
长白山笃斯越橘菌根形态结构及内生菌的分布特征
2013-02-20 09:21苗迎秋王贺新李根柱张自川
东北农业大学学报 2013年1期
苗迎秋,王贺新,李根柱,张自川,汤 莉
(1.大连大学医学院,辽宁 大连 116622;2.大连大学现代农业研究院,辽宁 大连 116622)
笃斯越橘(Vaccinium uliginosum Linn.)为杜鹃花科越橘属植物,落叶灌木,果实为蓝紫色小浆果,含有较高的花青素类(Anthocyanins)物质,具有较强的抗氧化活性和较高的保健价值[1-2]。在我国笃斯越橘主要分布在大兴安岭北部及小兴安岭和长白山地区。在长白山地区,野生笃斯越橘主要生长在海拔700~2 000 m之间的针阔混交林、林沟湿草地、沼泽等地带,呈块状或片状分布[3]。
菌根是植物根系和真菌所建立的共同体,是植物与微生物所形成的互惠共同体,几乎普遍存在于所有植物中[4]。一般认为,越橘属植物主要由欧石楠菌根真菌(Ericoid mycorrhiza,ERM)侵染,有时也有丛枝菌根真菌(Arbuscular mycorrhiza,AM)和暗隔内生菌根真菌(Dark septate endophytes,DSE)侵染[4-5]。但是,目前国内外相关研究报道仅涉及对笃斯越橘的资源调查、果实营养价值、树木生理和开发利用等方面,而有关其菌根方面研究鲜有报道。本研究组针对长白山地区笃斯越橘菌根的形态结构和菌根真菌的分布特征进行初步观察,为笃斯越橘菌根生态学研究及资源合理开发和管理提供理论依据。
1 研究地点与方法
1.1 取样地点概况
研究地点为长白山自然保护区东坡和长白山麓的吉林哈泥省级自然保护区。长白山自然保护区海拔1 300 m以下的笃斯越橘部分生长在泥炭藓沼泽草甸子里,部分生长在苔藓层较厚的落叶松林下。调查样地中心点位于42°02′37″N,128°21′36″E,距圆池入口西侧约10 km、边防公路北约2 km处。群落组成为8落2桦,林分为异龄复层林,笃斯越橘主要着生于透光度较好的林下或林间空地,呈团状或片状分布,郁闭度>6.0以上的林内很少见到团片状的笃斯越橘灌丛,偶遇少量笃斯越橘却株型细弱,生长不良。土壤垂直结构自上而下依次明显分为4层,最上面第1层为活苔藓层,相当于凋落物层(O层)的L层,其中混有大量新鲜的落叶松凋落物,分解较少;第2层是死苔藓层,相当于凋落物层(O层)的F层,多为半分解的死苔藓和落叶松凋落物;第3层主要由呈泥炭化的腐殖质层(A1层)及其下部少量较薄的淀积层(A2层)组成,此处合并称为泥炭层(A层),与下层过渡明显;第4层为火山灰层(C层),由黄白色至棕黄色的火山灰及其颗粒组成。本项目的另一取样地点为吉林哈泥省级自然保护区,位于吉林省长白山西侧的龙岗山脉中部。该区海拔557~1 212 m,属温带湿润、半湿润季风气候。调查样地中心点位于42°13′27″N,126°29′35″E。两处采样地点均为笃斯越橘-落叶松林型,乔木多为长白落叶松(Larix olgensis Henry),伴有白桦(Betula platyphylla)等阔叶树,下层大灌木有柴桦(Betula fruticosa Pall),小灌木主要有细叶杜香(Ledum palustre L.)、越橘(Vaccinium vitis-idaea L.)、蓝靛果忍冬(Lonicera edulis Turcz.)和山刺玫(Rosa davurica Pall)等。草本植物大部分是苔草(Carex spp.)和小叶章(Calamagrostis angustifolia Kom.Fern)等。各取样点的笃斯越橘根系主要生长在枯死苔藓层和疏松的枯枝落叶层内。
