
葡萄WRKY转录因子家族全基因组分析
2013-04-03 09:16李成慧
经济林研究 2013年4期
李成慧,蔡 斌
(1.苏州农业职业技术学院,江苏 苏州 215008;2.南京农业大学 园艺学院,江苏 南京 210095)
转录因子(transcription factors)在植物的生长发育及其对外界环境的反应中起着重要的调控作用。转录因子通过与基因启动子区域特异性结合,激活或抑制下游基因的转录,在信号传导和诱导下游相关功能基因表达中起着关键作用[1-2]。大部分的转录因子是以多基因家族的形式存在的。WRKY基因家族是植物中的一类重要转录因子家族。WRKY转录因子是锌指蛋白家族的一个亚家族,其N端包含一个由50个氨基酸组成的高度保守的结构域。WRKY转录因子结构域中的N端因有高度保守的7个氨基酸特征序列WRKYGQK而得名,其C末端为一个锌指结构。WRKY基因家族主要存在于植物中,在植物的生长和发育、环境信号刺激及其对病原物的防御反应过程中发挥着非常重要的作用(如损伤反应[3]、种子发育[4]、叶片衰老[5]、调控植物防御反应相关基因[6]等方面的作用)。WRKY基因还参与植物的抗非生物胁迫(如干旱[7-8]、高盐[9]、高温[10]等)反应。
葡萄在世界果树生产中占有重要的位置,具有很高的经济价值[11]。葡萄为完成全基因组测序的第一种水果作物和第四种开花植物[12]。有关葡萄WRKY转录因子的研究起步较晚,有关葡萄抗生物和非生物胁迫反应方面的研究有个别报道[13]。目前对葡萄WRKY转录因子的系统分析和功能研究还未见报道,因此,文中以12×葡萄基因组蛋白序列为数据来源,在全基因组水平上识别和分析WRKY基因家族,对其进行了系谱发生、共线性和表达谱等方面的分析,以期为进一步开展葡萄WRKY转录因子基因家族的基因功能和分子进化机制的研究奠定理论基础。
1 材料和方法
1.1 数据收集
从 CRIBI Genomics(http://genomics.cribi.unipd.it)中下载欧亚种Vitis viniferaL.黑比诺PN4002全基因组的注释序列。从Pfam数据库(http://www.sanger.ac.uk/Software/Pfam/) 中下载WRKY结构域的隐马可夫模型(HMM)PF03106。
1.2 数据库搜索
用 HMMER3(http://hmmer.janelia.org/)软件包以PF03106隐马可夫模型对葡萄蛋白组进行搜索,所用选项为“cut_ga”。将得到的基因提交到NCBI上的保守结构域数据库CCD(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)中搜索WRKY结构域进行验证。
1.3 系谱发生分析
