
白三叶种质资源形态变异与地理起源的关系
2012-12-24 00:53张婧源
草业科学 2012年11期
李 州,彭 燕,张婧源,马 啸
(四川农业大学动物科技学院,四川 雅安625014)
白三叶(Trifoliumrepens)为豆科(Leguminosae)三叶草属中具有重要农艺价值的多年生牧草,原产于欧洲、北非和亚洲西部,在温带及亚热带地区均有分布,是世界各地栽培利用最为广泛的豆科牧草之一,尤以新西兰、澳大利亚、欧洲西北部、北美东部等地区栽培面积最大,在我国东北、西北、西南及长江中下游等地均有栽培。白三叶生长适应性强,品质优良,适宜与多种禾草混播种植,刈割和耐牧特性好,在草地畜牧业生产中占据重要地位。近年来,由于白三叶的匍匐生长特性及突出的生物固氮能力,亦被广泛应用于园林绿化、水土保持、培肥地力等,被农业部列为向全国推广果园生草覆盖技术的首选草种,在国内环境和生态建设中发挥着重要作用。
白三叶为天然异源四倍体植物,具自交不亲和性。因此,在遗传组成上具有高度杂和的特性,无论是种群内还是种群间皆具有丰富的遗传变异,其形态特征可塑性较大[1]。此外,白三叶广泛分布于世界各地,适应生境具有多元化特点[2]。种质资源收集与遗传多样性研究是品种改良和育种的基础,因而,白三叶种质资源收集及包括形态多样性在内的农艺性状和遗传多样性一直受到国内外相关研究者的重视。为了改良新西兰白三叶品种特性,Widdup等[3]从美国收集了98个白三叶居群,并初步评价其农艺性状。Dzene等[4]从世界各地收集白三叶材料,以筛选优良白三叶种质改良本国现有品种。此外,美国、澳大利亚及欧洲各国都较系统地收集和研究了白三叶种质资源[5-9],而我国对白三叶种质资源的收集及其遗传多样性报道相对较少。安晓珂[10]研究了新疆野生白三叶与三叶草属其他种的遗传多样性及遗传关系,何俊[11]初步评价了从国内外收集的62份白三叶种质资源的遗传多样性,吴海涛[12]评价了从俄罗斯引进的18份野生白三叶种质资源的生长适应性、饲用性和景观坪用性,Xian等[13]对采自国内中部、西南和西北部的14个白三叶居群进行了形态特征和分子遗传多样性研究,切彩等[14]分析了新疆11份野生白三叶的形态变异,张婧源等[15]应用SRAP分子标记研究了原产于世界各地的41份白三叶种质的遗传变异。本研究以收集到的源自29个国家的68份野生白三叶种质为试材,研究其主要形态特征变异,并分析不同起源地白三叶形态变异分化趋势,以期为筛选出不同用途优良的白三叶种质及为白三叶核心资源库的建立提供基础资料。
1 材料与方法
1.1 供试材料 供试材料源自29个国家,共计68份,材料编号及来源见表1。
1.2 观测指标与方法 2008年8月取50粒白三叶种子于智能光照培养箱中催芽,幼苗在沙盘中培育1个月左右,2008年10月上旬将幼苗移栽至田间试验地,试验地位于四川洪雅阳平种牛场试验区,小区面积1m×1m,每小区移栽10株,株间距10 cm,重复3次。于2009年白三叶开花期进行观测,每小区观测10株。株高(cm)为地面至植株顶部的自然高度;生殖枝高(cm)为地面至生殖枝顶部的自然高度;用数码照相机拍下距匍匐茎末端倒数第3叶的中间小叶,并用Photoshop软件分析小叶面积(cm2);观测每小区开花的花序数,开花后,连续观测4周,取平均值得到单位面积花序数;每小区随机取5个完全开放的花序,观测其小花数,连续观测4周,取平均值得每花序小花数。

表1 供试白三叶种质及来源Table 1 Serial number and origin of Trifolium repens accessions

续表1
