
大白菜抽薹过程叶色转变前后的生理变化
2011-08-07 09:42韩丹萍李春刚张耀伟
中国蔬菜 2011年20期
韩丹萍 李春刚,2 张耀伟*
(1东北农业大学园艺学院,黑龙江哈尔滨 150030;2黑龙江省红旗农场,黑龙江哈尔滨 150088)
大白菜〔Brassic campestrisL. ssp.pekinensis(Lour.)Olsson〕属于二年生蔬菜作物,需要经历一个低温阶段才能顺利完成发育转变;萌动的种子或幼苗感受低温通过春化后,在一定的日照条件下即可由营养生长转到生殖生长,进而抽薹开花。国内学者从春化温度、光周期反应、春化和抽薹生理机制、耐先期抽薹鉴定与遗传方面对大白菜春化与抽薹特性进行了广泛研究(奥岩松和李式军,1997;刘春香 等,2001;余阳俊 等,2004;赵香梅 等,2005;夏广清 等,2006;张鲁刚 等,2007)。
大白菜通过春化至抽薹,叶片颜色由绿转灰,之后显蕾,进而抽薹开花,这个过程在弱冬性品种上表现尤为突出。有关作物春化到抽薹开花期间发生的叶色转变现象及其发生机制的研究尚未见报道。本试验通过探讨不同冬性的大白菜品种抽薹过程中不同时期的蛋白质、激素、色素含量的差异,从各种激素、色素含量变化和蛋白质组分变化方面探讨大白菜抽薹过程中发生叶色转变的生理机制。
1 材料与方法
1.1 试验材料
供试材料为东北农业大学大白菜课题组选育的 4个不同冬性大白菜高代自交系:弱冬性的A151、A8-1-1和强冬性的C13-1、C30。
1.2 试验方法
1.2.1 春化处理 试验于2009~2010年春季在东北农业大学设施中心和黑龙江省教育厅寒地蔬菜生物学重点实验室进行。4个大白菜自交系种子播于8 cm×8 cm营养钵内;子叶展平后放在光照培养箱内进行春化处理,温度为8 ℃/3 ℃(昼/夜,下同)、光照16 h/8 h;低温春化25 d后,于5月20日定植露地,使其在适温长日照下生长。在实体解剖镜下观察其花芽分化情况,在解剖镜下生长点可见数个透明无茎花芽突起即为花芽分化期,当花芽突起伸长出现花茎时即为花茎分化期,新叶颜色由绿转灰时即为叶色转变期,肉眼可见花蕾时为花茎伸长期。