1.2 方法
2010年7月于上述区域随机对笃斯越橘根部进行取样,取样时轻轻去掉根系表面附着的苔藓枯死残留物,确保根尖完整,然后取根段在流水下轻轻冲洗干净,选取不同粗细的营养根,切成1 cm长小段,用软化酸处理后固定于FAA固定液中。用下列两种方法进行观察。一是石蜡切片法,即将样根带回实验室后经脱水、透明、采用常规石蜡切片法包埋、切片,并用番红固绿对染和加拿大树胶封片,在光学显微镜下观察。二是对于纤细的毛根采用压片方法直接观察,即将FAA固定的根段经20%KOH透明处理、再经2%HCl酸化处理后,用0.02%棉蓝染液染色,直接压片后用光学显微镜观察。
2 结果与分析
2.1 笃斯越橘根形态结构
调查显示,笃斯越橘的根系实际上是由根和根态茎共同组成,而根系的骨干部分还是以茎为主,茎代行骨干根的功能。真正的根并不发达,绝大部分为须根,稀疏地着生在根态茎上,有少量发生粗大一些的根,呈不规则的小弯曲状,与既有研究一致[5]。成熟毛根非常纤细(直径为150 μm以下),次生结构不够典型。一般周皮由3~4层木栓细胞组成,其中最外一层呈凋落状;维管形成层、次生韧皮部和次生木质部均不明显,各占1~2层细胞;初生木质部占有中心很小部分空间(见图1A);表面没有根毛,包裹着1层或疏松或紧密的菌丝层(见图1B)。直径为200 μm以上的笃斯越橘根毛根的次生结构较为明显,周皮细胞5~6层,排列紧密,胞间隙小,壁木质化,最外围1~2层呈凋落状,经番红固绿染色后呈红色;维管形成层和次生韧皮部不明显,各占1~2层细胞,而次生木质部占有大部分空间(见图1C)。切片结果还显示笃斯越橘初生构造中侧根由中柱鞘细胞产生,对着木质部放射角的细胞分生能力强,发生分裂,向外突起,形成侧根。在侧根形成过程中,分化出维管组织,并且与主根维管组织连接在一起(见图1D)。
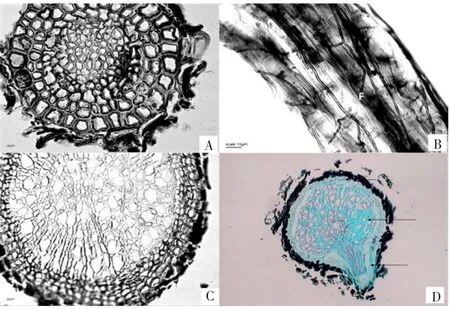
图1 笃斯越橘菌根显微形态结构Fig.1 Microscopic morphology structure of mycorrhiza of Vaccinium uliginosum
2.2 笃斯越橘内生菌根真菌的分布
从笃斯越橘根的连续石蜡切片中可以看到,在已经开始次生生长的毛根内,内生菌根真菌主要侵染根的表皮细胞和周皮细胞(见图2A、B),含有菌丝体的细胞壁有150 μm加厚迹象,形成典型的ERM菌丝圈。还发现在初生构造的毛根内,有菌根可以侵染到中柱情况,但没有改变根的基本解剖结构(见图2C、D)。
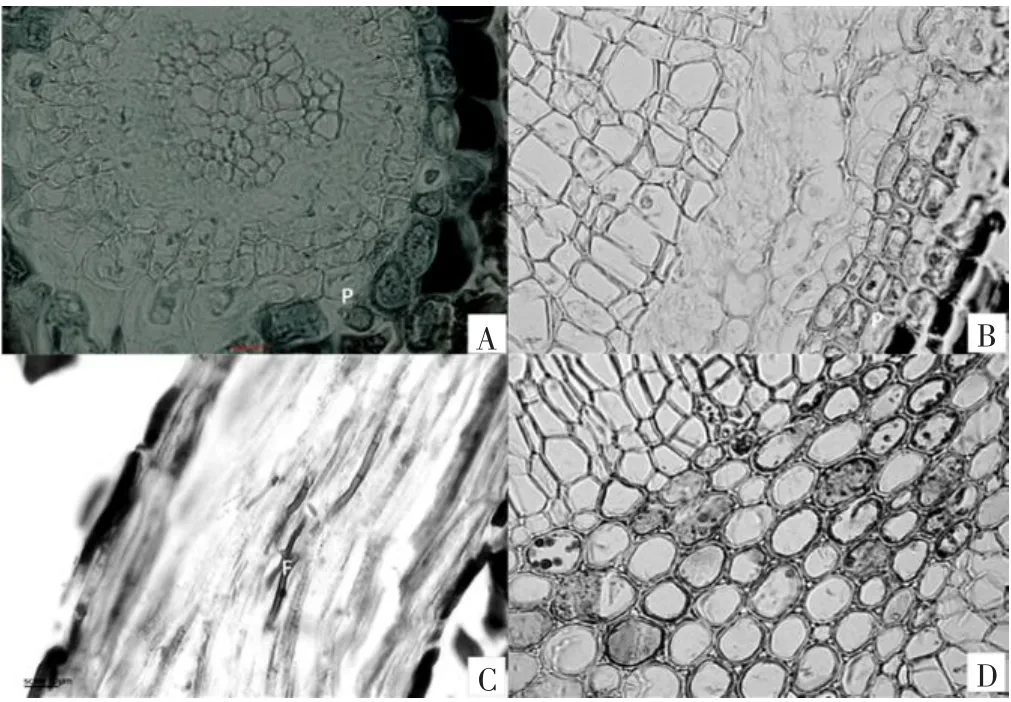
图2 笃斯越橘内生菌根真菌的分布Fig.2 Distribution of endotrophicmycorrhiza fungi of Vaccinium uliginosum
2.3 笃斯越橘内生菌根真菌的菌丝形态
棉蓝染色压片光镜观察结果显示,根据菌丝及菌丝圈的形态判断,至少有三种内生真菌侵染笃斯越橘根部细胞。
一种是具有明显的横隔膜真菌菌丝能穿破笃斯越橘根表层细胞壁,并在其中定殖,不断繁殖,并侵染皮层或周皮的下一层细胞,形成复杂浓密的菌丝群,即典型的欧石楠类菌丝复合体(见图3A),构成笃斯越橘的菌根菌的主体;
另一种是由具有明显的横隔膜、呈棕褐色且较粗的真菌菌丝形成的较疏松的暗隔内生菌菌丝复合体(见图3B),在调查的笃斯越橘根系中普遍存在;
