
人脑中脱碘酶相互作用蛋白的筛选与验证
2011-07-16 03:46张毅哲邵永红
深圳大学学报(理工版) 2011年5期
张毅哲,刘 琼,田 静,邵永红
1)深圳大学生命科学学院,深圳518060;2)深圳大学光电工程学院,深圳518060
硒是生物体必需的微量元素,缺硒会诱发许多重大疾病[1],补硒能降低克山病和大骨节病发病率.硒对阿尔茨海默症(Alzheimer's disease,AD)、精神分裂症和认知能力等也有影响[2-5].将硒用于缺硒大鼠,可维持其大脑硒水平,优先供给中枢神经系统[2-4].硒在脑内的生物功能主要通过各种硒蛋白实现,硒蛋白对维护脑的生长发育和预防脑神经细胞损伤具有重要作用[5].脱碘酶(iodothyronine deiodinases,DIO)是哺乳动物大脑中的一种重要硒蛋白,它在大脑中的生物功能及作用机制目前尚不清楚.脱碘酶3(DIO3)为3种类型脱碘酶中的一种[6],是使三碘甲状腺原氨酸(T3)、四碘甲状腺原氨酸(T4)失活的主要脱碘酶[7].因此筛选人脑中与DIO3相互作用的蛋白,探索DIO3作用途径及生物功能,有助于揭示硒预防脑疾病的分子机理.
DIO3具有硒蛋白基因结构特征,即其活性中心的重要氨基酸硒代半胱氨酸 (selenocysteine,Sec)由传统终止码TGA编码.为使DIO3能在酵母中表达,需将该TGA码突变为编码半胱氨酸(cysteine,Cys)的密码子,把基因突变对蛋白结构的影响降到最低.本研究采用酵母双杂交技术,以DIO3基因截短突变体为“诱饵”,筛选人胎脑cDNA文库,寻找人脑中与其相互作用的蛋白,并用荧光共振能量转移法 (fluorescence resonance energy transfer,FRET)进行验证.
1 实验
1.1 试剂和材料
Matchmaker Y2HGold酵母双杂交系统及人胎脑cDNA文库,酵母缺陷型氨基酸SD/-Leu、SD/-Trp、SD/-Ade/-Trp/-Leu、SD/-Ade/-His3/-Trp/-Leu混合物,X-α-Gal和 抗 真 菌抗 生 素 Aureobasidin A(AbA)均购自Clontech公司;人DIO3基因突变体、HEK293T细胞和Top10大肠杆菌为本实验室保存;DNA限制性内切酶、Taq酶和T4 DNA高效连接试剂盒购自Takara公司;酵母提取物、琼脂粉和酵母培养 (yeast nitrogen base without amino acids,YNB)基本氮源购自Difco公司;蛋白胨、3-氨基-1,2,4- 三唑和 212 ~300 μm 玻璃珠购自 Sigma公司;Tris和PEG3350购自Merck公司;pECFP-C1载体 (cyan fluorescent protein,CFP)和pEYFP-C1载体 (yellow fluorescent protein,YFP)由深圳大学田生礼副教授惠赠;DNA-MATE转染试剂购自深圳博纳泰克公司.主要仪器有共聚焦显微镜专用培养皿 (直径35 mm,Merck公司)、共聚焦显微镜(Olympus,FV1000)和PCR仪 (广州仪涛科学仪器有限公司,2720型).
1.2 人DIO3基因突变体的克隆及质粒构建
根据 DIO3基因(NM_001362.3)的编码序列(837碱基对 (base pair,bp)),以购自Gibco公司的成人肝cDNA文库质粒为模板,扩增人DIO3全基因,克隆后测序验证读码框准确性.以DIO3全基因为模版,采用重叠PCR技术,使DIO3基因中TGA码突变为TGC码,测序验证读码框准确性.将经DNA限制性内切酶SfiⅠ酶切后的DIO3突变截短体,用T4连接酶链接到NpGBKT7载体上,
获得诱饵质粒 NpGBKT7-DIO3.把 NpGBKT7-DIO3质粒转化到感受态大肠杆菌Top 10中,用含有卡那霉素的LB培养基筛选阳性克隆.