用WRKY基因的全长氨基酸序列作多序列联配,所用软件为ClustalX2,参数使用默认值。根据多序列联配的结果,使用MEGA5[14]软件构建系统发生树,所用算法为邻近法(neighbor joining,NJ),对构建的树进行自举检验,重复设为1 000。
1.4 共线性检测
所用软件为MicroSyn[15],采用该软件检测各基因所在基因组区段之间同源基因在数量和顺序上的保守程度。文中每个基因组区段包含100 个基因,在其中间位置为WRKY基因。
1.5 表达分析
Illumia RNA-Seq数 据 从NCBI的GEO(http://www.ncbi.nlm.nih.gov/gds)中下载,登陆号为GSE37743。表达量的单位以RPKM表示, 即每百万个测序片段(reads)中每个基因在1 kb长度上所对应的片段数。数据经log 2转换后提取WRKY基因的表达数据,制作热图(heatmap)。
2 结果与分析
2.1 葡萄WRKY基因的鉴定结果
以WRKY基因的HMM在葡萄全基因组蛋白序列中进行搜索,并将搜索到的基因提交到NCBI上的保守结构域数据库CCD中进行搜索,最后得到55个WRKY基因。根据WRKY基因在染色体上的位置给基因命名,这些基因的名称详见表1中的VvWRKY1~VvWRKY55。目前公布的12×葡萄基因组序列被组装成若干染色体序列,其中“chrN”(N代表染色体的编号,为1~19)表示将相应的序列骨架(contig)按位置和方向定位到染色体N上,无法定位到相应染色体上的序列骨架则组装到“chrUn”上。54个WRKY基因可以定位到18个染色体上,1个WRKY基因位于chrUn(如图1所示)。第4号染色体上WRKY基因的数量最多,为8个,第7号染色体上分布的数量次之,为7个,而在第6、9、11和18号染色体上分别只有1个WRKY基因,而第3号染色体上没有WRKY基因的分布。
2.2 进化分析
为了考察WRKY基因家族在葡萄中的系谱发生关系,在多序列联配的基础上,以NJ法构建WRKY基因家族的系谱发生树。为了更好地对葡萄WRKY基因家族进行分类,我们把葡萄的55个WRKY基因和拟南芥的70个WRKY基因作联合系谱发生树(如图2所示)。根据自举值的大小(一般>50%)和拟南芥中WRKY基因的分类信息,将葡萄的55个WRKY基因分为7个亚家族,分别命名为Ⅰ、Ⅱa、Ⅱb、Ⅱc、Ⅱd、Ⅱe和Ⅲ。其中,Ⅱa和Ⅱb的进化关系较近,Ⅱd和Ⅱe的进化关系较近。