第三种菌丝是没有明显的横隔膜,但具有典型的二分叉结构,并在细胞内形成泡囊(见图3C)和丛枝(见图3D)的泡囊-丛枝菌丝复合体,在调查根系中数量不多但也经常出现。

图3 菌根细胞内菌丝复合体形态Fig.3 Morphology of mycelium complex in mycorrhiza cell
3 讨论与结论
3.1 笃斯越橘根的分布及菌根形态特征
笃斯越橘是东北森林群落的重要组成成分之一,具有重要的生态和经济价值,相关研究很少。研究组在长白山实地观察结果显示,笃斯越橘根系由根状茎、行茎及其上面着生的细根和须根构成,与分布于北美的同属矮丛越橘(Vaccinium angustifolium Aiton)相似[6],但是须根数量不多,明显少于矮丛越橘。笃斯越橘根状茎和须根只生长在有苔藓枯死体和落叶松等凋落物形成的松软基质内,几乎没有矿质土壤成分[7]。须根在分枝前直径为50~80 μm,没有根毛。研究指出菌根对杜鹃花类植物的营养吸收以及增强对逆境因子的抗性等方面具有重要作用[8],笃斯越橘的菌根是否在某种程度上起到根毛的作用有待于深入研究。
笃斯越橘根的形态特征表明,在次生构造中具有明显的周皮和次生木质部,周皮细胞呈木栓化,胞内空隙较大。维管形成层和次生韧皮部均不明显,这可能是由于笃斯越橘的毛根过于纤细,不可能形成太多的细胞层次有关(见图1)。另外,根系横剖面图显示根表皮细胞脱落、变空,应该是根系次生生长产生的木栓保护层及其凋亡外层细胞。镜下可见笃斯越橘的根周皮细胞壁呈现木质化加厚,番红固绿双重染色后呈红色,此厚壁细胞的形成一般被认为是植物体的一种防御反应,而且切片结果显示直径为50 μm以下的毛根表皮细胞壁也有呈现木质化加厚,有可能是为了适应长白山特殊的土壤环境而形成的一种形态变化。
越橘属植物的菌根主要由ERM菌根菌构成[4-5,8],通常在表皮细胞内具有典型的ERM菌丝圈[9]。从图1B可以看出,在毛根的表面具有一层疏松的菌丝层,这与大多数杜鹃科植物相同,从图2A、B,图3A可以看出,经番红固绿双重染色后,细胞内含有明显深色团块状结构的就是受到菌丝侵染的细胞,整个皮层细胞层受到侵染的细胞随机分布,一些根系已经在表皮和周皮内初步呈现环状菌丝圈的特征。一般认为,表层细胞是真菌与植物体相互作用的活跃部位,当菌丝侵染到距内皮层1~2层细胞时便停止侵染,也就是说菌丝一般不能侵染内皮层及中柱,从而使根仍能保持生活力。但本研究采集的菌根中表皮细胞、皮层细胞和中柱都有不同程度内生真菌的侵染(见图2B、C),但没有改变根的基本解剖结构,这在其他杜鹃花科植物中未见报道。这种现象是偶然的还是普遍存在的现象,对笃斯越橘有无影响等,有待于我们今后详细研究确认。
3.2 笃斯越橘菌根菌的种类
调查发现,长白山野生笃斯越橘菌根菌以欧石楠类菌根菌ERM为主,也有暗隔内生菌根真菌DSE和泡囊-丛枝菌根真菌AM感染的现象(见图3)。有关杜鹃花科植物菌根研究偏注重于ERM菌根菌,有报道在兴安杜鹃菌根形态特征中发现除ERM外有少量AM侵染特征[10],也有利用暗隔内生菌根真菌(DSE)接种越橘类植物促进其生长报道[11]。说明3种菌根菌在杜鹃花科及越橘属植物根系内存在。