1.3 数据分析和统计方法 采用Microsoft office Excel 2003对数据进行处理;用SPSS 11.0软件按欧氏距离对数据进行聚类分析;用NTSYS-pc program计算遗传相似性系数矩阵和地理距离矩阵,用Mantel test检验两者的相关性。
2 结果与分析
2.1 供试白三叶种质形态变异 通过对68份白三叶种质资源的5个形态特征进行基本统计分析,结果表明(表2),不同材料之间存在很大差异,表现出明显的形态多样性。叶面积的变异最大,变异系数高达69.48%,其次是单位面积花序数和株高,变异系数分别为50.24%和44.30%,而生殖枝高度和每花序小花数变异较小,变异系数分别为19.90%和18.81%。
供试材料平均株高为12.29cm,最高为23.27 cm,最低仅为1.34cm,变异较大。生殖枝和株高相比,表现为3种情况,一是明显高于株丛,二是比株丛稍高,三是低于株丛。生殖枝平均高度为18.04 cm,最高为27.36cm,最短为9.50cm,变异较小。叶片大小变异最大,叶面积最大为18.44cm2,最小为0.23cm2,平均值为6.98cm2。68份白三叶开花能力差异很大,单位面积花序数平均为26.95个·m-2,最多可达50.17个·m-2,最少仅为2.67个·m-2。每花序小花数的变异最小,平均值为65.75个,最多为87.64个,最少为36.00个。
2.2 形态特征与地理距离的相关性分析Mantel检验表明,白三叶形态变异受地理环境的影响(表3),综合指标(包括株高、生殖枝高度、叶面积、单位面积花序数和每花序小花数5个指标)、株高、单位面积花序数、每花序小花数特征变化存在空间相关性,且综合指标、单位面积花序数、每花序小花数达到极显著水平(P<0.01),株高达到显著水平(P<0.05),而生殖枝高度、叶面积相关性不显著。
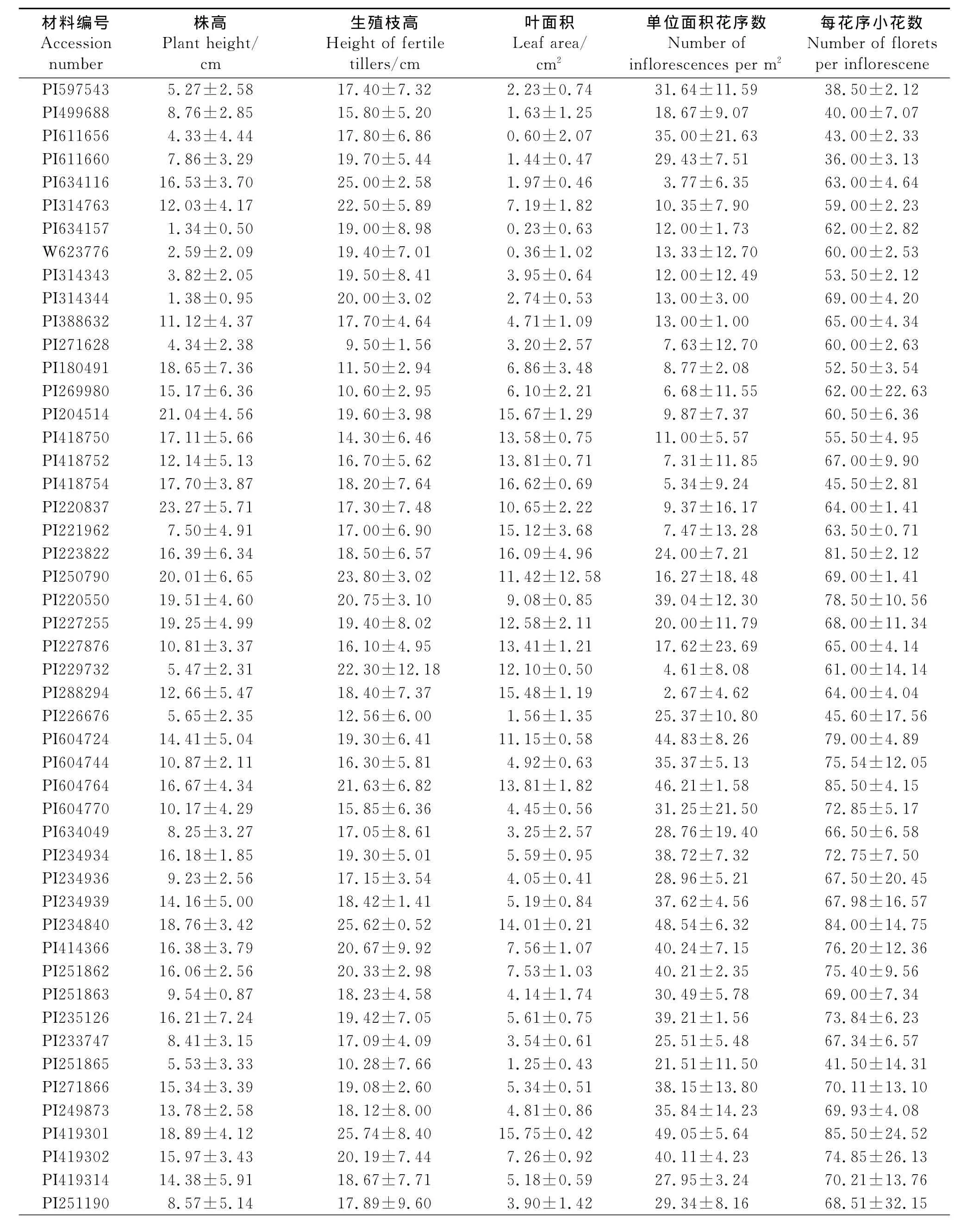
表2 白三叶种质形态特征及变异Table 2 Morphological characteristics and variations in Trifolium repens accession for test

续表2

表3 白三叶形态特征与地理距离相关性的Mantel检验Table 3 Mantel test between morphological characters and geographical distance of Trifolium repens
2.3 聚类分析 对供试材料形态特征按欧氏距离进行聚类分析的结果见图1,不同材料可明显地划分为4个类群,不同类群形态特征值见表4。第1类群包括33份材料,除1份亚洲材料和3份非洲材料外,其余29份全部来自欧洲,植株高度和生殖枝高度分别为(13.37±3.49)cm、(18.38±1.91)cm,叶面积为(6.24±2.74)cm2,单位面积花序数和每花序小花数较多,分别为(36.33±5.90)个·m-2和(72.98±4.96)个,表现为植株较高,中到小叶型叶片,有性繁殖能力较强;第2类群包括15份材料,其中亚洲10份(东亚4份,中亚4份,南亚、西亚各1份),欧洲(南欧)5份,形态指标为植株和生殖枝最低矮,分别为(4.92±2.08)cm 和(15.52±3.80)cm,叶面积最小,为(1.81±1.09)cm2,单位面积花序数较少,每花序小花数最少,分别为(20.51±8.26)个·m-2、(49.29±9.98)个,此类白三叶表现为植株低矮、小叶型、有性繁殖能力较弱;第3类群包括17份材料,全部来自亚洲,其中,西亚12份,中亚3份,南亚2份,植株和生殖枝高度分别为(15.11±4.87)cm、(18.17±3.90)cm,叶片较大,面积为(11.37±4.44)cm2,单位面积花序数最少,而每花序小花数较少,分别为(10.48±5.97)个·m-2
