
小麦胚芽贮藏过程中的酸败变质机理
2011-11-14 07:15苗文娟徐山元
中国粮油学报 2011年9期
徐 斌 李 波 苗文娟 徐山元 董 英
(江苏大学食品与生物工程学院1,镇江 212013)
(发达面粉集团有限公司2,夏津 253216)
小麦胚芽贮藏过程中的酸败变质机理
徐 斌1李 波1苗文娟1徐山元2董 英1
(江苏大学食品与生物工程学院1,镇江 212013)
(发达面粉集团有限公司2,夏津 253216)
小麦胚芽含有丰富的优质蛋白、多不饱和脂肪酸和VE等多种营养素,同时也富含脂肪酶、脂肪氧化酶和酯酶等多种酶类。新鲜小麦胚芽贮藏两周左右,就会在各种酶和微生物的协同作用下酸败变质,从而极大限制了小麦胚芽的高效利用。本文系统阐述了小麦胚芽脂肪酶和脂肪氧化酶的种类、酶学特性及其催化机制;并从小麦胚芽脂质的脂肪酶催化水解、自动氧化、脂肪氧化酶催化氧化以及微生物污染等多角度分析了小麦胚芽贮藏过程中的酸败变质机理。对小麦胚芽脂肪酶和脂肪氧化酶的深入了解,有助于研发可靠的小麦胚芽稳定化技术与装备,为其进一步深加工创造条件。
小麦胚芽 脂肪酶 脂肪氧化酶 微生物 酸败 机理
小麦胚芽(Wheat Germ)为小麦生长发育的基础,富含各种必需氨基酸、不饱和脂肪酸、矿物质和维生素,被誉为“人类天然的营养宝库”,可广泛应用于食品、医药与饲料工业。然而,新鲜小麦胚芽富含多种酶类,如脂肪酶(Lipase,简称LA)、脂肪氧化酶(Lipoxygenase,简称 LOX)、酯酶、α -淀粉酶和蛋白酶等[1-2]。LA能催化小麦胚芽脂质中的三酰甘油水解成高级脂肪酸[3];LOX催化氧化小麦胚芽脂质中的亚油酸,生成氢过氧化油酸,经分解生成挥发性的醛和酮类等[4]。由于LA和LOX等酶的共同作用,使得小麦胚芽贮藏稳定性极差。此外,制粉过程中小麦胚芽常会受到微生物污染,微生物代谢产生的酶进一步加速了脂质水解与氧化进程[5-6]。常温下,新鲜小麦胚芽存放两个星期左右就会酸败、变质[7],从而限制了小麦胚芽的高效利用[8]。
1 小麦胚芽脂肪酶
1.1 脂肪酶
脂肪酶(EC3.1.1.3)又称甘油酯水解酶,它作用于脂质分子中的羧酸酯键,将三酰甘油水解为脂肪酸和二酰甘油、单酰甘油或甘油,其总体反应历程如图1所示。LA也可催化甘油酯及水不溶性酯类的醇解、酯化、酯交换以及合成等反应[9]。脂肪酶主要来源于微生物、动物和植物,目前已有数十种商业化脂肪酶出现[10],并广泛应用于食品、药品、化工、造纸等行业[11-12]。
LA的催化反应不需要辅酶,且反应条件温和、副产物少[13],反应通常具有高度立体异构专一性和化学选择性[14-15]。LA另一重要特征是只能在异相系统,即在油-水界面上作用,分解脂肪的能力高度依赖于酶与脂质的接触面积,称为界面活化现象[16]。

图1 三酰甘油的脂肪酶催化水解反应历程
1.2 小麦胚芽脂肪酶的种类与特性
谷物来源脂肪酶是植物脂肪酶中研究较多的一类,主要包括小麦、燕麦、大麦、稻谷和玉米脂肪酶等,因小麦是全球最主要的粮食作物之一,所以小麦脂肪酶是谷物脂肪酶中最受关注的一种[17-18]。
小麦脂肪酶主要分布在麸皮与胚芽两个部位,其中前者约占75% ~80%,胚芽中约占20% ~25%。麸皮中LA对热敏感,而胚芽中LA热稳定性较高。当温度升至75℃时,二者的酶活基本接近[19]。一般而言,小麦在未发芽或遭破坏时,其LA活力较低,约为2.17 ~9.42 U/g,且较为稳定[20];一旦发芽,在三酰甘油为底物的情况下其活力会提高数十倍,但三乙酸酯和甘油酯为底物时并非如此[1]。
1948年,Singer等[21]初步分离了小麦胚芽脂肪酶,并认为其是一种酯酶,只对短链甘油酯起反应,而对于中长链的基本无效。1966年,Staufer等[22]发现在小麦胚芽脂肪酶粗酶中表现解酯酶活性的实际上有3种酶:脂肪酶、酯酶及三丁酸甘油酯酶。脂肪酶具有明显的界面活性,该酶可水解三酰甘油,但不能水解乙酯,反应符合米氏方程,其对中长链脂肪酸,尤其是油酸反应效率最高[23]。该酶分子质量约(143 ±2)ku,最适温度37 ℃,热稳定性好,在pH 8.0、60~90°C下保温1 h能保持80%酶活[24]。另有报道,小麦胚芽脂肪酶pI值为5.4,其分子质量为42 ku,最适 pH 8.0,适宜 pH 7.0 ~9.0,对 pH 敏感,当pH 7时酶活下降了50%;最适pH下,当有20%果糖,蔗糖等保护剂存在时,其Tm值约为60℃[25]。当pH大于10,酶蛋白的α-螺旋自解旋,导致活性中心部位结构发生变化,酶活力丧失;该酶对水分活度具有较高的依赖性,水分活度为0.90时具有较高活力,同时,钙、镁离子对其有促进作用,钾、钠离子则无明显效果[26]。