1.3 酵母转化
挑取酵母菌3~4个克隆 (菌落直径2~3 mm),接种于液体酵母完全培养基 (yeast peptone dextrose adenine,YPDA)中,30℃、250 r/min振荡培养16~18 h至 OD600≥1.5.利用 PEG/LiAc法制备新鲜的酵母感受态细胞,具体方法参见Clontech公司酵母操作手册;转入已构建好的诱饵蛋白质粒0.1 mg,同时设置阳性对照 (分别转入质粒pGBKT7-p53和pADT7-T)和阴性对照 (分别转入质粒 pGBKT7-Lam和pADT7-T).用200 μL双蒸水重悬酵母细胞,铺在缺陷型培养基SD/-Trp/X-α-Gal/AbA固体培养基上 (直径90 mm),同时将阳性对照和阴性对照组铺于SD/-Leu/-Trp/X-α-Gal/AbA固体培养基上,30℃倒置培养3~5 d,得到酵母诱饵菌株NpGBKT7-DIO3.将试验组与阳性对照及阴性对照组比较,通过酵母生长状态和酵母菌落是否变蓝排除试验组的自激活作用.
1.4 文库筛选
挑选在 SD/-Trp/X-α-Gal的 AbA固体培养基(含抗真菌抗生素)上生长的含有NpGBKT7-DIO3质粒的酵母菌落 (直径>2 mm),接种于50 mL SD/-Trp液体培养基中,30℃、250 r/min振荡培养16 ~18 h,至 OD600>1.5.再转移至 300 mL YPDA液体培养基中,培养至OD600值为0.4~0.6.采用PEG/LiAc/TE法,转入文库质粒50 μg.铺在SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA固体培养基平板上,30℃倒置培养1周.
1.5 酵母细胞内回转验证
从 SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA 固体培养基平板上挑取阳性克隆(菌落直径>2 mm),接种于5 mL SD/-Leu液体培养基,30℃、250 r/min培养过夜,从酵母中提取质粒,再转化到感受态大肠杆菌Top10中,用含有氨苄霉素的LB培养基筛选,并提取质粒(载体为pACT2),再与NpGBKT7-DIO3质粒共转化感受态Y2HGold酵母细胞,铺在SD/-Leu/-Trp/X-α-Gal/AbA固体培养基上,30℃倒置培养2~3 d,观察菌落是否变蓝.对筛选变蓝的菌落所转的质粒进行测序,应用美国国立生物技术信息中心 (National Center for Biotechnology Information,NCBI)等公共数据库对序列进行生物信息学分析,确定所筛选基因的名称、归属、同源相似率、启始码、终止码和编码区等基因相关信息.
1.6 FRET技术验证
重组质粒的构建 分别以质粒NpGBKT7-DIO3和pACT2-SERPINA3为模版进行PCR扩增,将获得的两段基因分别经EcoRⅠ和BamHⅠ双酶切,纯化回收后分别用T4连接酶连接到质粒pECFP-C1和pEYFP-C1上,构建重组质粒pECFP-C1-DIO3截短突变体和pEYFP-C1-SERPINA3截短体,测序验证读码框正确.
细胞转染 按2×105/孔接种HEK 293T细胞至共聚焦专用培养皿,待细胞长至80%融合时单转pECFP-C1-DIO3截短突变体,单转pEYFP-C1-SERPINA3截短体,共转染pECFP-C1-DIO3截短突变体和pEYFP-C1-SERPINA3截短体.以单转 pECFPC1、单转pEYFP-C1及共转pECFP-C1和pEYFP-C1为对照组.具体方法为:将DNA及转染试剂溶液混合,室温孵育30 min;将小皿中培养基换用2 mL无抗无血清DMEM细胞培养基,加入转染混合液并轻柔混匀,置于CO2培养箱中 (37℃,CO2体积分数为5%)孵育;4 h后弃去培养基,换用含10%小牛血清的DMEM(Dulbecco's modified Eagle's medium)继续培养.