和(62.71±7.62)个,表现为植株较高,叶片中到大叶型、有性繁殖能力较弱;第4类的3份材料,分别来自欧洲的德国、南欧的希腊和斯洛文尼亚,各形态指标均为最大值,株高、生殖枝高、叶面积、单位面积花序数和每花序小花数分别为(19.11±0.50)cm、(26.24±0.97)cm、(16.07±2.23)cm2、(49.25±0.83)个·m-2、(85.71±1.83)个,表现为植株高大、大叶型、有性繁殖能力强。

图1 白三叶种质资源形态特征聚类图Fig.1 Morphological clustering analysis of Trifolium repens accessions

表4 不同类群白三叶形态特征Table 4 Morphological characters of different groups for Trifolium repens accessions
3 讨论与结论
3.1 白三叶分布生境特点 白三叶起源于欧洲地中海地区,由于动物迁移等,传播至欧洲和西亚各地。目前,在年降水量大于750mm的温带地区都有分布[16]。本研究中,供试白三叶主要分布于东半球和北半球的中低纬度区域,尤以大于30°的中纬度和海拔1 000~2 000m的区域最为丰富。相关分析亦显示,白三叶分布地的纬度与经度、纬度与海拔呈显著负相关,而经度与海拔呈显著正相关。由此可见,白三叶分布的地理环境与白三叶生态习性相吻合,即白三叶喜冷凉-温暖的气候条件。
3.2 白三叶形态变异与生长特性的关系 叶片大小是区分白三叶类群的重要特征。按叶片大小白三叶可分为小叶、中叶、大叶和拉丁诺4个类型[17]。本研究显示,在白三叶形态变异中,叶片大小的变异最大。聚类形成的4个类群,叶片呈从小到大的梯度变化。除叶片外,白三叶株高和单位花序数的变异亦较大,这与Jahufer等[6]对澳大利亚的439份白三叶种质形态特征的研究结果相一致。株高比其他特征的变异相对较大,表明白三叶生长高度和开花特性的可塑性较大,易受环境的影响而发生变化。在本研究中,生殖枝高度和每花序小花数变异相对较小,这可能与植物繁殖特性主要受遗传制约而相对保守有关。在研究的形态特征中,除叶面积与单位面积花序数外,其余形态特征两两之间呈显著正相关(P<0.05),即大叶型的白三叶植株高大,开花时生殖枝亦较高,每花序小花数多,有性生殖能力较强,而小叶型的白三叶则植株低矮,有性生殖能力较弱。Finne等[9]也有类似报道,即白三叶叶丛高度与小叶长度和干物质产量呈两两正相关,种子产量与叶丛高度呈正相关。
3.3 白三叶形态变异与地理起源关系 白三叶形态多样性与分布生境的多元化有关18。本研究的Mantel相关分析表明,白三叶形态变化与地理环境之间存在显著相关性。国外有关白三叶形态与分布生境相互关系的报道较多。沿地中海盆地,白三叶叶片的大小梯度与温度和水分有关[18]。小叶型的主要见于北部地区,如英国,叶片较大的拉丁诺主要分布于意大利北部,而叶片更大的白三叶则出现在欧洲南部和地中海。就纬度和海拔而言,小叶白三叶主要分布于高纬度和高海拔地区,大叶白三叶则分布于南部和低海拔地区[17-18]。并与 Widdup等[3]报道的美国白三叶种质形态变异相一致,即白三叶叶片随纬度和海拔升高而变小,低纬度和低海拔地区,叶片最大。本研究显示,单位面积花序数、每花序小花数与地理距离呈极显著相关性,株高与地理距离呈显著相关性。
