
福建省肠道病毒71型分离株(2010FJLY008)全基因组核苷酸序列分析*
2011-01-24 02:13周朝晖沈晓娜张拥军谢剑锋吴冰珊王金章翁育伟郑奎城严延生
中国人兽共患病学报 2011年1期
陈 炜,周朝晖,沈晓娜,2,张拥军,谢剑锋,吴冰珊,王金章,翁育伟,郑奎城,严延生
肠道病毒71型(Enterovirus 71,EV71)为小RNA病毒科肠道病毒属成员,是引起患者手足口病(Hand,foot and mouth disease,HFMD)的主要病原体。
自1997年开始,由 EV71病毒引起的手足口病暴发公共卫生事件在东亚及东南亚逐渐增多,并经常引起社会恐慌,例如1997年的马来西亚及1998年中国台湾的手足口病暴发[1-2]。2008年我国安徽省阜阳市等地区出现了主要由EV71病毒引起的手足口病暴发流行,同年5月份我国将手足口病纳入丙类法定传染病进行管理。而2010年手足口病疫情的防控形势更加严峻,截止9月份,监测结果显示,无论是福建省还是全国,手足口病发病率、重症病例发生率、死亡率,均明显高于前两年的同期水平。为了解肠道病毒71型在福建省的遗传背景,追踪EV71流行过程中可能发生的变异,为今后福建省及国内手足口病疫情防控提供理论基础,我们对从福建省一名手足口病患儿标本分离到的EV71型分离株(2010FJL Y008)进行了全基因组核苷酸序列测定及分析,结果报告如下。
1 材料与方法
1.1 材料
1.1.1 毒株 肠道病毒 71型分离株(2010FJL Y008)由本实验室通过接种2010年福建龙岩地区一位3岁手足口病男患儿咽拭子标本于RD细胞,分离两代收获,具体方法及结果判定见文献[3]。
1.1.2 试剂 AMV逆转录酶购于Promega公司;随机引物、TaqDNA聚合酶、RNA酶抑制剂购于Takara公司;病毒核酸提取试剂盒、胶回收试剂盒购于QIA GEN公司(德国);质粒提取试剂盒购于TIANGEN公司 (北京)。pMDTM18-T载体与DH5α感受态细胞均购于 Takara公司。
1.2 方法
1.2.1 病毒RNA提取 采用QIAamp viral RNA mini kit提取病毒RNA,按照试剂盒说明书进行操作。从140μL的第二代细胞培养物上清液获得50μL的总RNA溶液。
1.2.2 cDNA的制备 以提取的RNA为模板,按照AMV逆转录酶使用说明书操作,加入AMV逆转录酶和随机引物,37℃1h,合成病毒基因组的cDNA。