敏化发射 (sensitized emission)法检测 转染48 h后,取培养皿置于激光共聚焦显微镜载物台上,分别以405 nm(检测供体pECFP-C1的青色荧光)和515 nm(检测受体pEYFP-C1的黄色荧光)为激发波长,在激光共聚焦显微镜下观察融合蛋白pECFP-C1-DIOⅢ截短突变体和 pEYFP-C1-SERPINA3截短体的共定位表达.采用 FV10-ASW 2.1 Viewer软件,计算FRET效率E、供体与受体之间的距离D.pECFP-C1-DIOⅢ截短突变体和pEYFPC1-SERPINA3 的 Foster距离 R0=5.276 7μm.
荧光寿命法检测 转染48 h后,采用时间相关单光子计数荧光寿命法分别测定pECFP与pEYFP共转 HEK293T,pECFP-DIO3'与 pEYFP-SERPINA3共转HEK293T,两组样品的荧光寿命.采用激光共聚焦显微镜、405 nm激光器、TCSPC采集卡及其配套SPC采集软件和图像分析软件进行图像采集和相关数据的分析与计算.
2 结 果
2.1 DIO3基因片段的克隆与测序
由于DIO3基因中有一个硒代半胱氨酸,正常情况下会导致翻译终止,要想得到真正具有生物活性的DIO3蛋白质,并利用酵母双杂交技术研究其相互作用,必须将硒代半胱氨酸突变成半胱氨酸.同时,由于DIO3基因全长的N端有一段跨膜结构域,而这段区域会阻碍酵母双杂交实验的进行,因此不扩增此区域,最终获得DIO3突变截短体基因片段,大小为729 bp.将随意挑选的 NpGBKT7-DIO3突变截短体重组质粒使用sfiⅠ限制性内切酶50℃酶切3 h后进行琼脂糖凝胶电泳,结果显示在729 bp处有目的条带出现 (图1),证明目的质粒构建成功.测序结果表明所扩增的目的基因片段序列与已报道序列完全匹配,编码框中编码硒代半胱氨酸的TGA码被突变为编码半胱氨酸的TGC码.
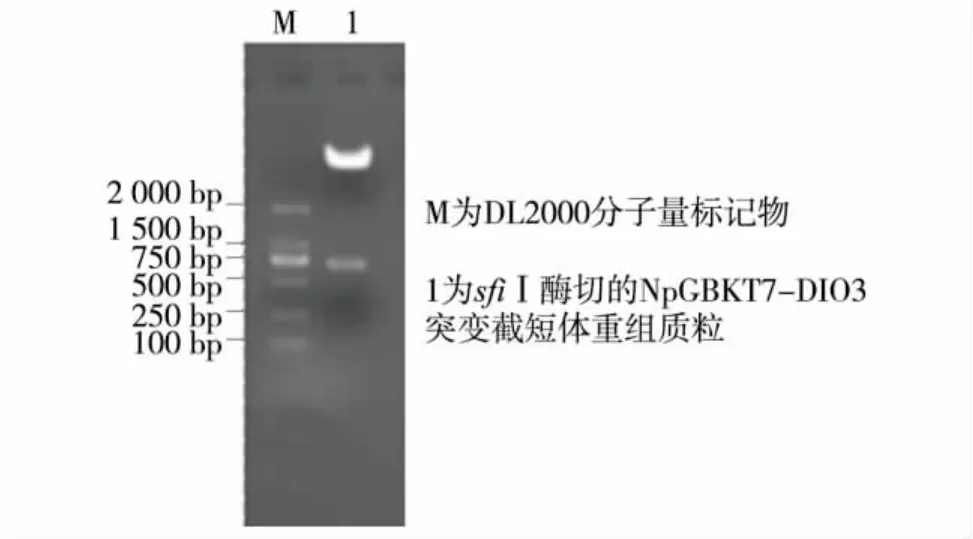
图1 DIO3基因片段重组质粒的酶切鉴定Fig.1 Enzymatic digestion of the recombinant plasmid NpGBKT7-DIO3
2.2 共转法筛选人胎脑cDNA文库
应用PEG/LiAc法分别将NpGBKT7-DIO3突变截短体与NpADT7空载共转化Y2HGold酵母菌空载涂布于SD/-Leu/-Trp/X-α-Gal/AbA平板,转化后2 d菌落生长饱满.根据与阳性对照组平板、阴性对照组平板的比较,NpGBKT7-DIO3突变截短体与pADT7空载共转化的酵母菌落颜色为白色,而阳性对照组平板上生长的酵母菌落为蓝色,且NpGBKT7空载单转的酵母菌落为白色,通过对比可知NpGBKT7-DIO3突变截短体本身没有自激活现象.
