
尖孢镰刀菌亚麻专化型生物学特性研究
2011-04-10 02:16关凤芝吴广文宋喜霞姜卫东王立群
东北农业大学学报 2011年7期
潘 虹,关凤芝,吴广文,宋喜霞,姜卫东,王立群*
(1.东北农业大学生命科学学院,哈尔滨 150030;2.黑龙江省农业科学院经济作物研究所,哈尔滨 150086)
亚麻是我国重要的经济作物之一,其种植区域主要集中在黑龙江和吉林两省,全国年种植面积15万hm2左右[1-2]。亚麻枯萎病是由尖孢镰刀菌亚麻专化型 [Fusarium oxysporum Schl.f.sp.lini(Bolley)Snyder&Hansen]病原体引起的一种世界性病害,各国的亚麻种植区均有本病报道,发病率一般在5%~10%左右,某些产区高达20%以上,甚至绝产[3-6]。亚麻枯萎病苗期发生的主要表现为叶片枯黄,成片萎蔫、状似火烧,幼根缢缩,最终全株萎凋倒伏而死[7-8],因此,该病害严重影响亚麻原茎产量和纤维质量。张志铭、王小静等对尖孢镰刀菌亚麻专化型病原体进行了鉴定及形态描述[9];杨学等对病害的发生症状、传播途径及其发生流行规律等进行了调查并提出了可行性的防治措施[10]。可见,国内外对该病原体的研究主要集中在病原鉴定、致病性测定及病害防治等方面,有关生物学特性方面的研究,目前未见系统的研究报道[11]。为此,本文从菌丝生长、分生孢子产生和萌发三方面对该病原体的生物学特性进行了全面系统的研究,以明确不同营养及环境条件对该病原体菌丝生长量、产孢量及孢子萌发率的影响,为研究亚麻枯萎病的发生、流行规律以及病害防治等提供相关依据。
1 材料与方法
1.1 病菌来源
黑龙江麻区枯萎病株。
1.2 病菌分离与鉴定
采用常规的组织分离法进行病菌分离。首先,切取亚麻植株病患组织小块,经无菌水冲洗、酒精消毒、再冲洗后接入PDA平板,25℃恒温培养5 d;其次,选取形态典型的培养菌丝对其进行纯化、镜检、鉴定;再次,将病菌接种于麦粒培养基上产孢培养,在亚麻幼株上进行回接试验,10 d后幼株发病,症状典型;最后,在患病幼株上重新分离到该病菌菌株,确定其为尖孢镰刀菌亚麻专化型病原体。
1.3 温度对菌丝生长、产孢量及孢子萌发的影响
菌龄7 d、直径为0.5 cm的菌碟接种于PDA平板中央,分别置于5、10、15、20、25、30、35和40℃的恒温培养箱中培养,5 d后采用“十”字法测量菌落直径[14],每处理3次重复。
上述温度培养10 d的菌落整体移入100 mL无菌水中、振匀、制成孢子悬液,血球计数板计数,测定各温度下的产孢量,每处理3次重复。