1.2.3 病毒基因组的克隆 以病毒基因组的cDNA为模板,在 TaqDNA聚合酶作用下,根据文献所用引物[4],分8个特异性片段扩增病毒基因组。PCR反应条件:94℃3min;94℃1min,50℃1min,72℃90sec,34个循环;72℃10min。PCR扩增的8个片段经1%琼脂糖凝胶电泳分离,用 QIAquick Gel Extraction Kit回收纯化后,分别克隆至pMDTM18-T载体。重组克隆经RV-M和M13-47通用引物鉴定。
1.2.4 DNA序列测定和分析 提取的阳性重组克隆质粒送 Takara公司测序。全部核苷酸和氨基酸序列分析和同源性比较应用DNASTAR软件完成,系统发生树构建应用Mega 4.0软件完成。所有参考序列均来自 GenBank,见表1。
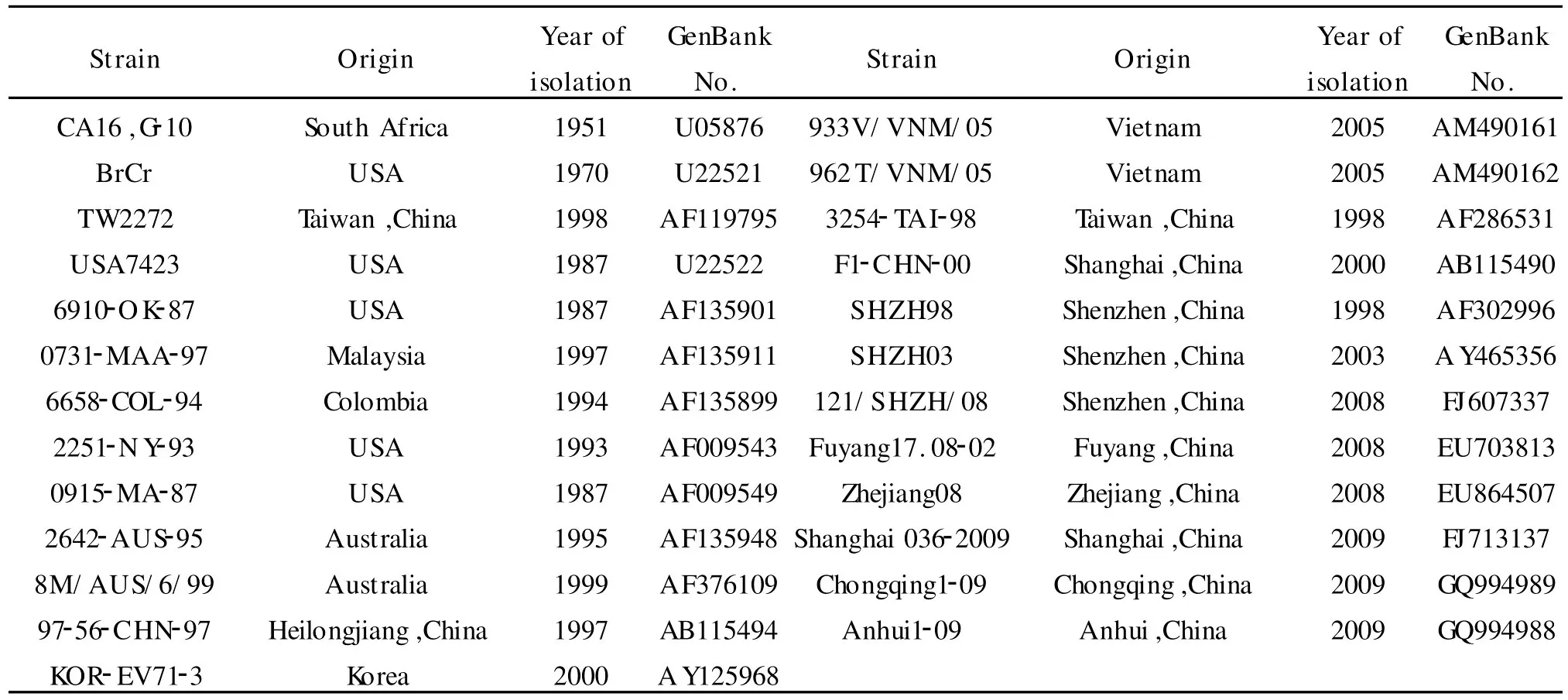
表1 序列分析中所用CoxA16和EV71毒株信息Table Background of CoxA16 and EV71 strains in sequence analysis
2 结 果
2.1 病毒全基因组片段的扩增、克隆与鉴定 从RD细胞培养上清液提取RNA,用随机引物做逆转录,得到覆盖肠道病毒71型分离株(2010FJL Y008)全长的cDNA片段,以此为模板,利用覆盖全基因组(除了多聚腺甘酸尾,即polyA尾)的8对引物进行PCR反应,1%琼脂糖凝胶电泳分离,得到7个1~1.4kb片段及一个近200bp的片段。8个片段经过回收纯化后分别克隆至pMDTM18-T载体。重组克隆经RV-M和M13-47通用引物鉴定,阳性质粒用于测序分析。
2.2 2010FJL Y008株全基因组核苷酸序列测定分析 经过序列测定、拼接、比对及分析后,获得肠道病毒71型分离株(2010FJL Y008)的全基因组核苷酸序列(未包括polyA尾),全长7 403bp,其中A占27.11%,G占23.94%,C占24.33%,U占24.63%,腺嘌呤核苷酸和尿嘧啶核苷酸较丰富(A+U=51.74%)。肠道病毒 71型分离株(2010FJL Y008)5′端的非编码区(5′U TR)长 740 个碱基,5′U TR之后是6582个碱基的病毒基因组开放读码框(ORF),编码一个2193个氨基酸的多聚蛋白,3′端的非编码区(3′U TR)长81个碱基。与其他EV71毒株相比,2010FJL Y008株在编码区没有核苷酸的缺失与插入,但在5′U TR与3′U TR区有核苷酸的缺失和插入存在。2010FJL Y008株全基因组的组成和结构符合 EV71型肠道病毒的特征,见表2。
2.3 基因组核苷酸序列同源性比较 将2010FJL Y008株与来自 GenBank的其他 CA16、EV71型毒株进行核苷酸序列同源性比较。从表3可以看出,无论是全长基因组,还是在5′U TR、P1、P2、P3、3′U TR各个分区中 ,2010FJL Y008 株与2008年安徽阜阳流行株 Fuyang17.08-2的同源性均最高;在全长基因组及 5′U TR、P1区中,2010FJL Y008株与CA16,G-10的同源性最低;在P2区,2010FJL Y008株与BrCr株同源性最低;在P3、3′U TR 区 ,2010FJL Y008株与 TW2272同源性最低。