酯酶不具有界面活性,能水解乙酯,但不能水解三酰甘油;反应不符合米氏方程[25]。该酶适宜pH 6 ~8,最适 pH 7.2 ~7.3,且对 pH 波动不敏感,当pH 6时还能保持80%酶活;对热较稳定,最适温度30℃,当温度升到50℃时,10 min内酶活损失50%[22]。
三丁酸甘油酯酶也具有明显的界面活性,该酶能同时水解三酰甘油和乙酯,反应基本符合米氏方程[25]。其对中短链脂肪酸,尤其是丁酸和己酸三甘油酯显示了最高活力;最适 pH 6.6~6.8,适宜pH 5.5 ~7.5,对 pH 敏感,当 pH 5.5 时,酶活约下降80%;对热不稳定,最适温度在20℃,当温度上升到25 ℃时,很快就完全失活[22]。
小麦胚芽脂肪酶具有LA的共同结构特征:具有α/β水解酶折叠结构;拥有Gly-X-Ser-X-Gly保守结构域,且Ser为催化三联体的一部分;催化中心由Ser-His-Asp三元基团组成,被一个具有两亲性的α-螺旋“盖子”所覆盖[27]。目前尚无小麦胚芽脂肪酶晶体结构的报道,Korneeva等[28]研究表明,LA活性中心由天冬氨酸或谷氨酸的羧基与组氨酸的咪唑基构成,丝氨酸的羟基在催化中起着关键作用;在苯甲基磺酰氟中培育1 h,其酶活下降70%以上;EDTA是 LA的非竞争抑制剂。另有报道,Ser174-His293-Asp230为其催化三联体,其核心结构与Rhizzomucor Miehei lipase丝状真菌的LA蛋白序列相似度超过35%[29]。共含350氨基酸,结构含约20%的α-螺旋和40%的β-螺旋折叠,其余均为回环[25]。小麦胚芽脂肪酶的催化机制和其他 LA相似,遵循乒乓 bi-bi机制[3]。
2 小麦胚芽脂肪氧化酶
2.1 脂肪氧化酶
1932年,Andre首次发现大豆豆腥味的产生是多不饱和脂肪酸的酶促氧化所致,其中关键酶即为脂肪氧化酶(EC1.13.11.12)[30],系统命名为 Linoleate -Oxygen Oxidoreductase,即亚油酸-氧氧化还原酶,俗称脂肪氧化酶或脂肪氧合酶[31]。
2.2 小麦胚芽脂肪氧化酶的种类与特性
1943 年,Sumner[32]首次报道小麦胚芽中存在LOX。1982年,Nicolas等[33]从小麦胚芽中分离获得3种LOX同工酶(L-1、L-2、L-3),凝胶过滤色谱测得它们的分子质量为90 000~95 000 u,SDSPAGE测得的分子质量为110 000 u。L-1比 L-2、L-3热稳定性好,3种同工酶和粗酶具有相同的最适pH范围6~6.5。
Shiiba等[34]报道小麦胚芽LOX包含4种同工酶(L-1、L-2、L-3、L-a),其中 L-3约占总量的50%,L-1、L-2、L-3这3种主要的同工酶最适pH均为5.5,其中 L-2范围较宽,为4.5~6.0,L -3对亚油酸、亚麻酸均具有较高的活性,而L-1与 L-2对亚麻酸氧化催化能力较弱;L-1、L-2、L-3的SDS-PAGE呈单一条带,阻滞系数为0.28;L-1、L-2、L-3具有相似的热敏性,在45℃具有最高的酶活,而75℃则完全被钝化。另有报道,即使在低水分条件下,LOX也表现出较高的活性[35]。
2.3 脂肪氧化酶的催化氧化机制
LOX主要催化3种不同类型的反应:(1)脂质的双加氧化反应(过氧化物酶反应);(2)氢过氧化脂质的次级转化(氢过氧化物酶反应)[36];(3)环氧白三烯的形成(白三烯合成反应)[37]。
LOX常见的底物为必需脂肪酸的3种同分异构体,亚油酸、亚麻酸和花生四烯酸[38],在植物中其底物主要是亚油酸和亚麻酸。亚油酸和亚麻酸上的加氧位置是C9和C13,在花生四烯酸上的加氧位主要是C5、C12和C15,也可在C8、C9和 C11位上加氧。LOX也可以三亚油酸甘油酯、三亚麻酸甘油酯或其他不饱和脂肪酸的类脂作为底物[39]。