菌龄10 d的菌落制成浓度约107个·mL-1的孢子悬液,以载玻片培养法[12-13]于上述温度保湿、暗培养24 h,检测孢子萌发率,每处理3次重复。
1.4 pH对菌丝生长、产孢量及孢子萌发的影响
配制 pH 为 2、3、4、5、6、7、8、9、10 和11的PDA培养基[14],分别制平板,接种同龄菌碟,25℃培养5 d,测量菌落直径。在上述pH条件下,测定产孢量及孢子萌发率,方法同1.3。
1.5 光照对菌丝生长、产孢量及孢子萌发的影响
用PDA平板接种同龄菌碟,分别置于持续光照(日光灯,MT-3095)、持续黑暗和12 h光暗交替的25℃培养箱中培养5 d,测量菌落直径。在上述光照条件下,测定产孢量及孢子萌发率,方法同1.3。
1.6 培养基对菌丝生长、产孢量及孢子萌发的影响
将马铃薯葡萄糖培养基(Potato dextrose agar,PDA)、马铃薯蔗糖培养基(Potato sucrose agar,PSA)、马铃薯麦芽糖培养基(Potato maltose agar,PMA)、查氏培养基(Czapek)、马丁氏培养基(Martin)和沙氏培养基(Sabouraud′s)分别制平板后,接种同龄菌碟,25℃培养5 d,测量菌落直径。在上述各培养基条件下,测定产孢量及孢子萌发率,方法同1.3。
1.7 碳源对菌丝生长、产孢量及孢子萌发的影响
以 Crapek(NaNO32 g、K2HPO41 g、KCl 0.5 g、MgSO40.5 g、FeSO40.01 g、蔗糖 30 g、琼脂 15~20 g、水1 000 mL、pH自然)为基础培养基,再分别以等碳量葡萄糖、麦芽糖、D-木糖、淀粉、甘露醇、果糖、山梨醇、乳糖替代蔗糖作为供试碳源,配成含不同碳源的培养基[15],制成平板,接种同龄菌碟,并设置无碳对照(CK1),25℃培养5 d,测量菌落直径。在上述各碳源下,测定产孢量及孢子萌发率,方法同1.3。
1.8 氮源对菌丝生长、产孢量及孢子萌发的影响
以Czapek为基础培养基,再分别以等氮量的尿素、牛肉膏、蛋白胨、L-天冬酰胺(L-Asn)、L-半胱氨酸(L-Cys)、DL-苯丙氨酸(DL-Phe)、L-组氨酸(L-His)、L-赖氨酸(L-Lys)替代硝酸钠作为供试氮源,配成含不同氮源的培养基,制成平板,接种同龄菌碟,并设置无氮对照(CK2),25℃培养5 d,测量菌落直径。在上述各氮源下,测定产孢量及孢子萌发率,方法同1.3。
1.9 数据分析