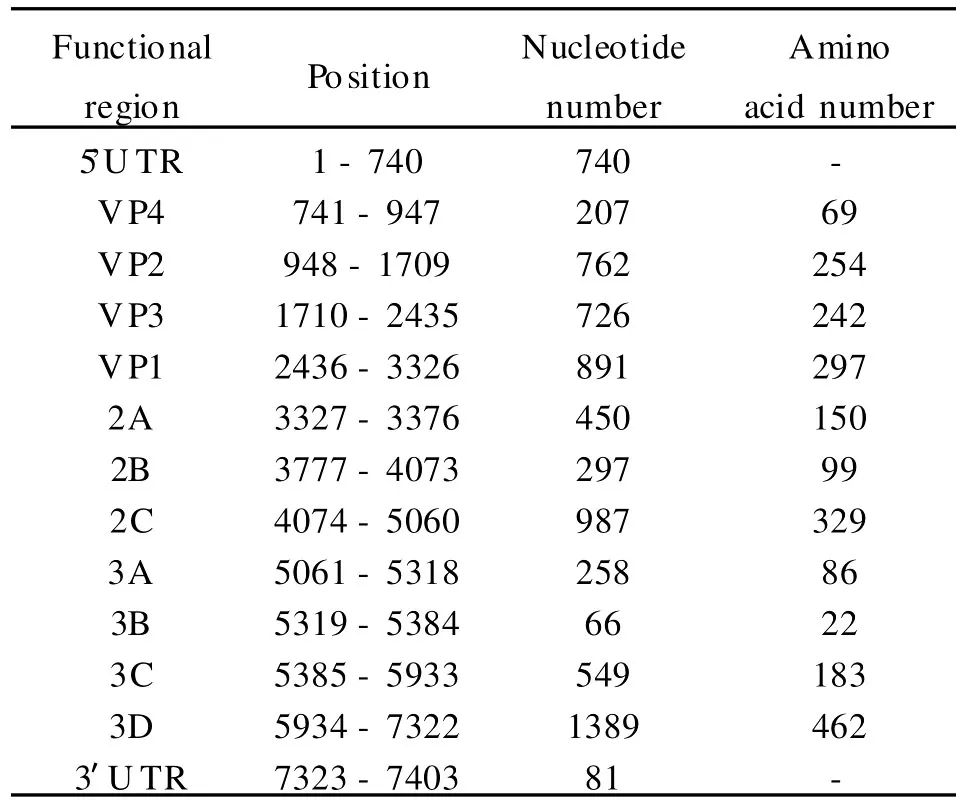
表2 2010FJLY008株基因组核苷酸及编码区氨基酸数目Table 2 Nucleotide andamino acid numbers in coding regions of 2010FJLY008
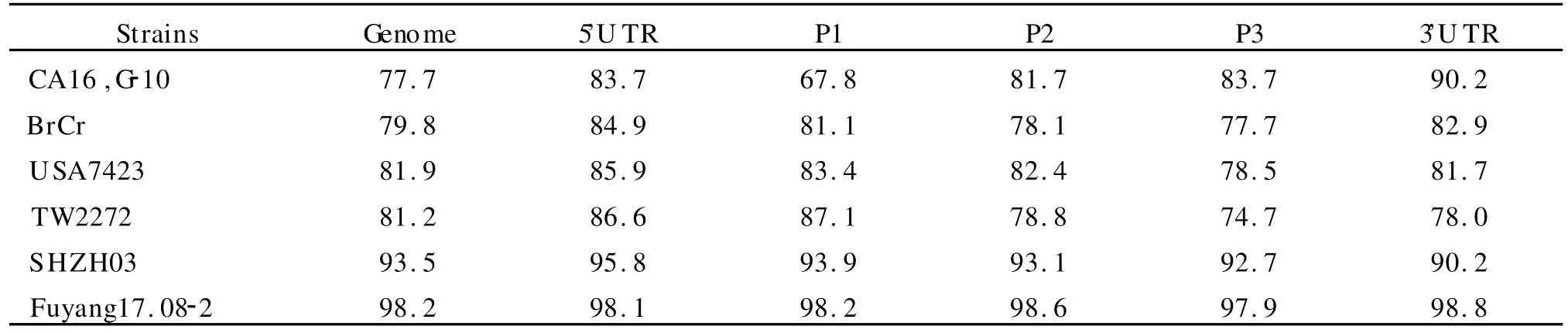
表3 2010FJLY008株与其他CA16、EV71型毒株核苷酸序列同源性比较(%)Table 3 Comparison of nucleotide sequence homology among 2010FJLY008 and other CA16 and EV71 strains(%)
2.4 编码蛋白氨基酸序列同源性比较2010FJL Y008株病毒基因组编码区全长6582个核苷酸,共编码2 193个氨基酸。将2010FJL Y008株与其他CA16、EV71型毒株氨基酸序列进行同源性比较,见表4。从编码的氨基酸组成成分分析,2010FJL Y008株与 Fuyang17.08-2株在 P1、P2、P3的同源性均最高;在 P1区,2010FJL Y008株与CA16,G-10株同源性最低;在 P2、P3区,2010FJL Y008株与 TW2272株同源性最低。从编码的结构蛋白分析,在 VP1、VP2及 VP4区,2010FJL Y008株与 Fuyang17.08-2株的同源性最高,其中在 VP2、VP4区同源性为 100%;在 VP3区,2010FJL Y008株与 SHZH03的同源性最高;在VP1、VP2、VP3及 VP4区,2010FJL Y008株与CA16,G-10的同源性均最低。
2.5 病毒基因的遗传进化分析 使用Mega 4.0软件,通过邻接法(Neighbor-joining),基于全长VP1基因,以 CA16,G-10为外群,对2010FJL Y008株与其它24株已知基因型的 EV71毒株进行遗传进化分析(图 1)。可以看出,2010FJL Y008株与Fuyang17.08-2株进化关系最近,同属于C4基因亚型的C4a进化分支;同时,以CA16,G-10为外群,对2010FJL Y008株与其它来自不同地区和年代的共11株EV71毒株的全长基因组核苷酸序列进行遗传进化分析(图2)。同样可以看出,2010FJL Y008株与 Fuyang17.08-2株的亲缘关系最密切,而2008、2009年以来国内不同省市分离到的 EV71毒株无论在VP1还是在全基因组核苷酸水平均存在密切联系,而与国外一些EV71毒株存在较大差异。