进一步的聚类结果显示,源于欧洲和亚洲的大部分材料,分别聚为不同的中-小叶型和中-大叶型两个类群,而其他两个分别为3份源自欧洲的大叶型类群和5份源自欧洲的与10份源自亚洲的小叶型类群。即原产地不同的多种材料聚为不同的类群,表明白三叶形态变异与地理起源存在较密切的相关性,而少数分别起源于欧洲和亚洲的不同白三叶种质亦聚为相同的一类,从形态上看,属于一个叶片最小的类群,且3份源自欧洲的叶片最大的材料单独聚为一类。表明不同起源的白三叶亦出现相似的遗传分化,原产欧洲的白三叶形态朝不同方向的进化趋势明显,可能与欧洲为白三叶起源中心,遗传多样性丰富有关。Widdup等[3]报道,由于各种选择压力,如夏季的高温、病毒病害、虫害以及基因渐渗,北美白三叶种质资源叶片具有朝小叶和中叶进化的趋势。Gustine等[19]研究亦显示,白三叶叶片大小变异很大,温和的选择压力可能使他们朝不同的方向发展。
近年来,白三叶在我国作为牧草、生态草、园林植物等栽培应用增长迅速。然而,我国生产中应用的白三叶品种主要从国外进口,多数表现为对干旱、高温的适应性差,病虫害严重,因而产量低、持久性差,阻碍了白三叶生产潜力的发挥。大量研究表明,不同生境的白三叶种质具丰富的形态变异以及多样化的生长和适应环境的特性[20-27],且不同地理起源的白三叶种质保持着丰富的分子遗传变异[28-32]。由此可见,在更广泛的范围内,不同生境条件下收集不同生态型的白三叶种质尤其是本土的种质,以选育和改良优良白三叶品种,满足不断增长的多元化生产需求具有重要意义。
[1]Voisey C R,White D W R,Wigley P J,etal.Release of transgenic white clover plants expressingBacillus thuringiensisgenes:An ecological perspective[J].Biocontrol Science and Technology,1994,4:475-481.
[2]Caradus J R,Forde M B,Wewala S,etal.Description and classification of a white clover(TrifoliumrepensL.)germplasm collection from southwest Europe[J].New Zealand Journal of Agricultural Research,1990,33:367-375.
[3]Widdup K H,Caradus J R,Green J,etal.White clover ecotype germplasm from the USA for development of New Zealand and overseas cultivars Grassland[J].Research and Practice Series,1996,6:149-154.
[4]Dzene I,Jansons A,Jansone B,etal.The study of variability of agro-morphological characteristics of white clover accessions from other countries in Latvian climate conditions[J].Agronomy Research(Special Issue III),2010,8:537-544.
[5]Gustine D L,Huff D R.Genetic variation within and among white clover populations from managed permanent pastures of the northeastern U.S[J].Crop Science,1999,39:524-530.