LOX在有氧环境里催化底物起连锁反应形成氢过氧化物。完整的脂质酶促氧化包含四步骤:(1)酶激活;(2)有氧途径;(3)厌氧途径;(4)非酶途径。LOX的催化类似于多不饱和脂肪酸自动氧化,所不同的是LOX催化反应比自动氧化更快,对反应底物有选择性,且对作用底物的氧化程度不一样[40]。大豆LOX催化的脂肪酸氧化必须具有顺,顺-1,4-戊二烯结构、且其甲基终端C8处有一个亚甲基团(两个双键之间的亚甲基)[41]。这类脂肪酸只在C6处发生氧化,而亚油酸在C6和C10都能发生氧化反应。就亚油酸而言,其主要氧化产物是具旋光性的C9和C13位氢过氧化物的同分异构体[42]。在O2和LOX存在的具有顺,顺-1,4-戊二烯结构的不饱和脂肪酸中,LOX中的Fe3+可能参与了电子的迁移,LOX必须以氧化态存在(Fe3+),它能催化除去油酸或亚油酸C11亚甲基的氢,形成一个C13自由基,同时LOX转化为还原态(Fe2+)。在有氧和缺氧条件下,反应都能进行,产生的氢过氧化物可进一步形成其他产物[43]。
Tappel等[44]则认为,LOX所催化的亚油酸的氧化不同于亚油酸自动氧化过程,首先是亚油酸、氧和LOX形成复合体;然后在酶的表面形成一个双游离基活化体,即一个氢离子和一个电子从亚油酸上转移至氧分子上;双游离基在酶分子表面结合形成亚油酸过氧化氢;此过氧化氢物与酶分离并脱落下来,至于亚油酸的哪个不饱和碳原子首先受到O2的进攻由LOX特性决定(图2)。

图2 LOX催化亚油酸氧化的步骤
3 小麦胚芽贮藏过程中的酸败变质
制粉过程中,当胚芽从胚乳中分离出来后,由于受到磨辊挤压,小麦胚芽中脂质接触到高活性脂肪酶,在其作用下酸败水解成游离脂肪酸,导致新鲜小麦胚芽的货架期很短,不再适合人类食用。小麦胚芽脂质水解首先是由内源酶(LA)催化,如果小麦胚芽中混入了麸皮等杂质,微生物代谢的酶也会引起脂质水解[5]。小麦胚芽脂质水解伴随多种营养与感官品质变化,如肥皂味般的异味、酸度上升、功能特性变化、脂肪酸氧化的易感性增加。而游离脂肪酸会进一步的氧化酸败(LOX),产生自由基,并通过裂变,最终产生异味,营养价值下降。归纳起来,小麦胚芽酸败主要有三种类型:即水解酸败、酶促氧化酸败和自动氧化酸败[5],如图3所示。
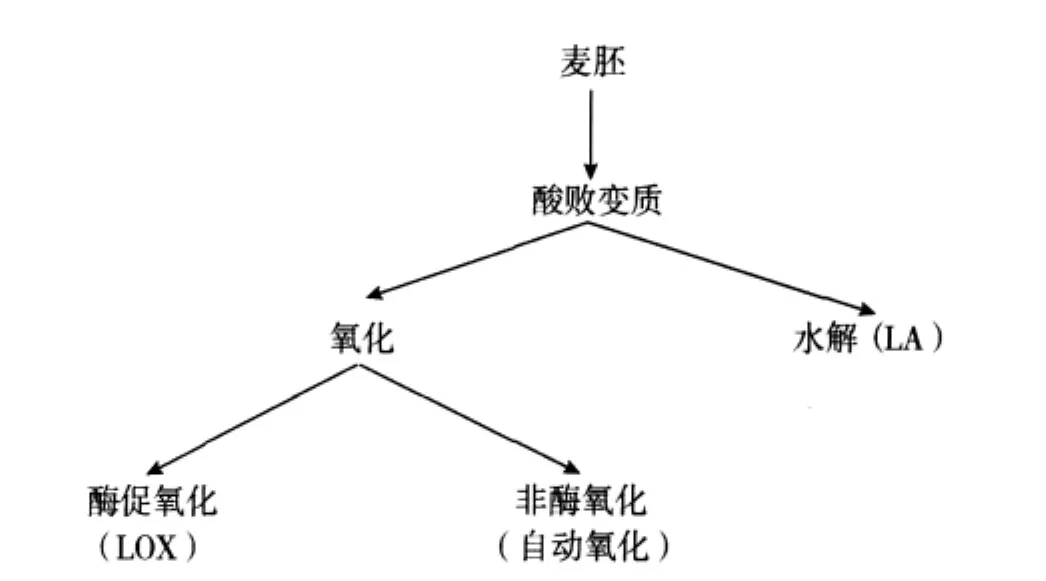
图3 小麦胚芽的变质途径
小麦胚芽中酶系统通常包括 LA、LOX、磷酸酶[45]、过氧化物酶[32]及乙醇脱氢酶[46]等,在它们的联合作用下,使小麦胚芽中的多元不饱和脂质降解生成小分子的醛、醇等(图4)。
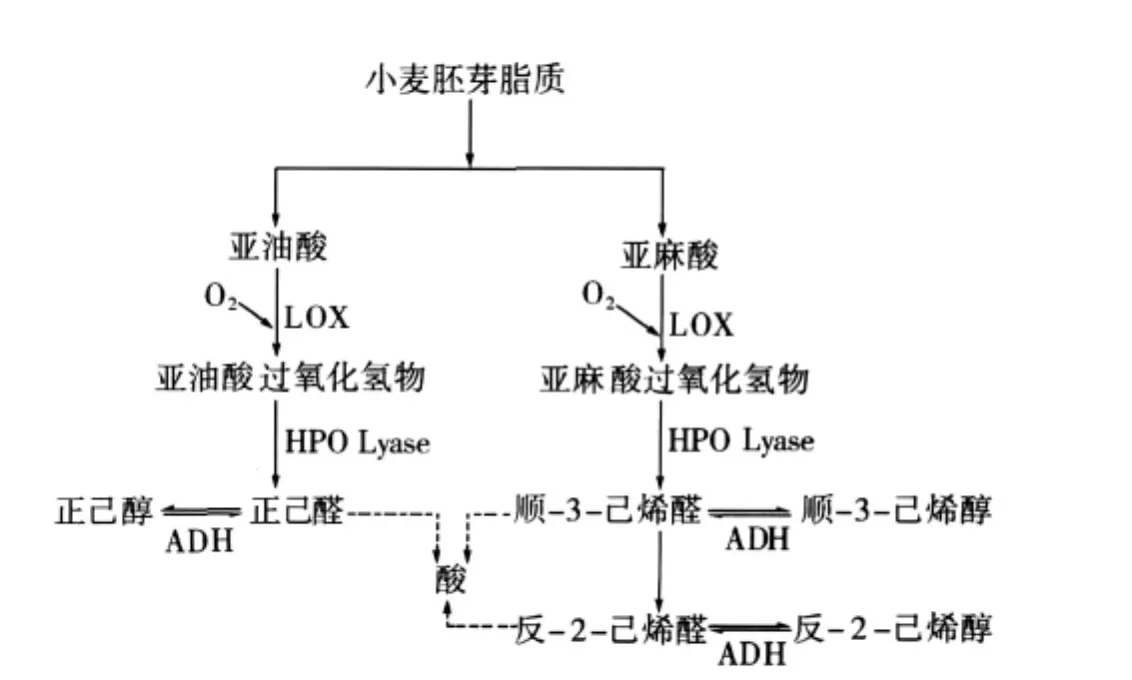
图4 从亚油酸和亚麻酸形成挥发性C6化合物的途径
3.1 小麦胚芽的酶促水解酸败
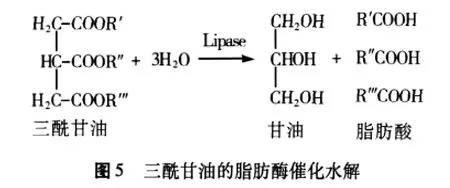
LA催化小麦胚芽油脂水解的一般路径为:小麦胚芽中的脂肪酶、三丁酸甘油酯酶等及微生物(如霉菌繁殖产生解脂酶)作用于三酰甘油,使之水解为脂肪酸、二酰甘油、单酰甘油和甘油(图5)。

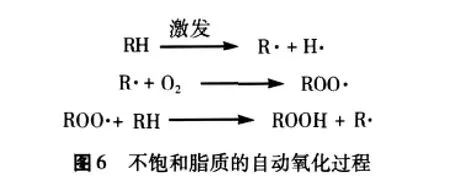
小麦胚芽脂质过氧化物的分解很复杂,伴随着小麦胚芽脂质过氧化物RO-OH的均裂形成烷氧基自由基(RO·),再经过C-C分离,形成醛、酮、醇、酯、呋喃和内酯等分解产物,造成小麦胚芽风味地恶化[50]。
3.3 小麦胚芽的酶促氧化变质
小麦胚芽中脂质酶促氧化酸败是由LOX参加的氧化反应,当小麦胚芽组织被破坏或者受伤时,其中的LA和LOX被激活,脂质的连续酶反应随着脂肪酸的释放而开始(LA),游离脂肪酸中的多不饱和脂肪酸被LOX催化氧化成脂肪酸氢过氧化物;氢过氧化物经过相应的裂解酶催化产生醛和醛酸;异构酶催化生成环氧羟基脂肪酸,后者又水解生成三羟基脂肪酸;过氧化物异构酶催化产生α-乙烯醇脂肪酸和β-乙烯醇脂肪酸;氢过氧化物经环化酶催化生成12 - 氧 - 植物二烯酸[48,51]。
LOX不仅催化小麦胚芽脂质多不饱和脂肪酸(如亚油酸和亚麻酸)中的亚甲基氧化,造成多不饱和脂肪酸的损失,同时LOX也会破坏维生素A,或与一些氨基酸间的相互作用会降低蛋白品质[52],LOX产生的脂肪酸过氧化氢物还能直接与食品中的营养成分如蛋白质和氨基酸结合,这些变化均降低了小麦胚芽的食用和营养价值[53]。
4 结束语
油脂精炼损失是毛油游离脂肪酸含量的2~3倍,随着小麦胚芽毛油中游离脂肪酸的含量增加,可食用小麦胚芽油产率将显著减少。只有游离脂肪酸质量分数低于5%才利于工业化生产,如果超过10%则很不经济。新鲜小麦胚芽通常仅含有1.5% ~2%的游离脂肪酸,但在贮藏数天后,其游离脂肪酸质量分数上升、很快超过5%,因而小麦胚芽必须进行及时稳定化处理。只有掌握了LA和LOX的酶学特性,在探明LA和LOX催化小麦胚芽脂质酸败变质的机制基础上,才能开发可靠的小麦胚芽稳定化技术与装备,为小麦胚芽的进一步利用创造条件。