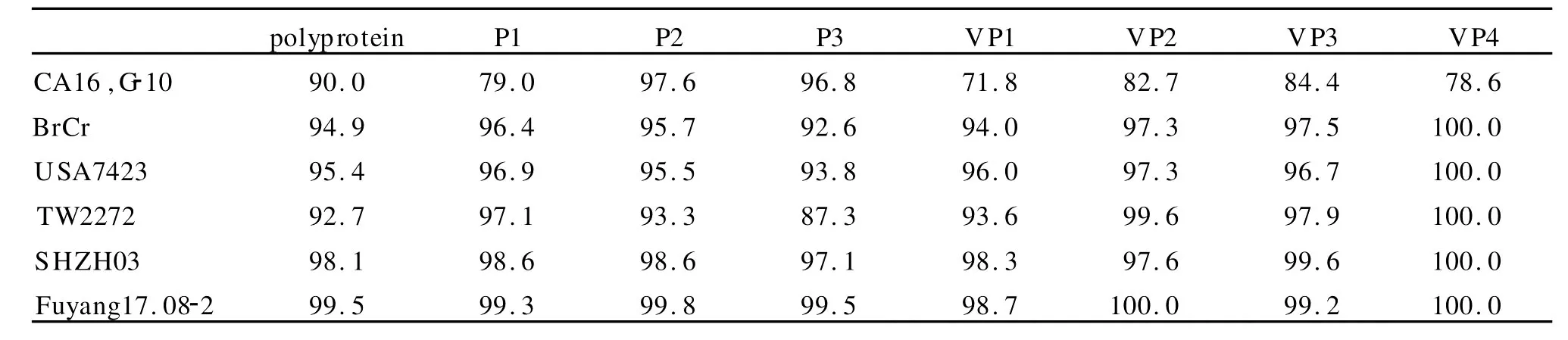
表4 2010FJLY008与其他CA16、EV71型毒株氨基酸序列同源性比较(%)Table 4 Comparison of amino acid sequence homology among 2010FJLY008 and other CA16 and EV71 strains of(%)

图1 lEV71病毒VP1区的遗传进化分析Fig.1 Phylogenetic tree of VP1 regions in EV71 viruses
3 讨 论
2010FJL Y008株是2010年在福建省龙岩地区手足口病患儿(普通病例)的咽拭子标本中分离的毒株,经过Real-time RT-PCR鉴定为 EV71型毒株。病毒全基因组序列分析表明,2010FJL Y008株与其它EV71型毒株一样,具有相同的基因组结构。2010FJL Y008株全长7403bp(未包括polyA尾),仅有一个开放编码框(ORF),位于全基因组的第741-7322位核苷酸,编码2193个氨基酸的多聚蛋白,该多聚蛋白可进一步被水解成 P1、P2、P3三个前体蛋白,P1前体蛋白编码 VP4、VP2、VP3、VP1四个病毒外壳蛋白,P2、P3前体蛋白主要编码蛋白水解酶及RNA聚合酶,在编码区的两端分别有740个核苷酸的5′U TR及81个核苷酸的3′U TR。
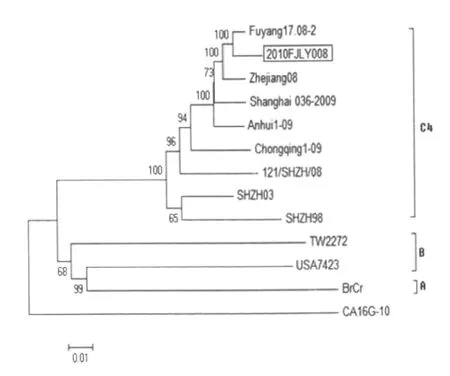
图2 EV71病毒全长基因组核苷酸序列的遗传进化分析Fig.2 Phylogenetic tree of genomic nucleotide sequences in EV71 viruses
基因组核苷酸序列同源性比较及编码氨基酸序列同源性比较显示:2010FJL Y008株与EV71标准株BrCr相比,核苷酸变异为20.2%,氨基酸变异为5.1%;而与2008年阜阳流行株(Fuyang17.08-2)相比,核苷酸及氨基酸变异分别只有1.8%、0.5%;2010FJL Y008株在 5′U TR与 3′U TR区有核苷酸的缺失和插入存在,在编码区没有核苷酸的缺失和插入。