[6]Jahufer M Z Z,Cooper M,Harch B D.Pattern analysis of the diversity of morphological plant attributes and herbage yield in a world collection of white clover(TrifoliumrepensL.)germplasm characterised in a summer moisture stress environment of Australia[J].Genetic Resources and Crop Evolution,1997,44:289-300.
[7]Paolo A,Rosemary P C,Flavio F,etal.Variation in cold tolerance and spring growth among Italian white clover populations[J].Euphytica,2001,122:407-416.
[8]Negri V,Veronesi F.Possible role of natural populations of white clover for reclamation purposes in subhumid and humid Mediterranean environments[J].Agronomie,1987,7:703-708.
[9]Finne M A,Rognli O A,Schjelderup I.Genetic variation in a Norwegian germplasm collection of white clover(TrifoliumrepensL.)[J].Euphytica,2000,112:33-44.
[10]安晓珂.三叶草属三种植物遗传多样性的RAPD分析[D].北京:中国农业科学院草原研究所,2008.
[11]何俊.62份白三叶种质资源遗传多样性初步研究与评价[D].贵州:贵州农业大学,2008.
[12]吴海涛.俄罗斯引进野生白三叶种质资源的价值评价[D].江苏:扬州大学,2009.
[13]Xian Z,Ying J Z,Rong Y,etal.Genetic variation of white clover(TrifoliumrepensL.)collections from China detected by morphological traits,RAPD and SSR African[J].Journal of Biotechnology,2010,9(21):3032-3041.
[14]切彩,李卫军,朱昊.11份白三叶材料的形态特征及生长表现[J].新疆农业科学,2010,47(1):135-140.
[15]张婧源,彭燕,罗燕,等.不同产地白三叶种质遗传多样性 的 SRAP 分 析 [J].草 业 学 报,2010,19(5):130-138.
[16]Lane L A,Ayres J F,Lovett J V.A review of the introduction and use of white clover(TrifoliumrepensL.)in Australia-significance for breeding objects[J].Australia Journal of Experimental Agriculture,1997,37:831-839.