分别在低温春化处理前和处理后、花芽分化期、花茎分化期、叶色转变期、花茎伸长期取样,取植株生长点部位新生幼嫩叶片测定叶片中色素、激素含量,并进行可溶性蛋白电泳分析。
1.2.2 色素含量测定 色素提取采用丙酮法,以80%丙酮为空白,在波长663、645、470 nm处测定光吸收值。3次重复。
1.2.3 激素含量测定 取0.5 g鲜样,3次重复,用液氮冷冻后置于-80 ℃保存。内源激素(IAA、ABA、GA3)采用高效液相色谱法(美国Waters HPLC系统)(罗正荣 等,1990)测定。色谱条件:色谱柱为Sperisorb C18,ID 4 nm×250 mm,5 μm;预柱为C18,ID 4 nm×30 mm,柱温为40 ℃,0.8 mL·min-1恒流洗脱;检测波长为207 nm。流动相为乙腈∶甲醇∶60%乙酸=5V∶50V∶45V。
1.2.4 可溶性蛋白分析 可溶性蛋白提取采用尚礼邰等(1993)的方法,SDS-PAGE采用陈钧辉等(2003)的方法。电泳后,把胶放入脱色液微波加热30 s,在脱色摇床上摇5 min至条带很淡,然后换染色液微波加热30 s,摇至条带清晰,然后用凝胶成像系统照相。
1.2.5 数据处理方法 色素和激素含量采用DPS数据处理系统(唐启义,2010)进行方差分析,用Duncan’s新复极差法进行显著检验。
2 结果与分析
2.1 大白菜抽薹过程中色素含量的变化
2.1.1 大白菜抽薹过程中3种色素含量变化分析 从图1、2可以看出,强冬性品系C13-1和C30的叶绿素a、叶绿素b、类胡萝卜素含量在花芽分化期均处于较低水平,而后迅速增加,在花茎分化期达到最高,随后下降,至叶色转变期达到低值,随后升高;弱冬性品系A151和A8-1-1的 3种色素含量均在花茎分化期降到最低,在花茎伸长期达到最高。即强冬性品系色素含量最高值出现的时期恰是弱冬性品系色素含量最低值出现的时期;另外,强冬性品系从花茎分化期开始3种色素含量均显著高于弱冬性品系。