虽然目前还没有确切证据表明EV71的毒力或感染结局与基因组的任何特异位点有关,大部分研究都集中在 EV71病毒的VP1区。Obereste等在HEV71病毒的VP1区分子进化研究及其应用于分类小RNA病毒中指出:VP1区编码蛋白含有大量的病毒中和因子,全长VP1区核苷酸序列分析,可作为肠道病毒属内不同血清型分类的依据,也可以作为小RNA病毒科内不同属的分类参考[5]。基于全长VP1区核苷酸序列进行遗传进化分析被公认为是进行 EV71基因型鉴定方法。根据Brown等[6]关于EV71病毒分子流行病学及进化研究,他对1970年至1998年在美国及其他5个国家分离的113株 EV71型毒株的全长VP1区基因进行了分析,把所有的 EV71分为A、B、C三个基因型。本实验通过对全长 VP1区基因的遗传进化分析,2010FJL Y008株与 Fuyang17.08-2株的亲缘关系密切,包括 SHZH03在内同属于 C4基因亚型的C4a进化分支。杨秀惠等[7]在2008年对我省福州、泉州及龙岩地区的8株 EV71进行全长VP1区基因种系发生分析,2008年福建与阜阳分离株和中国大陆1998年以来的病毒株均为 C4亚型。因此,与2008年相比,2010年福建省的流行株(2010FJL Y008)的基因型并未发生变异,未产生明显抗原漂移及变异,仍然与2008年阜阳分离株一样,为C4基因亚型。同时,全长基因组核苷酸序列的遗传进化分析,2010年福建省的流行株(2010FJL Y008)与2008年阜阳流行株(Fuyang17.08-2)的亲缘关系同样接近。另外,通过全长基因组核苷酸序列的遗传进化分析,我们发现,EV71株与CA16株依然各自形成一个分支。而在EV71株的分支中,基于VP1区核苷酸序列遗传进化分析认为是B基因型的毒株(USA7423)与A基因型毒株(BrCr)存在较近的进化关系。
总之,2010年福建省 EV71型病毒流行株(2010FJL Y008)在病毒全基因组结构、基因组核苷酸同源性比较、氨基酸同源性比较、全长基因组核苷酸序列遗传进化分析及VP1区基因遗传进化分析等方面,均与2008年阜阳流行株(Fuyang17.08-2)关系密切。但是,随着对手足口病诊疗水平及重视程度的提高,2010年福建省乃至中国大陆,由EV71型病毒引起的手足口病发病率、重症率、死亡率均明显高于2008、2009年,尽管全基因组进化分析显示近几年国内不同地区的 EV71分离株联系紧密,是否存在病毒基因组未知位点的变异导致病毒致病能力的变化,依然缺乏证据。因此,今后手足口病的监测工作不能有丝毫松懈,需要进一步开展病毒基因片段核酸序列分析,追踪病毒在流行过程中发生的任何遗传变异,为 EV71疫苗研制和有效控制手足口病奠定基础。
