3 讨论和结论
植物在有性繁殖与无性繁殖间存在着权衡,在某一系统中植物采取何种方式与系统的历史、干扰程度和立地条件等有关[14].一般认为,无性繁殖可以使植物在种苗更新困难的条件下维持局域种群的存活[15].在极端条件下(如干旱、高寒),无性繁殖具有优势[16-17].本研究表明,内蒙古阿拉善荒漠区的垫状锦鸡儿以克隆繁殖为主.
在极端干旱的内蒙古西部,垫状锦鸡儿结实率低,成熟种子离开母体灌丛因干旱而难以萌发,即使萌发了,幼小个体也不能适应极端干旱的环境和经受牛羊的践踏,故难以存活.垫状锦鸡儿几乎无法靠有性繁殖来延续种族,通过克隆繁殖新产生的分株与母株借助间隔子相互连接,从而极大地提高了其成活率.因此,在干旱和重度动物胁迫的荒漠区环境中,无性繁殖是垫状锦鸡儿的有效更新对策.克隆生长和克隆分株的不断出现,使垫状锦鸡儿基株实现在时空上的散布,迅速扩大种群数量,增加对地表的覆盖,在沙埋胁迫的逆境中承受风险,获得生存并固定了地表基质,进一步改善了自己的生存条件.可见这种繁殖对策的选择是其对荒漠环境的适应策略之一.
赵晓英等[18]的研究表明,在干旱地区,中间锦鸡儿以种子更新为主,根化枝条更新为辅,白毛锦鸡儿和甘蒙锦鸡儿以克隆生长为主,种子更新为辅.牛西午[19]的研究也表明,柠条的枝条被沙埋后能从枝上产生不定根,并从上部萌生出许多新的枝条,其萌生和再生能力极强.闫兴富[20]的调查也发现,在长期的干旱胁迫下,柠条的一种适应方式就是发挥营养繁殖的优势.本研究和前人的这些研究结果说明锦鸡儿属植物通过调整繁殖方式来适应环境.
克隆构型是由克隆生长过程中3个形态学性状决定的,即间隔子长度、分枝强度和分枝角度.间隔子长,分枝少,并不断沿原方向生长,在水平方向迅速扩展,形成近直线型的子代分株链,具有这种生长格局的植物被称为“游击型”克隆植物.相反,间隔子短,分枝强度强,但方向性差,扩展慢,形成不规则圆形株丛,具有这种生长格局的植物被称为“密集型”克隆植物[21-22].不同的克隆构型对应着不同的生态对策:密集型克隆植物表现出“巩固”对策;游击型克隆植物表现出“觅食”对策或“保守”对策[23].
研究发现,荒漠区垫状锦鸡儿主要是通过沙埋枝条产生不定根形成克隆分株,并且新产生的分株与母株之间的间隔子都比较短,一般在6~8 cm左右,它们形体紧密相连,交错生长在一起,在形态建成上形成一个面积可达2.8 m2的不规则圆形株丛,所以荒漠区垫状锦鸡儿克隆生长构型为密集型.这与另一类克隆植物(克隆器官为枝条和根条)如绵刺[24]、花棒[25]、四合木[26]在资源贫乏的环境中通过枝条繁殖形成的游击型克隆构型不同.造成这种差异的原因在于前者的分株直接由母株枝条产生,故它们紧密相连在一起,而后者产生的分株生长到一定的阶段,其枝条逐渐被沙埋,又进行克隆生长形成次级克隆体,造成分株间的距离(间隔子)较长,分株比较分散.可见,具有同一克隆类型的不同植物在相似的环境中可形成不同的克隆构型,预示着它们具有不同的适应策略,同时说明,垫状锦鸡儿在克隆机理和适应性方面具有很大的独特性.
密集型克隆构型使垫状锦鸡儿子株可以近距离从母株获取所需的养分和水分等,对于生产力低下的荒漠区是一个节约的扩展方式;密集型克隆构型使灌丛内风速降低,有利于丛内水分保持,从而在内部形成一个相对湿度较高的微生境,提高了对干旱生境的适应能力;密集型克隆构型使垫状锦鸡儿母株能给分株更有效的保护(防风沙、防动物啃食和践踏等),增强了对干扰的抗性;密集型克隆构型植物还能通过沃岛效应为分株提供有效的营养供应,提高了对养分贫瘠生境的适应能力.由此看来,垫状锦鸡儿密集型克隆构型是对荒漠区环境的重要适应对策.