[1]Tavener R J A,Laidman D L.The induction of lipase activity in the germinating wheat grain[J].Phytochemistry,1972,11(3):989-997
[2]Rothe M.Studien zur Braunung und Aromabildung bei der Maillard - Reaktion[J].Nahrung,1963,7(1):50 -59
[3]Kapranchikov V S,Zherebtsov N A,T N Popova.Purification and Characterization of Lipase from Wheat(Triticum aestivum L.)Germ[J].Applied Biochemistry and Microbiology,2004,40(1):84 -88
[4]Schwimmer S.Source Book of Food Enzymology[M].Westport:The AVI Publishing Co.Inc,1981
[5]Barnes P,Galliard T.Rancidity in cereal products[J].Lipid Technology,1991(3):23 -28
[6]Connor J O,Perry H J,Harwood J L.A Comparison of Lipase Activity in Various Cereal Grains[J].Journal of Cereal Science,1992,16(2):153 -163
[7]Kent N L.Technology of Cereals[M].3rd ed.Oxford:Pergamon Press Ltd,1983
[8]Amado R,Arrigoni E.Nutritive and Functional Properties of Wheat Germ[J].International Food Ingredients,1992(4):30-34
[9]Haki G D,Rakshit S K.Developments in industrially important thermostable enzymes:a review[J].Bioresource Technology,2003,89(1):17 -34
[10]Faber K.Biotransformations in Organic Chemistry[M].5th ed.Berlin:Springer- Verlg,2004
[11]Patel R N.Biocatalysis:Synthesis of Chiral Intermediates for Pharmaceuticals[J].Current Organic Chemistry,2006,10(11):1289-1321
[12]Sharma R,Chisti Y,Banerjee U C.Production,purification,characterization,and applications of lipases[J].Biotechnology Advances,2001,19(8):627 -662
[13]张树政.酶制剂工业:下册[M].北京:科学出版社,1984:655-670
[14]Rantwijk F V,Sheldon R A.Enantioselective acylation of chiral amines catalysed by serine hydrolases[J].Tetrahedron,2004,60(3):501 -519
[15]Alfonso I,Gotor V.Biocatalytic and biomimetic aminolysis reactions:useful tools for selective transformations on polyfunctional substrates[J].Chemlnform,2004,35(30):201 -209