在葡萄和拟南芥的联合进化树中,亚家族Ⅱ b包 含VvWRKY37、VvWRKY2、VvWRKY50、VvWRKY42、VvWRKY20、VvWRKY29、VvWRKY35和VvWRKY52等8个基因。利用MircoSyn软件对包含这些基因的基因组区段进行共线性分析。结果表明,在葡萄基因组内,VvWRKY2、VvWRKY42和VvWRKY50彼此之间存在显著的共线性关系,区段间所有同源基因对的同义突变率(Ks)平均值也比较相近(如表2)。同样,VvWRKY29、VvWRKY35和VvWRKY52之间也存在显著的共线性关系,区段间所有同源基因对的Ks平均值也比较相近。VvWRKY2、VvWRKY42和VvWRKY50分别位于第1、14和17号染色体上,而这3条染色体存在三体(triplet)的关系。据Jaillon等人[12]的研究结果,葡萄基因组内同源区段大部分以三体的形式出现,即一个基因组区域和另外两个区域存在同源关系,这种基因组三体的特性可能是由于葡萄基因组来源于古代的三个近缘基因组,发生了“古代六倍体化”事件。根据VvWRKY2、VvWRKY42和VvWRKY50之间的共线性关系和系谱发生树中亚家族Ⅱb成员间的拓扑关系,我们可以推测,位于第1号染色体上的VvWRKY2、第14号染色体上的VvWRKY42及第17号染色体上的VvWRKY50可能在葡萄物种的“古代六倍体化”过程中形成。同样,位于第10号染色体上的VvWRKY29、第12号染色体上的VvWRKY35及第19号染色体上的VvWRKY52也可能起源于这个“古代六倍体化”过程中。
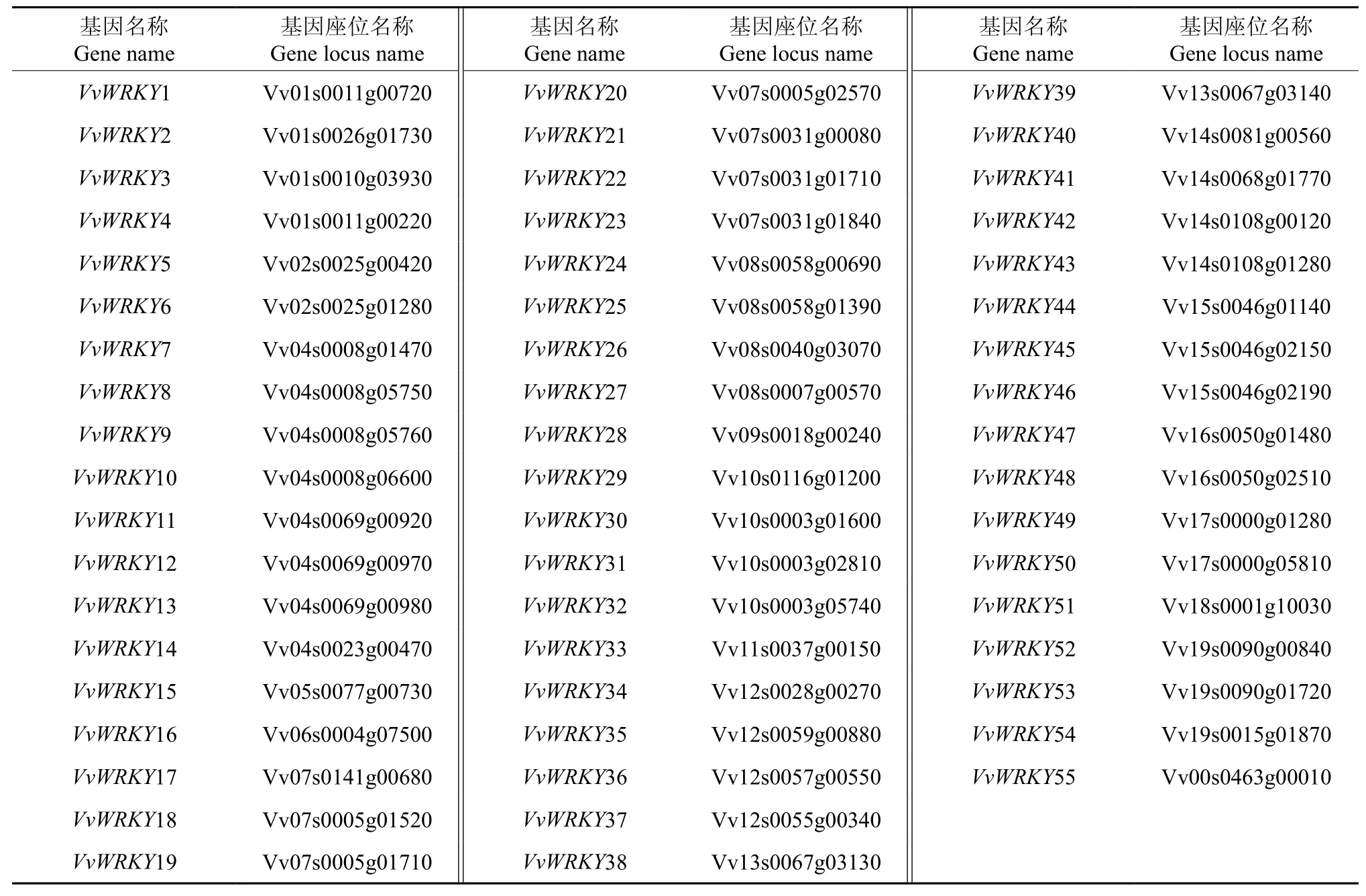
表1 葡萄WRKY基因的鉴定结果†Table 1 WRKY genes in grape
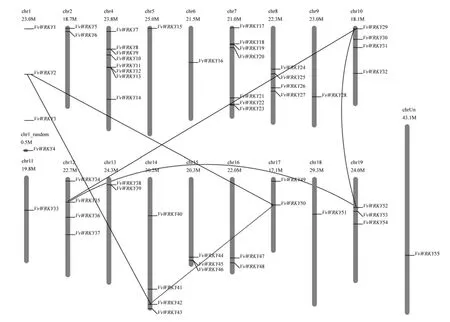
图1 葡萄WRKY基因在染色体上的定位图Fig.1 Chromosomal locations of WRKY genes in grape
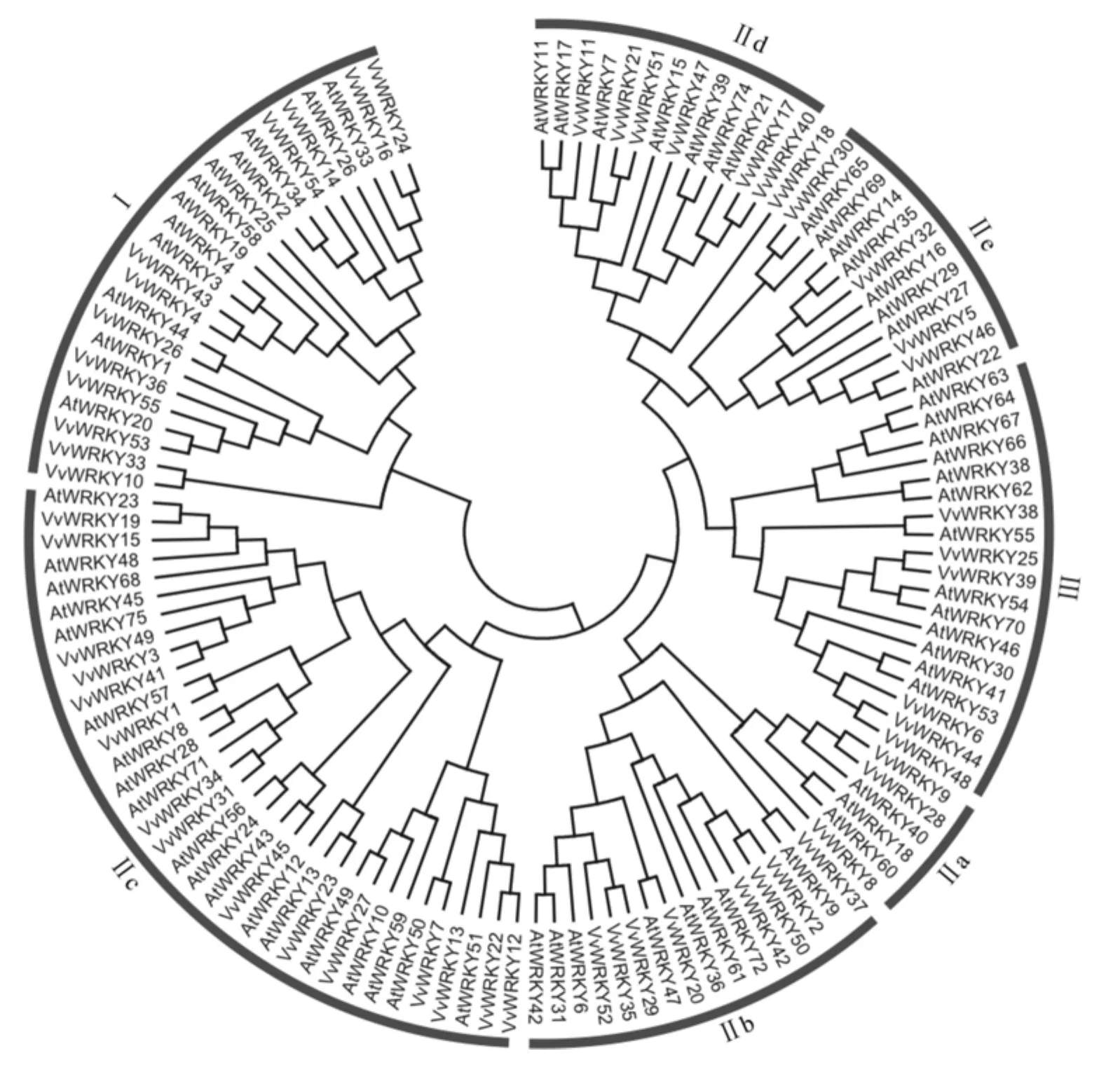
图2 葡萄和拟南芥中WRKY蛋白的联合系谱发生树Fig.2 Joined phylogenetic tree of WRKY proteins in grape and Arabidopsis
2.3 表达谱分析
GSE37743包含了葡萄非生物胁迫(紫外线和物理伤害)和生物胁迫(霜霉病侵染)的Illumia RNA-Seq数据,图3为各处理条件下跟对照相比表达值的热图。结果显示,有9个基因在植物受伤、紫外线照射和受到霜霉病侵染后,基因表达量显著提高。在这些基因中,有4个基因属于Ⅱb亚家族,其他5个基因则属于Ⅱc亚家族。

表2 亚家族Ⅱb内各基因之间的共线性关系†Table 2 Colinear relationships of the genes in Ⅱb subfamily
3 结论与讨论