在根系调查中有时可看到连接垫状锦鸡儿分株的间隔子已部分枯死腐烂,而分株地上及地下部分依然正常生长的现象.这说明垫状锦鸡儿的分株在某种情况下可以与母株分离而独立生长繁殖.分株与母体分离可以降低互相连累的风险,这是垫状锦鸡儿通过分株风险分摊,适应恶劣环境的一种机制.
根据克隆器官的不同,克隆植物分成根茎型、匍匐茎型、块茎型、压条型、根出条(根蘖)型等.垫状锦鸡儿的基部枝条遭流沙掩埋后形成许多无性系分株,即垫状锦鸡儿的克隆器官是地上枝条,所以属于枝源型克隆植物(也叫压条型克隆植物).
在调查的5个集群中有3个是复合灌丛.其形成原因可能是掉落在灌丛内部的种子,由于母体灌丛(或其它灌丛)改变了内部的物理环境(风力、湿度、温度、土壤含水量和土壤肥力等),提高了种子萌发率;萌发的新个体受到原来灌丛的保护,避免了羊的踩踏.这样,存活下来的新个体与原来的灌丛组成复合大灌丛.在荒漠区,垫状锦鸡儿通过无性繁殖和有性繁殖两种方式来实现局域种群的延续和更新.尽管无性繁殖对克隆植物种群的扩增起首要作用,但有性繁殖通过个体间的异交维持了较高的遗传多样性,加快在环境选择过程中对环境的适应与物种形成;有性后代扩散的距离大于克隆后代,在拓殖新生境方面要优于克隆后代;此外有性过程产生的种子可通过休眠机制而使植物安全地度过不利环境时期.可见,保持一定比例的有性繁殖对垫状锦鸡儿适应荒漠环境有重要意义.
垫状锦鸡儿具有发达的根系,根深和根幅远大于株高和冠幅.其不定根尤为发达,入土深度多数在近地表至40 cm土层中,可长达80 cm.这使其扩大了在严酷生境中对水分和养分的吸收范围,同时增强了植株对风沙的抵抗力.主根深入地下,可利用较深层土壤蓄积的水分,这就导致垫状锦鸡儿不易受干旱的胁迫.这些特性充分体现了垫状锦鸡儿在进化过程中适应环境的能力,使其能够很好地在特定的环境条件下生长,从而确立其在群落中的地位.可见发达根系是垫状锦鸡儿适应荒漠环境的又一重要对策.
从以上的分析和讨论可以得出以下结论:1)在荒漠区,垫状锦鸡儿的营养繁殖在种群的更新和扩展中占主导地位,有性繁殖为辅;2)垫状锦鸡儿为枝源性克隆植物,其克隆生长构型为密集型;3)密集型的克隆构型和发达的根系,赋予垫状锦鸡儿很好的水土保持性能和抗旱性,为其种群繁盛创造了良好条件.
[1] 刘瑛心.锦鸡儿属[M]//中国植物志编辑委员会.中国植物志.第42卷第1册,北京:科学出版社,1993,13-67.
[2] 周道玮.锦鸡儿属植物分布研究[J].植物研究,1996,16(4):428-435.
[3] Xu X Y,Zhang R D,Xue X Z,et al.Determinaiton of evapotranspiration in the desert area using lysimeters[J].Commun Soil Sci Plant Anal,1998,29(1/2):1-13.
[4] 王生军,刘果厚.内蒙古灌木资源[M].呼和浩特:内蒙古大学出版社,2006.
[5] 马成仓,高玉葆,郭宏宇,等.内蒙古高原西部荒漠区锦鸡儿属(Caragana)优势种的形态适应特征[J].生态学报,2006,26(7):2308-2312.
[6] Ma C C,Gao Y B,Guo H Y,et al.Physiological adaptations of four dominantCaraganaspecies in the desert region of the Inner Mongolia Plateau[J].J Arid Environ,2008,72(3):247-254.
[7] 马成仓,高玉葆,李清芳,等.内蒙古高原荒漠区几种锦鸡儿属(Caragana)优势植物的生理生态适应特征[J].生态学报,2007,27(11):4643-4650.
[8] 陈玉福,董鸣.毛乌素沙地根茎灌木羊柴的基株特征和不同生境中的分株种群特征[J].植物生态学报,2000,24(1),40-45.
[9] 陈世璜,张昊,王立群,等.中国北方草地植物根系[M].长春:吉林大学出版社,2004.
[10] Price E A C,Marshall C.Clonal plants and environmental heterogeneity[J].Plant Ecol,1999,141:3-7.
[11] Schenk H J.Clonal splitting in desert shrubs[J].Plant Ecol,1999,141:41-52.