图1 大白菜抽薹过程叶绿素a、叶绿素b含量变化
2.1.2 大白菜抽薹过程中色素间比值变化分析大白菜抽薹过程中色素之间的比值变化分析结果表明(图3):供试 4个大白菜自交系中除A8-1-1叶绿素 a/b的值外,其他均在叶色转变期最高,而A8-1-1在叶色转变期叶绿素a/b的值也高于前期,在花茎伸长期达到最高。而叶绿素(a+b)/类胡萝卜素的值变化较为复杂,总体上是春化后比值比春化前稍微增高,花芽分化期下降,此后比值逐渐增高,在叶色转变期达到最高,花茎伸长期稍微下降。
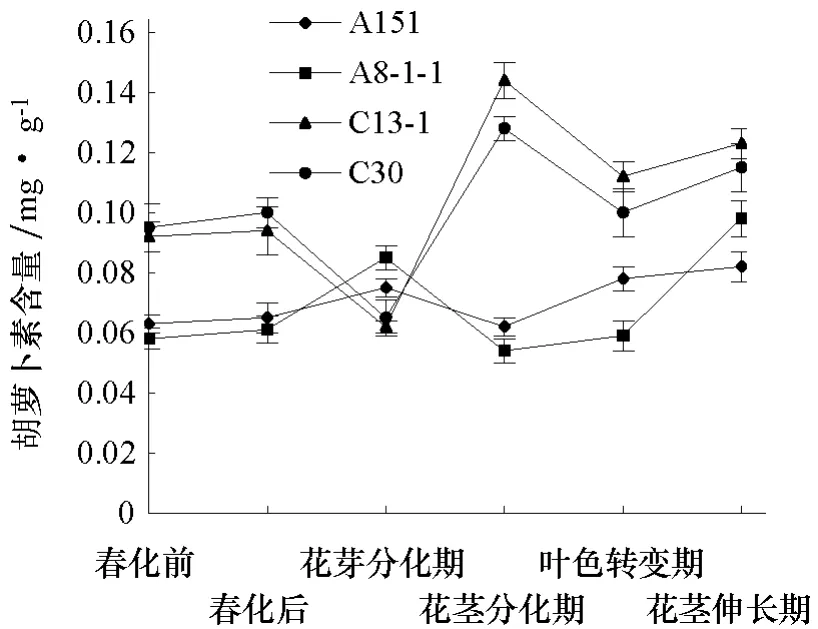
图2 大白菜抽薹过程胡萝卜素含量变化
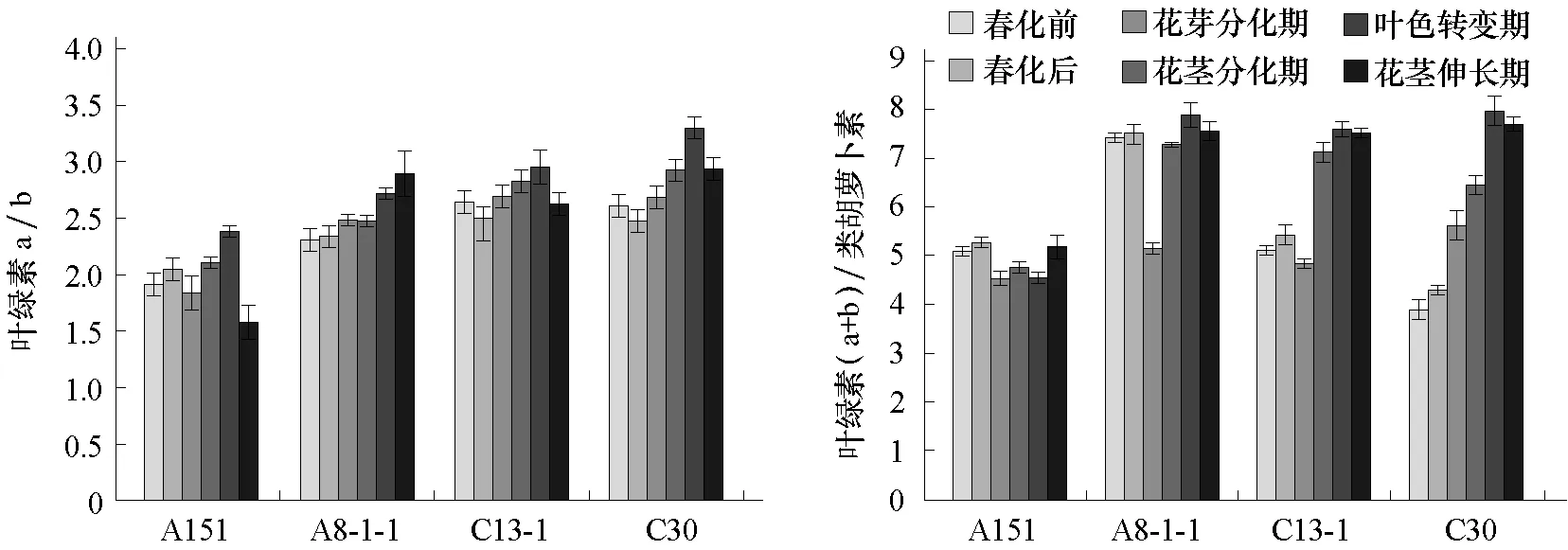
图3 大白菜抽薹过程叶绿素a/b、叶绿素(a+b)/类胡萝卜素变化
2.2 大白菜抽薹过程中激素含量变化
由图4可以看出,不同冬性的大白菜品系抽薹过程中IAA含量均出现两个峰值,分别在花芽分化期和叶色转变期,在叶色转变期达到最高,而且在花芽分化期与叶色转变期随品系冬性的增强,IAA含量逐渐降低。
不同冬性的大白菜品系 GA3含量变化趋势基本一致,在花芽分化期稍有降低,而随着显蕾期的临近GA3含量逐渐上升,在叶色转变期达到最大值,而且此时强冬性的品系其GA3含量低,其后伴随花茎的伸长GA3含量逐渐下降。
从图5可以看出,不同冬性的大白菜品系ABA含量变化趋势也基本一致,除在花茎分化期稍降低外,随着发育进程ABA含量越来越高,在花茎伸长期达到最大值,同期强冬性品系ABA含量高于弱冬性品系。
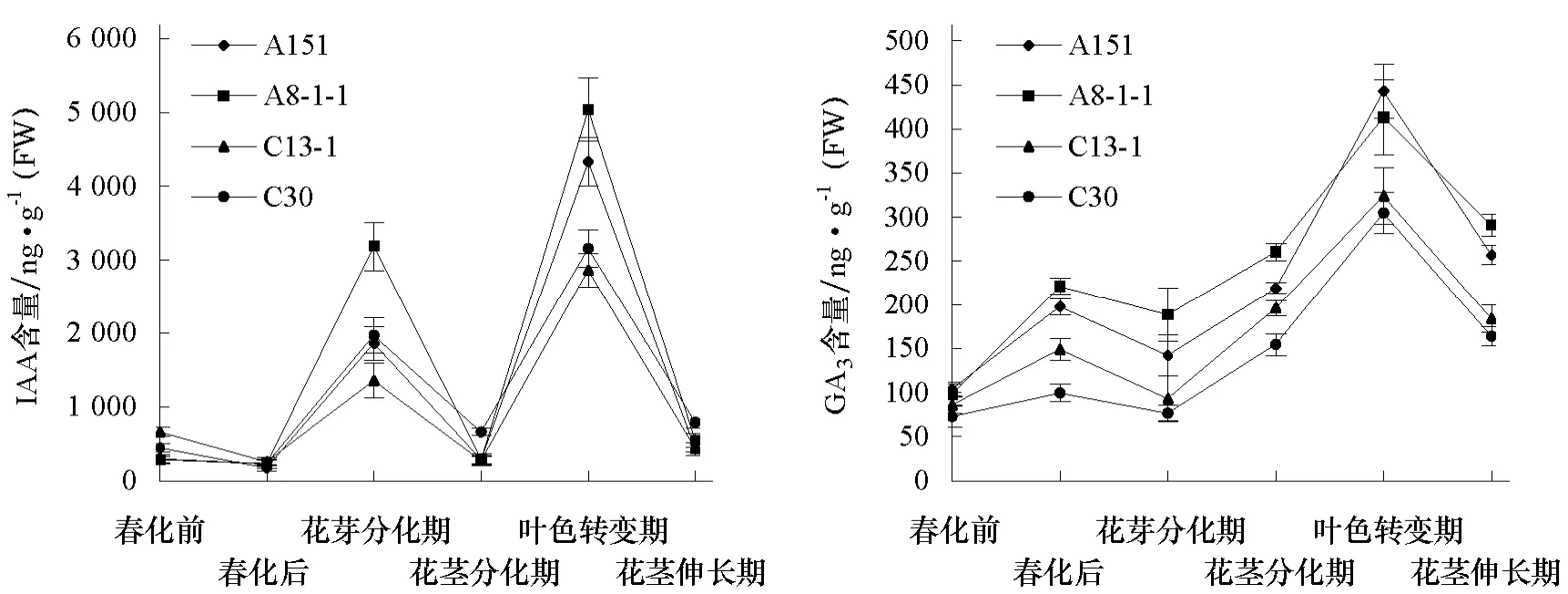
图4 大白菜抽薹过程中IAA、GA 3含量变化
2.3 大白菜抽薹过程中可溶性蛋白分析