采用PEG/LiAc法共转pGBKT7-DIO3突变截短体质粒(100 μg)和Clontech人胎脑cDNA文库质粒(50 μg)进入Y2HGold酵母菌中,铺于直径150 mm的 SD/-Ade/-His/-Leu/-Trp/X-α-Gal/AbA平板上,培养7 d后,在板上有较大的菌落长出,见图2.
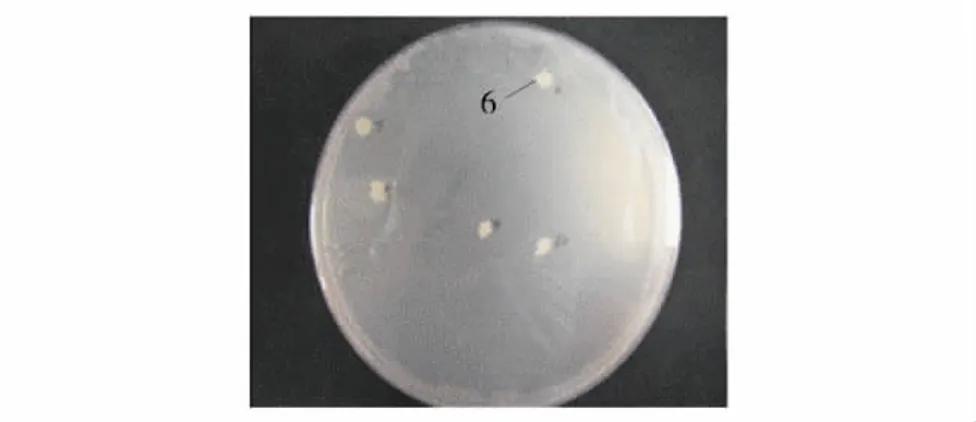
图2 人胎脑cDNA文库中与DIO3突变截短体相互作用的蛋白的筛选平板图Fig.2 Screening the interactive protein of DIO3 from human fatal brain cDNA library
挑选图2中的6号菌落,提取分离其中的AD质粒,转化Top10大肠杆菌,挑选阳性克隆送测序,并在NCBI上进行比对,结果表明,猎物基因为人丝氨酸蛋白酶抑制剂A族蛋白3(Homo sapiens serpin peptidase inhibitor,clade A,member 3,SERPINA3),基因登记号为 NM_001085.4.
2.3 回转酵母验证筛选所得蛋白相互作用
将NpGBKT7-DIO3突变截短体质粒和分离出来的pACT2-SERPINA3质粒共转进Y2HGold酵母菌中;同时用pGBKT7-p53和pADT7-T共转Y2HGold酵母菌作为阳性对照;用pGBKT7-Lam和pADT7-T共转酵母菌作为阴性对照.铺板3 d后,共转NpGBKT7-DIO3和pACT2-SERPINA3的一组以及阳性对照组在 SD/-Leu/-Trp/X-α-Gal/AbA平板上有较大的蓝色菌落长出,而阴性对照组的菌落小且没有变蓝,说明DIO3突变截短体和SERPINA3之间确实存在相互作用,见图3.