[12] Zhao W,Zhang Z,Li Q.Growth and reproduction ofSophoramoorcroftianaresponding to altitude and sand burial in the middle Tibet[J].Environ Geol,2007,53:11-17.
[13] 董清,付和平,斯琴高娃,等.内蒙古阿拉善左旗啮齿动物区系[J].内蒙古草业,2004,16(1):38-39.
[14] Bond W J,Midgley J J.Ecology of sprouting in woody plants:The persistence niche[J].Trends Ecol Evol,2001,16:45-51.
[15] 王洪义,王正文,李凌浩,等.不同生境中克隆植物的繁殖倾向[J].生态学杂志,2005,24(6):670-676.
[16] 赵晓英,任继周.三种锦鸡儿属植物的克隆生长特性及其植被恢复意义[J].中国水土保持科学,2005,3(4):102-107.
[17] Grime J P.Plant Strategies and Vegetation Processes.Chichester[M].New York:Great Britain at The Pitman Press,1979.
[18] 赵晓英,任继周,胡自治.三种锦鸡儿属植物的更新对策研究[D].兰州:甘肃农业大学,2004.
[19] 牛西午.柠条生态学研究[J].华北农学报,1998,13(4):122-129.
[20] 闫兴富.柠条的繁殖体及其在荒漠植被恢复中的作用[J].内蒙古林业科技,2007,32(2):23-27.
[21] Harper J L,Rosen B R,White J.The growth and form of modular organisms[J].Philosophical Transactions of the Royal Society,Series B,1986,313:3-5.
[22] Leakey R R B.Adaptive biology of vegetatively regenerating weeds[J].Adv Appl Bio,1981,6:57-90.
[23] 宋明华,董鸣.群落中克隆植物的重要性[J].生态学报,2002,22(11):1960-1963.
[24] 高润宏,金洪,张巍,等.阿拉善荒漠特有珍稀濒危植物绵刺克隆生长构型研究[J].干旱区资源与环境,2001,15(4):92-96.
[25] 袁秀英,袁登胜,白玉明.固定和半固定沙丘花棒克隆生长构型的比较研究[J].西北林学院学报,2006,21(6):32-34.
[26] 田福东,刘果厚.四合木不同种群间克隆构型及分株种群特征的比较研究[J].内蒙古农业大学学报,2007,28(4):98-101.
Propagation ofCaraganatibeticaand its ecological role in adaptation to arid desert environment
ZHANGJianhua1,MAChengcang1,LIQingfang1,Wuyunqiqige3,GAOYubao1,2
(1a.College of Life Science,1b.Tianjin Key Laboratory of Cyto-Genetical and Molecular Regulation,
Tianjin Normal University,Tianjin 300387,China;
2.College of Life Science,Nankai University,Tianjin 300071,China;
3.Xilinhot City Grassland Work Station,Xilinhot 026000,Inner Mongolia Autonomous Region,China)
In order to understand the reproductive characteristics ofCaraganatibeticain the desert areas and reveal its mechanism of adaptation to environment,the shrub structure,ramet characteristics,breeding tendency and clonal growth characteristics ofC.tibeticaclusters in the Alshan desert region of the Inner Mongolia Plateau were studied.The results showed that clonal propagation ofC.tibeticawas the dominant way in the arid desert region,andC.tibeticareproduced ramets mainly by branches covered by sand and had a clonal architecture of“phalanx”.C.tibeticawas well adapted to the arid environment of desert region by the higher proportion of asexual reproduction,phalanx clonal architecture and well-developed root systems,which made it be one of the dominant species in the desert region.
Caraganatibetica;asexual reproduction;arid desert region;adaptation
Q145
A
1671-1114(2011)04-0055-05
2011-05-27
国家自然科学基金资助项目(31170381);国家重点基础研究发展规划项目(2007CB106802)
张建华(1978—),女,硕士研究生.
马成仓(1963—),男,博士,教授,主要从事环境生物学和植物生理生态方面的研究.E-mail:machengcang@163.com
(责任编校 纪翠荣)
猜你喜欢
种子(2022年5期)2022-06-27
军民两用技术与产品(2022年1期)2022-06-01
中学生数理化(高中版.高考理化)(2021年12期)2021-03-08
防护林科技(2020年5期)2020-07-21
水土保持研究(2020年1期)2020-04-27
西藏农业科技(2019年2期)2019-07-10
西藏农业科技(2018年4期)2018-04-25
北京航空航天大学学报(2017年10期)2017-04-20
果树实用技术与信息(2016年8期)2016-09-21
航天返回与遥感(2014年4期)2014-07-31