由图6可以看出,不同冬性的大白菜品系抽薹过程中可溶性蛋白表达水平不同,其中分子量约为51 kD的蛋白质表达量最大,其在不同冬性材料的各个阶段表达,但在春化前表达微弱,在春化后表达量骤然上升,随着营养生长到生殖生长的转变其表达量又有所下降。该蛋白质在弱冬性品系中表达量要比强冬性品系稍高一些。另外分子量约为 18 kD的蛋白质在各品系春化前不表达,春化后微弱表达,随着生长发育表达量逐渐增强,在叶色转变期表达量达到最大,随后在花茎伸长期只有微弱表达或者不表达。
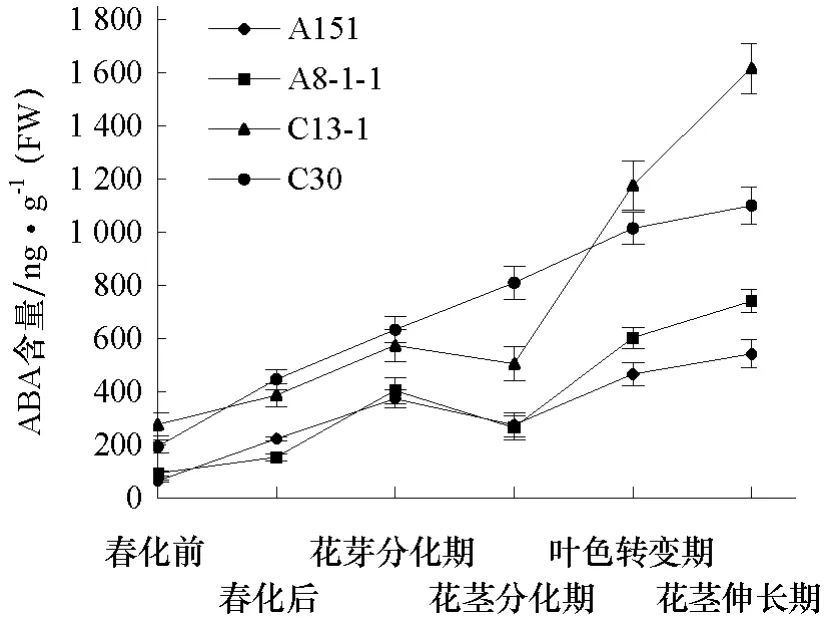
图5 大白菜抽薹过程中ABA含量变化

图6 大白菜抽薹过程中可溶性蛋白分析
3 结论与讨论
从本试验结果可以看出,强冬性的品系从花茎分化时期开始3种色素含量均显著高于弱冬性品系,强冬性品系色素含量最高值出现的时期恰是弱冬性品系色素含量最低的时期,同时发现抽薹过程中叶绿素a/b的值在叶色转变期达到一峰值,有理由推断其与叶色转变存在一定关系。
本试验还发现,不同冬性的大白菜品系抽薹过程中IAA含量在花芽分化期和叶色转变期有2个峰值,与王淑芬等(2003)研究在萝卜抽薹开花过程中,IAA含量大约在花芽分化期和抽薹前期出现 2个峰值的结果相似;而且与夏广清等(2006)对不同生态型的大白菜抽薹时内源激素含量研究结果比较一致的是发现冬性越弱的品种IAA含量越高,说明IAA的增加有利于大白菜花芽分化,能够促进大白菜由营养生长向生殖生长的转变,同时也促进抽薹。
Pharis和King(1985)研究发现赤霉素在低温春化抽薹开花过程中发挥着重要的调节作用。本试验中发现大白菜抽薹过程中 GA3含量在花芽分化期相对较低,而随着显蕾期的临近含量逐渐上升,在叶色转变期达到最大值,而且冬性越强的品系赤霉素含量越低,说明大白菜抽薹前夕GA3有一临界浓度,达到这一临界浓度则促进大白菜抽薹开花。
关于ABA是否参与或影响花芽分化及抽薹开花的认识不一致。本试验发现ABA含量在大白菜抽薹过程中叶色转变后、花茎伸长期达到峰值,并且冬性越强的品系ABA的含量越高,说明较高浓度的ABA可能促进大白菜抽薹开花。
对应植株任何一种形态上的变化,实际上都是由于植株体内蛋白质的变化所引起的,大白菜发育过程中形态发生变化源于植株体内蛋白质发生变化。本试验中蛋白质电泳分析结果表明,一种分子量约为18 kD的蛋白质在叶色转变期表达量最大,可能与叶色转变、抽薹有关,但其是否为成花特异蛋白质,还需进一步分离纯化并对其进行结构和功能研究。