[1]Lui J,Zhang W,Jing H,et al.Bog bilberry(Vaccinium uliginosum L.)extract reduces cultured Hep-G2,Caco-2 and 3T3-L1 cell viability,affects cell cycle progression and has variable effects on membrane permeability[J].Journal of Food Science,2010,75(3):103-107.
[2]张晓松,孙艳梅,胡振生,等.都柿果酒在酿制过程中总酚和花色苷含量和抗氧化活性[J].东北农业大学学报,2010,41(3):120-124.
[3]郝瑞.长白山笃斯越橘的调查研究[J].园艺学报,1979,6(2):87-93.
[4]刘润进,陈应龙.菌根学[M].北京:科学出版社,2007:1-124.
[5]Monika A G,Sarah H,Hugues B M.Community structure of ericoid mycorrhizas and root-associated fungi of Vaccinium membranaceum across an elevation gradient in the Canadian Rocky Mountains[J].Fungal Ecology,2012,5(1):36-45.
[6]顾姻,贺善安.蓝浆果与蔓越橘[M].北京:中国农业出版社,2001:17-19.
[7]李根柱,王贺新,陈英敏,等.长白山落叶松林下笃斯越橘群落生物量的空间分布[J].生态学杂志,2012,31(6):1404-1410.
[8]张春英,戴思兰.杜鹃花类菌根研究进展[J].北京林业大学学报,2008,30(3):113-119.
[9]Massicotte H B,Melville L H,Peterson R L.Structural characteristics of root-fungal interactions for five ericaceous species in eastern Canada[J].Canadian Journal of Botany,2005,83:1057-1064.
[10]贾锐,杨秀丽,闫伟,等.兴安杜鹃菌根形态特征和土壤理化性质的关系研究[J].内蒙古农业大学学报:自然科学版,2011,32(3):63-66.
[11]Vano I,Sakamoto K,Inubushi K.Selection of dark septate endophytes from Ericaceae plants to enhance blueberry(Vaccinium corymbosum L.)seedling growth[C].Abstracts of 7th International Symposium on Integrated Field Science,2009:15.
猜你喜欢
食品与药品(2021年6期)2021-12-21
学生天地(2020年34期)2020-06-09
园林科技(2020年2期)2020-01-18
辽金历史与考古(2019年0期)2020-01-06
中国工程咨询(2015年8期)2015-02-16
中国酿造(2014年12期)2014-04-12
安阳工学院学报(2014年5期)2014-03-21
安徽农业科学(2014年35期)2014-03-20
中国果业信息(2014年2期)2014-01-23
植物营养与肥料学报(2011年6期)2011-10-24