菌落直径测量:菌落直径(cm)=菌落测量直径(cm)-菌碟直径(cm);产孢量测定:选用规格为25×16格的血球计数板,设五个中方格中总孢子数A,孢子悬浮液稀释倍数B,样品稀释度要求显微镜下每方格内约有25~50个孢子为宜,每菌落产孢量(个)=A/5×25×B×1 000=5 000A×B(个);孢子萌发率检查:显微镜下每个视野中检查50~70个孢子,以芽管长度大于孢子一半时计为萌发,萌发率(%)=萌发孢子数/检查孢子总数×100%。使用DPS 7.05软件进行数据统计分析[16]。
2 结果与分析
2.1 温度对菌丝生长、产孢量及孢子萌发的影响
由图1可知,所试各温度下尖孢镰刀菌亚麻专化型病原体的菌丝均可生长,但不同温度间差别较大。其中30℃下菌丝生长最快,与其他处理相比,统计结果差异显著,低于5℃和高于40℃时无生长。
从图2中可以看出,该菌在10~35℃范围内均能产孢及孢子萌发。25℃时产孢量最大,且显著高于其他各处理。25和30℃下孢子萌发率高达60%以上,且二者与其他处理相比差异显著。在5、40℃时产孢量及萌发率均为零。
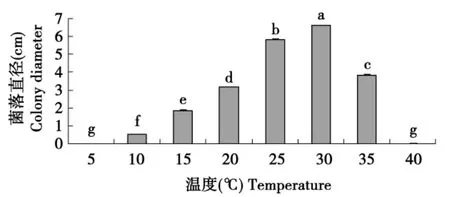
图1 温度对菌丝生长的影响Fig.1 Influence of temperature on mycelium growth of F.oxysporum Schl.f.sp.lini
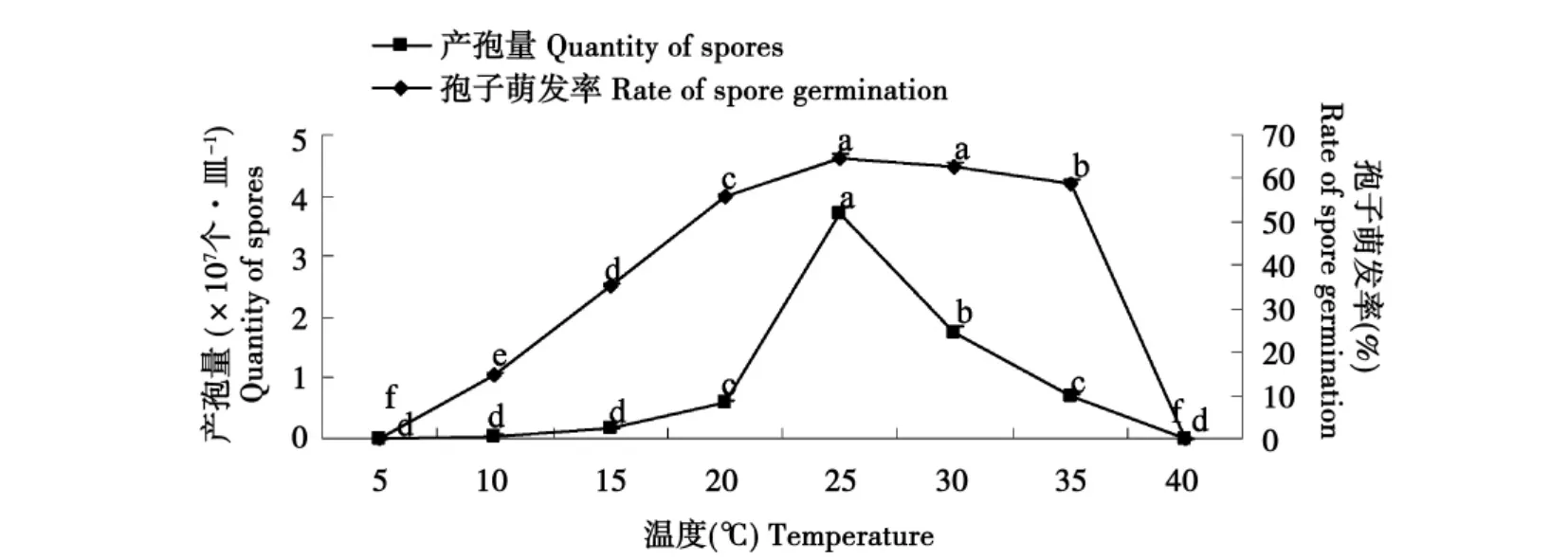
图2 温度对产孢量和孢子萌发的影响Fig.2 Influence of temperature on quantity of spores and spore germination of F.oxysporum Schl.f.sp.lini
2.2 pH对菌丝生长、产孢量及孢子萌发的影响
结果见图3。
尖孢镰刀菌亚麻专化型病原体在pH 2~11范围内均可生长,在pH 7、8时菌丝生长量与其他处理比差异显著,此外相应递减,但酸性条件比碱性下生长量减少幅度大。
可见该菌与其他多数喜好酸性环境的真菌不同,即适应碱性的能力较酸性强。

图3 pH对菌丝生长的影响Fig.3 Influence of pH on mycelium growth F.oxysporum Schl.f.sp.lini
从图4可以看出,产孢量在pH 7、孢子萌发率在pH 8时最高,其他处理均较低,但偏碱性环境的产孢量会略大于偏酸性,说明中性或略偏碱性环境更利于产孢。
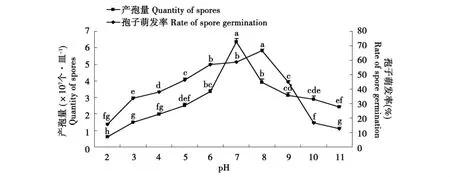
图4 pH对产孢量和孢子萌发的影响Fig.4 Influence of pH on quantity of spores and spore germination of F.oxysporum Schl.f.sp.lini