综合分析,大白菜抽薹过程中,叶绿素a/b的值、IAA和GA3含量均在叶色转变期出现峰值,叶色转变后开始花茎伸长,说明叶色转变是大白菜抽薹的一个生理信号。
奥岩松,李式军.1997.大白菜发育过程中可溶性蛋白质的变化.中国蔬菜,(2):19-21.
陈钧辉,陶力,李俊,朱婉华,袁玉荪.2003.生物化学实验.北京:科学出版社:110-114.
刘春香,韩玉珠,栗长兰,张汉卿.2001.大白菜的春化特性及未熟抽薹研究进展.吉林农业大学学报,23(3):61-64.
罗正荣,朱丽华,吴谋成.1990.植物组织中赤霉酸含量的高效液相色谱测定.植物生理学通讯,(2):50-52.
尚礼邰,乔新芝,石晓东.1993.豆科植物可溶性叶蛋白的测定和提取.生物学通报,(11):45.
唐启义.2010.DPS数据处理系统.北京:科学出版社:85-87.
王淑芬,徐文玲,何启伟,刘继宝,张传坤.2003.春化深度对萝卜抽薹的影响及抽薹过程GA3和IAA含量的变化.山东农业科学,(6):23-27.
夏广清,何启伟,王翠花.2006.赤霉素处理对大白菜开花的诱导效应.中国蔬菜,(5):19-20.
余阳俊,张凤兰,赵岫云,张德双,张振贤.2004.大白菜晚抽薹性快速评价方法.中国蔬菜,(6):16-18.
张鲁刚,孔小平,惠麦侠,张明科.2007.大白菜新品种‘冠春’的春化特性及其遗传表现.西北农林科技大学学报:自然科学版,35(2):93-96.
赵香梅,孙守如,张晓伟,原玉香.2005.大白菜春化与抽薹特性的研究进展.中国蔬菜,(1):33-35.
Pharis R P,King R W.1985.Gibberellins and reproductive development in seed plants.Annual Review of Plant Physiology,36:517-568.