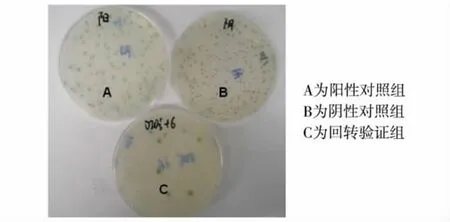
图3 酵母细胞内回转验证DIO3突变截短体与SERPINA3的相互作用Fig.3 Verification of DIO3 and SERPINA3 interaction by co-transformation of the two gene fragments into yeast cells
2.4 FRET技术验证蛋白质相互作用
敏化发射法检测蛋白质相互作用 为在活体细胞内检测蛋白质相互作用,将DIOⅢ突变截短体插入荧光蛋白表达载体pECFP-C1中,SERPINA3插入荧光蛋白表达载体pEYFP-C1中,构建CFP和YFP标记的融合蛋白表达载体pECFP-C1-DIOⅢ'和pEYFP-C1-SERPINA3截短体.
将 pECFP-C1-DIOⅢ'(5 μg)和pEYFP-C1-SERPINA3截短体分别单转HEK293T细胞;再将上述两质 粒 以 质 量 比 1∶1(均 为 2.5 μg) 共 转 染HEK293T细胞,共3个样品,转染24 h后,在共聚焦荧光显微镜下进行FRET检测.采集图像前,在显微镜下找到荧光表达适度的细胞,用固定的成像参数对其分别进行CFP通道和YFP通道成像.固定的成像参数设置如下:CFP通道,7%激光功率,HV为400;YFP通道,7%激光功率,HV为400.依据敏化发射方法要求,分别利用3个样品采集7张图像,以单转CFP空载、单转YFP空载、共转CFP空载和YFP空载的样品作为阴性对照组,选取了10个不同的感兴趣区域 (region of interest,ROI)进行测定.
根据FRET敏化发射法原理,如果代表供体和受体的两个荧光蛋白间存在相互作用,蛋白间距离应在10 nm之内.通过采集以下7张图 (图4),可计算蛋白质之间的距离及荧光共振能量转移效率.阴性对照组也同样采集上述7张图,单转质粒分别为不含相互作用蛋白基因片段的相应荧光蛋白载体,即pECFP-C1空载单转细胞、pEYFP-C1空载单转细胞、pECFP-C1和pEYFP-C1空载共转细胞.采用FV10-ASW 2.1 Viewer软件,根据实验中的原始数据,计算得出 pECFP-C1-DIOⅢ截短突变体与pEYFP-C1-SERPINA3截短体间的能量转移率和作用距离.