2.3 光照对菌丝生长、产孢量及孢子萌发的影响
由表1可知,各光照处理下菌丝生长状况良好,气生菌丝繁茂,彼此间生长量均表现为差异极显著,具体为:持续黑暗条件下的菌丝生长最快,光暗交替次之,连续光照最差。说明黑暗有利于菌丝生长,光照有抑制作用,而这种光照的抑制作用除了影响真菌的代谢机制,更重要表现在其对培养基成分的破坏[17]。
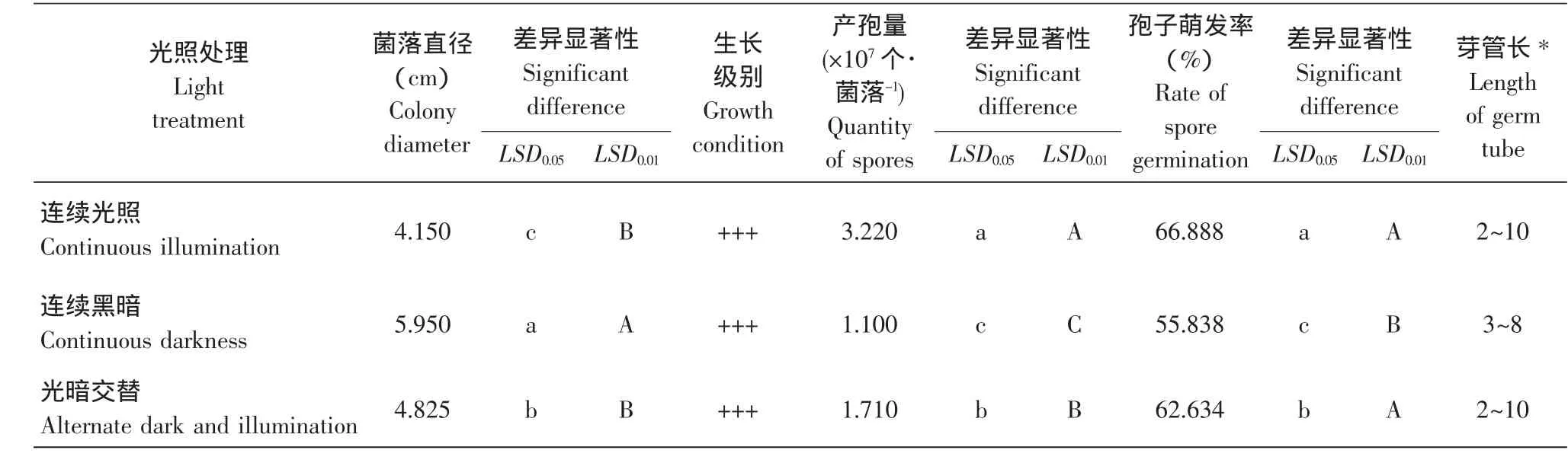
表1 不同光照处理对尖孢镰刀菌亚麻专化型菌丝生长、产孢量和孢子萌发的影响Table 1 Influence of light treatment on mycelium growth,quantity of spores and spore germination of F.oxysporum Schl.f.sp.lini
从产孢量和孢子萌发率来看,持续光照下二者数值最高,其次是光暗交替,持续黑暗最低。可见,当该病原体进入繁殖阶段时,分生孢子的产生和萌发都具喜光性,这一点与Cochrane等的假说,即光触发真菌产孢机制之论断一致[18-19]。
2.4 培养基对菌丝生长、产孢量及孢子萌发的影响
由表2可知,所试6种培养基中的PDA、PSA、Sabouraud′s及Crapek有利于菌丝生长, 菌落厚而致密、呈絮状,菌落直径均可达到6.0 cm以上;PMA、Martin上生长的菌落直径明显小于以上,其中Martin上的菌丝稀疏,仅有少量生长。
从产孢量和孢子萌发率来看,分生孢子在PSA上产量最大,每皿达到6.900×107个;PMA、Martin上产量最低,不足107个。孢子萌发率是PDA上最高,达到65%以上;以后排列次序为PSA、Sabouraud's、Czapek、PMA和Martin,并且Martin的孢子萌发率与其他处理相比差异极显著。由此可见,Martin对分生孢子的产生及萌发均有不利影响。

表2 不同培养基对尖孢镰刀菌亚麻专化型菌丝生长、产孢量和孢子萌发的影响Table 2 Influence of medium on mycelium growth,quantity of spores and spore germination of F.oxysporum Schl.f.sp.lini
2.5 碳源对菌丝生长、产孢量及孢子萌发的影响
由表3可知,菌丝在所试的10种碳源中,以淀粉、蔗糖、麦芽糖为碳源的培养基上生长较快,但三者在统计学0.01水平上无明显差异;在以甘露醇、山梨醇、葡萄糖、果糖及乳糖为碳源的培养基上生长相对缓慢;而在以极少被真菌利用的D-木糖上,菌丝生长量下降明显;甚至低于无碳对照,说明D-木糖对其生长非但不是碳源,而且还有负面的抑制作用。