[17]Caradus J R,MacKay A C,Woodfield D R,etal.Classification of a world collection of white clover cultivars[J].Euphytica,1989,42:183-196.
[18]Davies W E,Young N R.The characteristica of European,Mediterranean and other populations of white clover(TrifoliumrepensL.)[J].Euphytica,1967,16:330-340.
[19]Gustine D L,Voigt E C,Brummer P W,etal.Genetic variation of RAPD markers for North American white clover collections and cultivars[J].Crop Science,2002,42(2):343-347.
[20]Campbell B D,Caradus J R,Hunt C L.Temperature responses and nuclear DNA amounts of seven white clover populations which differ in early spring growth rates[J].New Zealand Journal of Agricultural Research,1999,42:9-17.
[21]Fraser J.Characteristics of naturalized populations of white clover(Trifoliumrepens)in Atlantic Canada[J].Canadian Journal of Botany,1989,67:2297-2301.
[22]Caradus J R,Christie B R.Winter hardiness and artificial frost tolerance of white clover ecotypes and selected breeding lines[J].Canadian Journal of Plant Science,1998,78:251-255.
[23]Van D B J,Black I K,Cousins G R,etal.Enhanced drought tolerance in white clover[J].Proceedings of the New Zealand Gmssland Association,1993,55:97-101.
[24]Jahufer M Z Z,Cooper M,Lane L A.Variation among low rainfall white clover(TrifoliumrepensL.)accessions for morphological attributes and herbage yield[J].Australian Journal of Experimental Agriculture,1995,35:1109-1116.
[25]Jahufer M Z Z,Cooper M,Brien L A.Genotypic variation for stolon and other morphological attributes of white clover(TrifoliumrepensL.)populations and their influence on herbage yield in the summer rainfall region of New South Wales[J].Australian Journal of Agricultural Research,1994,45(3):703-720.
[26]Lane L A,Ayres J F,Lovett J V,etal.Morphological characteristics and agronomic merit of white clover(TrifoliumrepensL.)poulations collected from northern New South Wales[J].Australian Journal of Agricultural Research,2000,51:985-997.
[27]Caradus J R,Hay R J M,Woodfield D R.The positioning of white clover cultivars in New Zealand[J].Grassland Research and Practice Series,1996,6:45-50.
[28]Jahufer M Z Z,Barrett B A,Griffith G S,etal.DNA fingerprinting and genetic relationships among white clover cultivars[J].Proceedings of New Zealand Grassland Association,2003,65:163-169.
[29]Kölliker R,Jones E S,Jahufer M Z Z,etal.Bulked AFLP analysis for the assessment of genetic diversity in white clover(TrifoliumrepensL.)[J].Euphytica,2001,121:305-315.
[30]Kölliker R,Jones E S,Drayton M C,etal.Development and characterisation of simple sequence repeat(SSR)markers for white clover(TrifoliumrepensL.)[J].Theoretical and Applied Genetics,2001,102:416-424.
[31]George J M P,Dobrowolski E Z,Jong N O,etal.Assessment of genetic diversity in cultivars of white clover(TrifoliumrepenseL.)detected by SSR polymorphisms[J].Genome,2006,49:919-930.
[32]Fernanda B,Miguel D A,Maria T S W.Molecular characterization of the USDA white clover(TrifoliumrepensL.)core collection by RAPD markers[J].Genetic Resources and Crop Evolution,2006,53:1081-1087.
猜你喜欢
现代园艺(2023年3期)2023-02-01
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14
江苏农业科学(2019年5期)2019-09-02
植物资源与环境学报(2019年1期)2019-04-02
现代园艺(2017年22期)2018-01-19
南风(2017年34期)2018-01-16
广东农业科学(2017年5期)2017-08-29
科学种养(2017年6期)2017-06-13
科学种养(2017年5期)2017-05-17
应用海洋学学报(2014年1期)2014-11-22