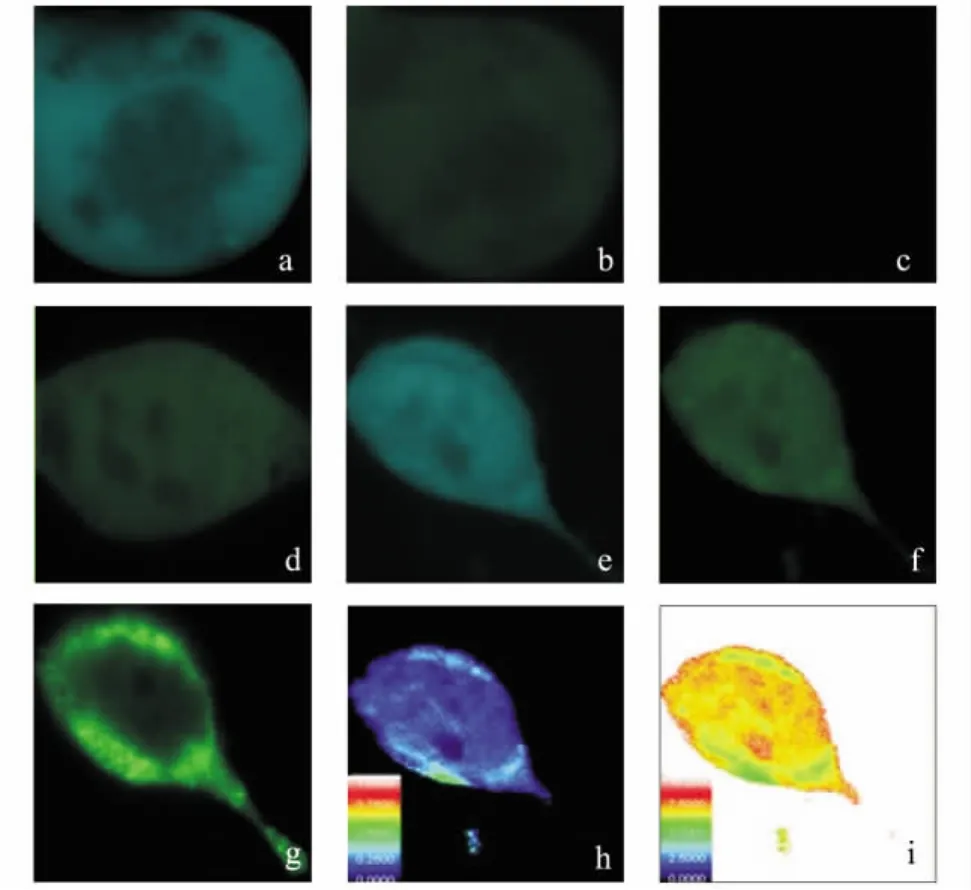
图4 FRET敏化发射法验证DIOⅢ与SERPINA3的相互作用Fig.4 Verification of the interaction between DIOⅢand SERPINA3 proteins by the method of sensitized emission of FRET
表1列出了所选取的10个不同ROI中测定的E和D.由表1可算出FRET平均效率E=23.4%,蛋白间的距离 D=6.632 nm.结果表明,pECFPC1-DIOⅢ截短突变体和pEYFP-C1-SERPINA3截短体在细胞内能够发生能量转移,证明DIOⅢ截短突变体与SERPINA3截短体确实存在相互作用.
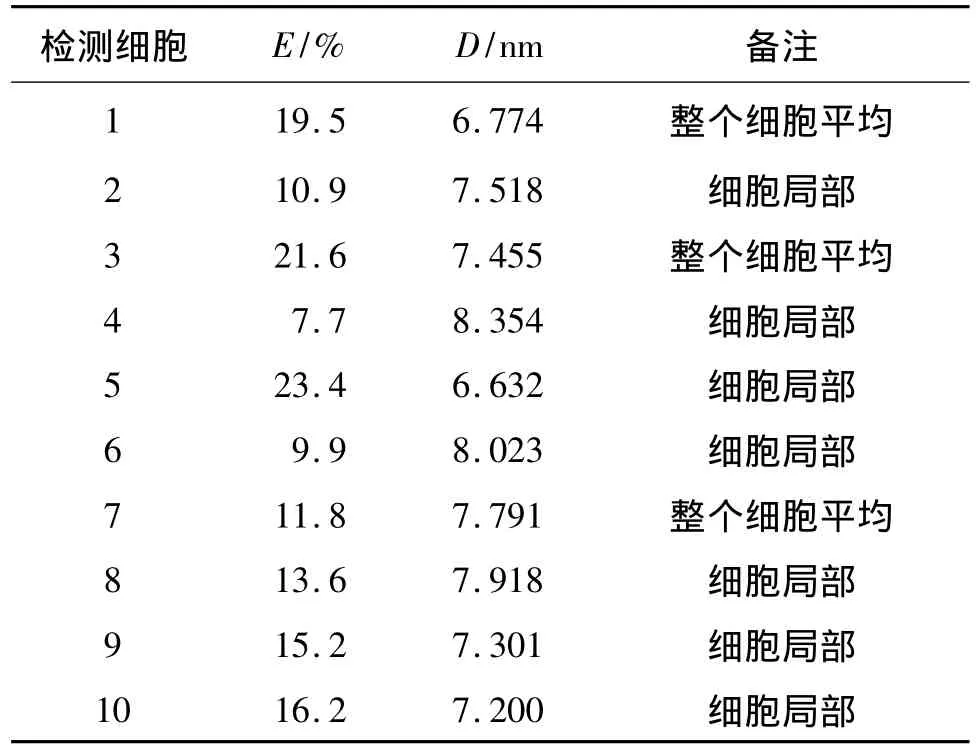
表1 不同细胞的荧光共振能量转移效率和蛋白间的距离Table1 The FRET efficiency and the distance between two interactive proteins
荧光寿命法检测蛋白质相互作用 为了在活细胞内检测是否存在上述蛋白质相互作用,将DIO3突变截短体插入荧光蛋白表达载体pECFP-C1,将SERPINA3插入荧光蛋白表达载体pEYFP-C1中,构建了CFP和 YFP标记的融合蛋白表达载体pECFP-C1-DIO3和 pEYFP-C1-SERPINA3截短体.采用时间相关单光子计数的荧光寿命法测定pECFP-C1与 pEYFP-C1共转化 HEK293T细胞、pECFP-DIO3突变截短体与pEYFP-SERPINA3共转HEK293T细胞,利用TCSPC采集卡配套的一个SPC采集软件,一个SPC image分析软件进行相关数据的分析与计算.图5为任意选取的一对阴性对照组和相互作用蛋白组的荧光寿命检测曲线.对照组与蛋白相互作用验证组的细胞均选取10个进行检测,每个细胞选取至少8个ROI.表2列出任意选取的8个ROI的荧光寿命值 (tm).表2列出图5中任意选取的8个ROI的tm.由图5和表2的荧光寿命值可见,共转DIO3突变截短体与SERPINA3的细胞与阴性对照组相比较,荧光寿命衰减的更快,说明DIO3突变截短体与SERPINA3间具有相互作用.