试验表明不同碳源对产孢量和萌发率的影响较大。该菌在以乳糖作碳源时产孢量最大,是其他各处理的3~13倍,统计结果差异极显著。以果糖作碳源时孢子萌发率最高,达60%以上;山梨醇、甘露醇为碳源是孢子萌发效果不及无碳对照,因此可见,二者对孢子的萌发而言同样不可作为碳源,并表现为抑制作用。

表3 不同碳源对尖孢镰刀菌亚麻专化型菌丝生长、产孢量和孢子萌发的影响Table 3 Influence of carbon sources on mycelium growth,quantity of spores and spore germination of F.oxysporum Schl.f.sp.lini
2.6 氮源对菌丝生长、产孢量及孢子萌发的影响
由表4可知,所试10种氮源中,尖孢镰刀菌亚麻专化型病原体对氯化铵的利用最好;之后排序为:尿素、蛋白胨、L-Asn,硝酸钠、L-His、牛肉膏、L-Cys、无氮处理和亚硝酸钠。其中的亚硝酸钠在P=0.01的水平上与前9种氮源相比,均差异极显著,表现为其生长量极少,甚至低于无氮对照,说明此种氮源在该菌的生长过程中产生抑制作用。
对于产孢量而言,氯化铵、L-Asn为氮源时孢子产量均可高达每个菌落2.8×107个以上。在对该菌菌丝生长及产孢均不好用的L-Cys处理下的孢子萌发最好,且显著高于其他处理。
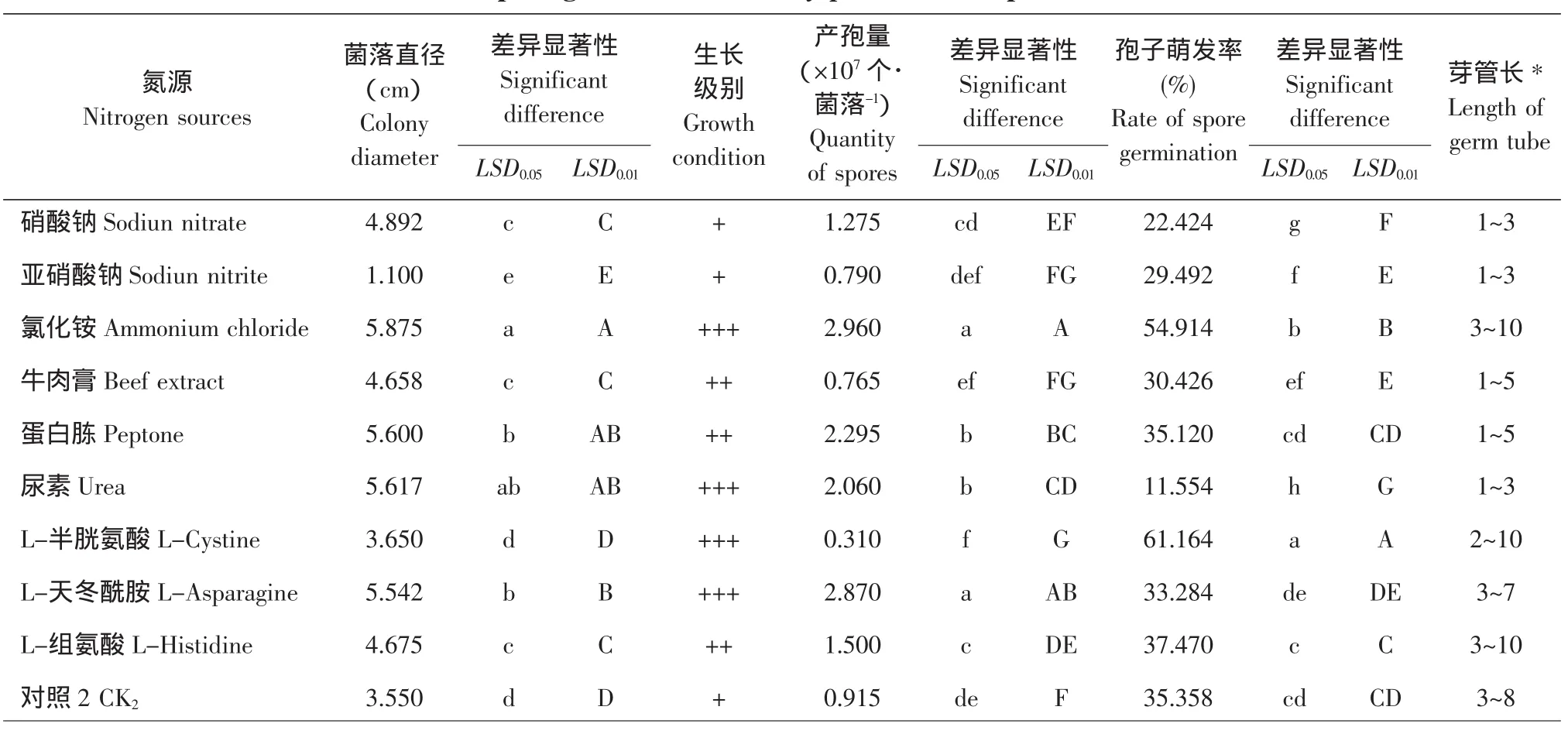
表4 不同氮源对尖孢镰刀菌亚麻专化型菌丝生长、产孢量和孢子萌发的影响Table 4 Influence of nitrogen sources on mycelium growth,quantity of spores and spore germination of F.oxysporum Schl.f.sp.lini
3 讨论与结论
试验结果表明,尖孢镰刀菌亚麻专化型病原体菌丝生长和孢子萌发的温度范围为10~35℃,最适温度分别是30和25℃,可见,前者对温度的需求略高,即是说,在适宜温度下,尽管菌丝生长状况良好,但并非有利于孢子萌发,实践中可通过维持高温生长条件来控制病原菌繁殖。野外调查发现,黑龙江省麻区土温在20℃时亚麻植株开始发病;25~30℃时最适合枯萎病的发生,是发病的高峰期;超过35℃时,病情停止发展[7]。可见,尖孢镰刀菌亚麻专化型病原体的这一温度特性与患病植株每年田间的消长规律密切相关,这也是病原菌与寄主之间在自然环境条件下长期适应和选择的结果。