郭战玲,沈阿林,寇长林,马正华,王守刚.2008.不同小麦品种开花后硝酸还原酶活性与氮效率的关系.中国农学通报,24(5):219-223.
顾明霞.2010.钾素营养对烟株氮代谢及烟叶品质形成的影响.中国农学通报,26(9):214-219.
郝建军,刘延吉.2001.植物生理学试验技术.沈阳:辽宁科学技术出版社:64-67.
韩延,周世民,刘艳英,刘国顺,符云鹏.2003.氮用量对香料烟生理特性及品质的影响.烟草科技,(4):3-6.
黄勤妮,印利萍,蔡晓清.刘祥林.洪剑明.1995.不同氮源对小麦幼苗谷氨酰胺合成酶的影响.植物学报,39(5):856-862.
李海波,李全英,陈温福,孟雷.2003.氮素不同用量对水稻叶片气孔密度及有关生理性状的影响.沈阳农业大学学报,34(5):340-343.
李文华,赵明珠.2009.不同类型大豆品种氮同化和蛋白质积累规律的研究〔硕士论文〕.哈尔滨:东北农业大学.
刘春玲,陈慧萍,刘娥娥,彭新湘,卢少云,郭振飞.2003.水稻品种对几种逆境的多重耐性及与ABA的关系.作物学报,29(5):725-729.
刘玉梅.2009.不同施氮水平对嫁接和自根黄瓜生长和生理的影响.安徽农业科学,37(26):12481-12484.
林清华,李常键,彭进,张楚富,Peng S B,Bennett J.2000.NaCl对水稻谷氨酸合酶和谷氨酸脱氢酶的胁迫作用.武汉植物学研究,18(3):206-210.
欧吉权,魏国威,张楚富,张吉,林清华.2003.甜瓜子叶发育过程中蛋白质水平和氨同化酶活性变化.武汉大学学报:理学版,49(4):538-542.
尚庆茂,易旸.2009.水杨酸对水分胁迫下黄瓜幼苗的光合及氮代谢的调节作用〔硕士论文〕.北京:中国农业科学院.
杨东,游晴如,谢鸿光,涂诗航,董瑞霞,张水金.2008.氮肥水平对超级稻优航1号生长中后期生理生化特性的影响.江西农业大学学报,30(1):7-10,30.
闫桂萍.1994.不同供氮水平甜菜GS、GOGAT与GDH活性动态的研究〔硕士论文〕.哈尔滨:东北农业大学.
于海彬,蔡葆,王桂岚.1995.甜菜氮营养对蔗糖积累转化及有关酶活性的调节.中国甜菜糖业,(6):6-11,45.
周宝利,姜荷.2001.茄子嫁接栽培效果和抗病增产机制的研究进展.中国蔬菜,(4):52-54.
周宝利,尹玉玲,徐研,韩璐.2010.嫁接对茄子根际土壤微生物和叶片硝酸还原酶活性的影响.园艺学报,37(1):53-58.
张衍鹏,于贤昌,张振贤.2004.日光温室嫁接黄瓜的光合特性和保护酶活性.园艺学报,31(1):94-96.
张忠臣,金正勋.2010.插秧密度对寒地粳稻碳氮代谢关键酶活性及产量的影响.东北农业大学学报,41(10):5-9.
Cren M,Hirel B.1999.Glutamine synthetase in higher plant:regulation of gene and protein expression from the organ to the cell.Plant Cell Physiology,40:1187-1193.
Lea P J,Miflin B J.1979.The assimilation of ammonium nitrogen by ehloroPhylious tissue.Nitrogen Assimilation of Plant:475-486.
Srivastava H S,Singh R P.1987.Role and reguLation of L-glutamate dehydrogenase activity in higher plants.Phytochemistry,26:597-610.
猜你喜欢
蔬菜(2022年1期)2022-11-11
麦类作物学报(2022年4期)2022-05-23
孩子·小学版(2021年6期)2021-12-26
语文世界(小学版)(2021年5期)2021-06-04
浙江农业学报(2021年4期)2021-04-27
湖北农业科学(2020年1期)2020-05-09
山东林业科技(2019年2期)2019-06-03
江苏农业科学(2018年11期)2018-06-29
天然产物研究与开发(2018年4期)2018-05-07
山东农业科学(2018年3期)2018-03-29