图5 基因转染后的HEK293T细胞的荧光寿命衰减曲线Fig.5 The decay curves of the fluorescence lifetime in the gene-transfected HEK293T cells

表2 DIO3突变截短体与SERPINA3共转细胞后不同ROI的荧光寿命值Table 2 The fluorescence lifetime of cells co-transfected with DIO3 and SERPINA3 genes 单位:ps
3 讨论
本研究采用酵母双杂交技术和FRET技术筛选和初步验证了与DIO3突变体相互作用的蛋白为SERPINA3.DIO3突变体中用 Cys取代了硒蛋白DIO3中的Sec,由于Cys和Sec的结构及性质均十分相近,因此能最大程度降低基因突变对蛋白质相互作用位点的改变.
SERPINA3是糖基化的丝氨酸蛋白酶抑制剂.有研究表明SERPINA3能加速Aβ原纤维的解聚,从而在机体内形成炎症反应导致 Aβ的聚集[8-9].SERPINA3充当分子伴侣的角色,能够增加Aβ的神经毒性,促使淀粉样蛋白丝的形成.转基因小鼠模型也证实SERPINA3既能够抑制Aβ的降解同时又能够促进Aβ在脑中的聚集[10].近年大规模基因组研究显示,SERPINA3的等位基因突变与AD及年龄依赖性神经退行性疾病密切相关[11-12].因此,SERPINA3与AD关系密切.
有报道称[5],缺硒直接导致老年人认知功能降低.DIO3是脑中主要表达的一种硒蛋白,对脑的发育十分重要[6],而硒对AD的作用途径目前尚不清楚.本研究结果表明,DIO3与SERPINA3的相互作用.DIO通常用活性中心的硒与含巯基的抑制剂发生作用[13],而SERPINA是丝氨酸蛋白酶和半胱氨酸蛋白酶抑制剂[14],因此推断DIO3与 SERPINA3的相互作用可能与脱碘酶上的硒醇基 (或突变体上的巯基)及SERPINA3涉及的丝氨酸和半胱氨酸相关联.然而真实作用机制有待今后深入研究,对上述蛋白质相互作用的认识将有助于揭示硒和DIO3在AD形成过程中的重要作用.