[16]Pleiss J,Fischer M,Peiker M,et al.Lipase Engineering Database:Understanding and exploiting sequence-structurefunction relationships[J].Journal of Molecular Catalysis B:Enzymatic,2000,10(5):491 -508
[17]Tuhkanen A C.Purification and Characterization of Barley Lipase[D].Fargo:North Dakota State University of Agriculture and Applied Science,2000
[18]Sullivan B,Howe M A.Lipase of wheat[J].Journal of the American Chemical Society,1933,55(1):320 -324
[19]O'Connor J,Perry H J,Harwood J L.A comparison of lipase activity in various cereal grains[J].Journal of Cereal Science,1992,16(2):153 -163
[20]Rose D J,Pike O A.A Simple Method to Measure Lipase Activity in Wheat and Wheat Bran as an Estimation of Storage Quality[J].Journal of the American Oil Chemist's Society,2006,83(5):415 -419
[21]Singer T P,Hofstee B H J.Studies on wheat germ lipase.I.Methods of estimation,purification and general properties of the enzyme[J].Archives of Biochemistry and Biophysics,1948,(18):229 -244
[22]Stauffer C E,Glass R L.The glycerol ester hydrolases of wheat germ[J].Cereal Chemistry,1966,43:644 -657
[23]Fadiloglu S,Sylemez Z.Preliminary Kinetic Studies on the Esteratic and Lipolytic Components of a Commercial Wheat Germ Lipase[J].Journal of Agricultural and Food Chemistry,1996,44(10):3015 -3017
[24]Kapranchikov V S,Zherebtsov N A,Popova T N.Purification and Characterization of Lipase from Wheat(Triticum aestivum L.)Germ [J].Applied Biochemistry and Microbiology,2004,40(1):84 -88
[25]Rajeshwara A N,Prakash V.Efect of Denaturant and Cosolvents on the Stability of Wheat Germ Lipase[J].Journal of Agricultural and Food Chemistry,1996,44(3):736 -740
[26]Rao K S,Rajendran S,Rajeshwara A N,et al.Structure Stability of lipase from wheat germ in alkaline pH[J].Journal of Protein Chemistry,1991,10(3):291 -299