碳源不仅为真菌细胞的关键组分提供了所需要的碳素,而且其氧化过程也为真菌基本生命活动提供了能源。试验证明,该病原体菌丝生长喜好淀粉等难分解的碳源,而较少利用多数真菌易于吸收的葡萄糖、果糖,究其原因,可能是病原体在生长代谢过程中受到环境因素诱导而分泌胞外淀粉酶,进而分解供试营养以便于机体吸收。另外,多数真菌不能利用CO2作为主要碳源,但少数的种已经被证实,如镰刀菌属的一些种,在特定条件下具有固定CO2的能力[18-20],这可能是本试验无碳对照处理下菌丝有少量生长的原因。
氮源对真菌的生长和发育亦是不可缺少的,他的作用主要是合成多种关键的细胞组分,如氨基酸、核苷酸、嘌呤、嘧啶等。尖孢镰刀菌亚麻专化型病原体在生长发育的不同阶段对所需氮源分子结构需求不同,其利用形式分析如下:铵作为参与代谢过程中的直接底物,其重要作用是影响真菌中一系列发育系统的调节,如形成可育的子囊壳,故多数真菌通常优先利用铵态氮作为主要的氮素营养[18],本菌亦如此;另外,大部分真菌喜好的有机氮也是该病原体较好利用的氮源,机体内有机氮被吸收后首先转换为铵态氮,而后进一步合成所需物质参与细胞代谢,如对尿素、L-Asn的利用;该菌对硝酸盐和亚硝酸盐的利用较弱,可能是由于自身不能合成硝酸盐还原酶的缘故,而亚硝酸盐对氨基酸的脱氨基作用和干扰硫代谢的能力,使得其对多数真菌是有毒的,仅被极少数真菌作为氮源利用,对本菌的毒性作用尚待研究。因此,该病原体对氮素的有效利用顺序可归纳为,铵态氮>有机氮>硝态氮。
尽管该病原体为亚麻专化型寄生菌,但其对碳、氮源的利用范围较广,这一点与多种绝对寄生菌对营养物质的高要求略有差异。在该菌喜好的营养中,氮源中的有机氮及碳源中的单、双糖两类物质在亚麻植株组织中的含量较高,这为病原体的快速侵入、生长及繁殖提供了一个适宜的营养环境[21]。
尖孢镰刀菌种内的致病株具有高度的寄主专化性,其中因侵染黄瓜、西瓜、番茄等作物而导致世界范围内的毁灭性病害。生物学特性方面,黄瓜、西瓜、番茄各专化型病原体在5~35℃温度范围内均可生长、产孢及萌发,其中以25~30℃为最适温度,这一点与尖孢镰刀菌亚麻专化型病原体一致;尖孢镰刀菌种内各致病株对碳、氮源的选择性差异较小,碳源以对蔗糖、麦芽糖、葡萄糖及乳糖的利用较好,氮源以对蛋白胨、牛肉膏、氯化铵、脲素的利用较好[22-25]。
本文对尖孢镰刀菌亚麻专化型病原体的生物学特性进行了相关研究,将为进一步开展亚麻枯萎病发生、流行规律、生物防治技术以及高效低毒药剂筛选等研究提供必要的基础理论依据。
[1] 杨学.亚麻苗期病害发生特点及防治技术研究[J].中国麻业,2003,25(5):223-227.