致谢:衷心感谢倪嘉缵院士对本实验整体方案设计的悉心指导!
[1]Hatfield D L.硒的分子生物学及其在人类健康中的作用[M].波士顿:Kluwer学术出版社,2001:283-297.(英文版)
[2]Gwona A R,Parka J S,Parka J H,等.硒降低β-淀粉样蛋白的产生及β-淀粉样蛋白诱导产生的神经元死亡[J].神经科学快报,2010,469(3):391-395.(英文版)
[3]Tung Y T,Hsuc W M,Wang B J,等.亚硒酸钠通过激活细胞外信号调节激酶抑制γ-分泌酶活性[J].神经科学快报,2008,440(1):38-43.(英文版)
[4]刘 琼,姜 亮,田 静,等.硒蛋白的分子生物学及与疾病的关系[J].化学进展,2009,21(5):819-830.
[5]GAO Su-juan,JIN Yin-long,Hall K S,等.中国农村老年人硒水平与认知功能[J].美国流行病学杂志,2007,165(8):955-965.(英文版)
[6]Schweizer U,Brauer A U,Kohrle J,等.硒与脑功能:一个少为人知的领域[J].脑研究综述,2004,45(3):164-178.(英文版)
[7]Hoppe B,Bräuer A U,Kühbacher M,等.脑细胞系及不同脑区中硒蛋白表达的生化分析[J].细胞组织研究,2008,332(3):403-414.(英文版)
[8]Kordula T,Bugno M,Rydel R E,等.人星形胶质细胞中α1-抗糜蛋白酶基因的白介素-1和肿瘤坏死因子依赖性调节机制[J].神经科学杂志,2000,20(20):7510-7516.(英文版)
[9]Perrin R J,Fagan A M,Holtzman D M.阿尔茨海默症的多种诊断和预测技术[J].自然,2009,461:916-922.(英文版)
[10]Janciauskiene S,Rubin H,Lukacs C M,等.阿尔茨海默症的Aβ1-42肽与α-抗糜蛋白酶的两个β-折片相结合将其从抑制剂转为底物[J].生物化学杂志,1998,273(43):28360-28364.(英文版)
[11]Cacabelos R,Martinez-Bouza R.痴呆症的基因组学和药物基因组学[J/OL].神经科学与疗法,2010,1-11[2010-08-16].http://onlinelibrary.wiley.com/doi/10.1111/j.1755-5949.2010.00189.x/pdf.(英文版)
[12]Soler-López M,Zanzoni A,Lluís R,等.相互作用组图谱揭示阿尔茨海默症的详细潜在机制基因组研究[J].2011,21(9):364-376.(英文版)
[13]Robertson J G.酶靶药物的作用基础[J].生物化学,2005,44(15):5561-5571.(英文版)
[14]Silverman G A,Bird P I,Carrell R W,等.丝氨酸酶抑制剂是结构相似但功能多样的蛋白超家族——其进化、抑制机理、新功能和名称修正[J].生物化学杂志,2001,276(36):33293-33296.(英文版)
猜你喜欢
The Crop Journal(2019年6期)2019-12-20
生命科学研究(2018年1期)2018-05-29
中国调味品(2017年2期)2017-03-20
安徽医科大学学报(2016年12期)2017-01-15
创新作文(小学版)(2016年16期)2016-11-11
现代检验医学杂志(2016年5期)2016-08-20
天津医科大学学报(2015年2期)2015-12-22
烟草科技(2015年8期)2015-12-20
中国科技信息(2015年2期)2015-11-16
山东医药(2015年40期)2015-02-28