[27]Rajeshawara A N,Prakash V.Structure Stability of lipase from wheat germ [J].International Journal of Peptide and Protein Research,1994 ,44(5):435 -440
[28]Korneeva O S,Popova T N,Kapranchikov V S,et al.Identification of Catalytically Active Groups of Wheat(Triticum aestivum)Germ Lipase[J].Applied Biochemistry and Microbiology,2008,44(4):349 - 355
[29]夏小乐,杨博,王永华.小麦胚芽脂肪酶的研究进展[J].现代食品科技,2008,24(10):1068 -1070
[30]Audre E,Hou K W.The presence of a lipoid oxidase in soybean,Glycine soya.Lieb [J].Comptes Rendus de l'Académie des Sciences,1932,194:645 -647
[31]Hildebrand D F.Lipoxygenase[J].Physiologia Plantarum,1989,76(2):249 -253
[32]Sumner R.Lipoid oxidase studies A method for the determination of lipoxidase activity[J].Industrial and Engineering Chemistry-Analytical Edition,1943,15(1):14 -15
[33]Nicolas J,Autran M,Drapron R.Purification and some properties of wheat germ lipoxygenase[J].Journal of the Science of Food and Agriculture,1982,33(4):365 -372
[34]Shiiba K,Negishi Y,Okada K,et al.Purification and characterization of lipoxygenase isozymes from wheat germ[J].Cereal chemistry,1991,68(22):115 - 122
[35]Bhirud P R,Sosulski F W,Sosulski K.Optimizing assay and extraction of lipoxygenase in wheat germ [J].Journal of Food Science,1993,58(5):1090 -1094
[36]Kühn H,Wiesner R,Rathmann J,et al.Formation of ketodienoic fatty acids by the pure pea lipoxygenase-1[J].Eicosanoids,1991b,4(1):9 -14
[37]Shimizu T,Radmark O,Samuelsson B.Enzyme with dual lipoxygenase activities catalyses leukotriene A4 synthesis from arachidonic acid[J].Proceedings of National Academy of Science of the United States of America,1984,81(3):689-693
[38]Gardner H W.Lipid enzymes:lipases,lipoxygenase and“hydroperoxidases”[M]//Simis M G,Karel M.Autooxidation in Food and Biological Systems.New York and London:Ple-num Press.1979:447 -504