[2] 中国农作物病虫害编辑委员会.中国农作物病虫害(上册)[M].北京:农业出版社,1979:768.
[3] 魏景超.真菌鉴定手册[M].上海:上海科学技术出版社,1979:618.
[4] 布斯C著.镰刀菌属[M].陈其焕,译.北京:农业出版社,1988:24-27,187-188.
[5] Spielmeyer W,Lagudah E S,Mendham N,et al.Inheritance of resistance to flax wilt(Fusarium oxysporum f.sp.lini Schlecht)in a doubledhaploidpopulationof Linumusitatissimum L.[J].Euphytica,1998(3):287-291.
[6] Kroes G M L W,Sommers E,Lange W.Two in vitro assays to evaluate resistance in Linum usitatissimum to Fusarium wilt disease[J].EuropeanJournalofPlantPathology,1998,104(7):561-568.
[7] 杨学,王玉富,关凤芝,等.亚麻枯萎病发生规律及其综合防治措施[J].中国麻业,2002,24(1):23-26.
[8] Carl A B,Scott H,Robert A H.Evaluation of fungicide seed treatmentsonflaxcultivarsdifferinginseedcolor[J].IndustrialCrops and Products,2007,25:301-304.
[9] 王小静,李敏权.甘肃中部地区亚麻枯萎病病原菌及其致病性差异研究[J].中国麻业科学,2007,29(4):207-211.
[10] 杨学.亚麻病害症状及检索表[J].中国麻业,2002,24(5):23-27.
[11] Hano C,Addi M,Fliniaux O,et al.Molecular characterization of cell death induced by a compatible interaction between Fusarium oxysporum f.sp.linii and flax(Linum usitatissimum)cells[J].Plant Physiology and Biochemistry,2008,46(5-6):590-600.
[12] 方中达.植病研究方法[M].3版.北京:中国农业出社,1998:141,151-154.
[13] 俞大绂.植物病理学和真菌学技术汇编[M].北京:人民教育出版社,1977:410-411.
[14] Collins C H,Patricia M L.Microbiological methods[M].Fourth Edition.London:Butterworth,1976:110-113.
[15] Jennings D H.The physiology of fungal nutrition[M].Cambridge:Cambridge University Press,2007:87-109.
[16] 唐启义,冯明光.DPS数据处理系统:实验设计、统计分析及数据挖掘[M].北京:科学出版社,2002:85-98.
[17] Carlile M J,Watkinson S C.The fungi[M].Academic Press,1994:139-140.
[18] 邢来君,李明春.普通真菌学[M].北京:高等教育出版社,1999:51,58,62-63,100-101.
[19] 阿历索保罗C J,明斯C W著.真菌学概论[M].余永年,宋大康,译.北京:农业出版社,1983:14.
[20] 柏内特J H著.真菌学基础[M].刘锡琎,白金铠,译.北京:科学出版社,1989:170.
[21] 黄思良,廖明,岑贞陆,等.八角炭疽病菌生物学特性的初步研究[J].中国森林病虫,2006,5(1):1-4.
[22] 陈霞,刘东,张艳菊,等.黄瓜枯萎病病株镰孢菌的分离与鉴定[J].东北农业大学学报,2010,41(7):37-44.
[23] 陈秀蓉,魏勇良.甘肃省西瓜枯萎病病原菌鉴定及生物学特性测定[J].甘肃农业大学学报,1991(3):258-294.
[24] 肖荣凤,刘波,朱育菁,等.西瓜枯萎病原菌鉴定及其生物学特性的研究[J].武夷科学,2004,20:70-73.
[25] 徐艳辉,李烨,许向阳.番茄枯萎病的研究进展[J].东北农业大学学报,2008,39(11):128-134.
猜你喜欢
草原与草坪(2022年5期)2023-01-29
吉林农业科技学院学报(2022年1期)2022-05-14
麦类作物学报(2019年6期)2019-07-24
西南农业学报(2018年11期)2018-12-11
中国麻业科学(2018年6期)2018-04-09
纤维复合材料(2018年4期)2018-02-18
西南农业学报(2016年5期)2016-05-17
纺织服装流行趋势展望(2016年6期)2016-05-04
纺织服装流行趋势展望(2016年2期)2016-05-04
中国酿造(2016年12期)2016-03-01