[1]Shih S R,Ho M S,Lin K H,et al.Genetic analysis of enterovirus 71isolated from fatal and non-fatal cases of hand,foot and mouth disease during an epidemic in Taiwan,1998[J].Virus Research,2000,68(2):127-136.
[2]AbuBakar S,Chee H Y,Al-Kobaisi M F,et al.Identification of enterovirus 71 isolates from an outbreak of hand,foot and mouth disease(HFMD)with fatal cases of encephalomyelitis in Malaysia[J].Virus Research,1999,61(1):1-9.
[3]国家脊髓灰质炎和国家麻疹实验室.手足口病实验室手册[R].4版.北京:中国疾病预防控制中心,2010:20-26.
[4]周世力,李琳琳,何雅青,等.我国分离的肠道病毒 71型(SHZH03)病毒株全基因组核苷酸序列分析[J].病毒学报,2004,20(1):7-11.
[5]Obereste M S,Maher K,Kilpatrick D R,et al.Molecular evolution of the human enteroviruses:correlation of serotype with VP1 sequence and application to picornavirus classi cation[J].J Virol,1999,73(3):1941-1948.
[6]Brown BA,Oberste M S,Alexander J P,et al.Molecular epidemiology and evolution of enterovirus 71 strains isolated from 1970 to 1998[J].J Virol,1999,73(12):9969-9975.
[7]杨秀惠,严延生,周勇,等.2008年福建省肠道病毒71型分离株的遗传特征分析[J].海峡预防医学杂志,2009,15(6):4-7.
猜你喜欢
今日畜牧兽医(2022年1期)2023-01-06
世界科学技术-中医药现代化(2022年3期)2022-08-22
肝博士(2022年3期)2022-06-30
科学大观园(2022年2期)2022-01-23
中国饲料(2019年19期)2019-03-25
中国猪业(2017年11期)2017-12-11
中国医药指南(2017年3期)2017-11-13
右江医学(2014年1期)2014-03-22
郑州大学学报(理学版)(2014年3期)2014-03-01