[39]R Yamauchi,M Kojima,K Kato.Lipoxygenase - catalyzed oxygenation of monogalactosyl dilinolenoyl glycerol in dipalmitoyl phosphatidylcholine liposomes[J].Agricultural and biological chemistry,1985,49(8):2475 -2477
[40]DeGroot J J M C,Veldink G A,Vliegenthart J F G,et al.Demonstration by EPR spectroscopy of the functional role of iron in soybean lipoxygenase - 1[J].Biochemica Biophysica Acta(BBA)- Enzymology,1975,337(1):71 -79
[41]Hamberg M,Samuelsson B.On the specificity of the oxygenation of unsaturated fatty acids catalyzed by soybean lipoxidase[J].The Journal of Biological Chemistry.1967,242(22):5329-5335
[42]Theorell H,Holman R T,Akeson A.A note on the preparation of crystalline soybean lipoxidase[J].Archives of Biochemistry and Biophysics,1947,14:250 -252
[43]O'Connor T P,O'Brien N M.Significance of lipoxygenase in fruits and vegetables[M].New York:Elsevier Science Publishing Co.Inc,1991:338 -364
[44]Tappel A L,Boyer P D,Lundberg W O.The reaction mechanism of soybean lipoxidase[J].Journal of Biological Chemistry,1952,199:267 -281
[45]Verjee Z H M.Isolation of three acid phosphatases from wheat germ [J].European Journal of Biochemistry,1969,9(3):439-444
[46]Barnett R C,Stafford H A,Conn E E,et al.Phosphogluconic dehydrogenase in higher plants[J].Plant Physiology,1953,28(1):115-122
[47]Takano K.Mechanism of lipid hydrolysis in rice bran[J].Cereal Foods World,1993,38(9):695 -698
[48]Sjovall O,Virtalaine T,Lapvetelainen A.Development of Rancidity in Wheat Germ Analyzed by Headspace Gas Chromatography and Sensory Analysis[J].Agricultural and Food Chemistry,2000,48(8):3522 -3527
[49]Frankel E N.Lipid oxidation:Mechanisms,products and biological significance[J].Journal of American Oil Chemists'Society,1984,61(12):1908 -1917
[50]Frankel E N.Volatile lipid oxidation products[J].Progress in Lipid Research,1982,22(1):1 -33
[51]Royo J,Vancanneyt T G,Perez A G,et al.Characterization of three potato lipoxygenase with distinct enzymatic activities and different organspecific and wound-regulated expression patterns[J].Journal of Biological Chemistry,1996,271(35):21012-21019
[52]Richardson T,Hyslop D B.Enzymes[M]//Fennema O R.Food Chemistry.2nd ed.New York:Marcel Dekker Inc,1985:447
[53]Gardner H W.Lipid hydroperoxide reactivity with proteins and amino acids:a review [J].Journal of Agricultural and Food Chemistry,1979,27(2):220 -229.
Mechanism of Acidification and Deterioration of Wheat Germ during Storage
Xu Bin1Li Bo1Miao Wenjuan1Xu Shanyuan2Dong Ying1
(School of Food and Biological Engineering,Jiangsu University1,Zhenjiang 212013)
(Fada Flour Co.,Ltd.2,Xiajin 253216)
Wheat germ(WG)contains abundant nutriments,such as high quality proteins,polyunsaturated fatty acid and vitamin E,as well as lipoxygenase(LOX),lipases(LA)and ester enzyme.Fresh WG deteriorates within two weeks at room temperature and catalyzed by enzymes and microbes,which greatly limited its utilizing efficiency.This paper expatiated the types of LOX and LA from WG,as well as their structure and catalyzing mechanism.Also,the author analyzed the mechanism of acidification and deterioration of WG during storage from perspectives of catalyze hydroxylation and oxidation of LOX,LA,and contamination of microbes.Deep understanding of wheat germ LOX and LA contributed greatly to technology and equipment devising of WG stabilization.
wheat germ,lipoxygenase,lipases,microbe,mechanism,rancidity
TS210.7
A
1003-0174(2011)09-0123-06
江苏省研究生培养创新工程(CX09B-213Z)
2010-11-29
徐斌,男,1969年出生,副教授,粮油加工副产物的深加工技术研究与装备开发
猜你喜欢
食品与发酵工业(2021年23期)2021-12-17
青年文学家(2021年4期)2021-03-18
农民致富之友(2020年19期)2020-07-18
保健与生活(2020年1期)2020-05-07
家庭百事通·健康一点通(2019年11期)2019-09-26
中国洗涤用品工业(2019年4期)2019-05-11
肉类研究(2018年5期)2018-09-26
中国粮油学报(2015年5期)2015-02-06
绿色科技(2014年1期)2014-11-28
饮食科学(2014年8期)2014-08-22

- 中国粮油学报的其它文章
- 二维相关近红外光谱快速鉴别食用植物油种类