植物转录因子在细胞发育、抗逆和信号转导等方面具有重要作用,而大部分的转录因子是以多基因家族的形式存在的。随着愈来愈多物种全基因组的公布,从全基因组水平上鉴定和分析转录因子家族,已成为基因组学研究的重点之一。WRKY转录因子家族在植物生长发育和植物防卫机制上具有重要作用。文中利用生物信息学的方法在葡萄全基因组中鉴定了55个WRKY基因,并对其进行了系谱分类和进化分析。系谱分析结果表明,葡萄WRKY基因家族的55个成员可以划分为7个亚家族。

图3 葡萄WRKY基因家族在非生物和生物胁迫条件下的表达水平热图Fig.3 Heat map of expression of WRKY gene family under different abiotic and biotic stress
在其亚家族Ⅱb中的8个WRKY基因中,VvWRKY2、VvWRKY42和VvWRKY50及VvWRKY29、VvWRKY35和VvWRKY52可能起源于形成葡萄物种的一次“古代六倍体化”过程中;另外2个成员VvWRKY20和VvWRKY37和其他成员间不存在共线性关系,我们推测,VvWRKY20和VvWRKY37可能存在于“古代六倍体化”事件之前。从表达数据来看,VvWRKY20、VvWRKY37、VvWRKY50和VvWRKY52对生物和非生物胁迫的反应,具有相似的表达谱模式(图3),说明这4个WRKY基因在“古代六倍体化”事件之前已经参与逆境胁迫的调控网络,而VvWRKY2、VvWRKY42、VvWRKY29和VvWRKY35在进化过程中其表达特性发生趋异分化现象。
参考文献:
[1] Latchman D S.Transcription factors: an overview[J].Int J Biochem Cell Biol,1997,29(12):1305-1312.
[2] 张 莉,苏曼琳.植物抗旱基因HDCS1的克隆和表达载体的构建[J].中南林业科技大学学报,2012,32(6):115-117.
[3] Hara K, Yagi M, Kusano T,et al.Rapid systemic accumulation of transcripts encoding a tobacco WRKY transcription factor upon wounding [J].Mol Gen Genet, 2000, 263(1):30-37.
[4] Luo M, Dennis E S, Berger F,et al.MINISEED3 (MINI3), a WRKY family gene, andHAIKU2 (IKU2), a leucine-rich repeat(LRR)KINASEgene, are regulators of seed size inArabidopsis[J].Proc Natl Acad Sci USA,2005,102(48): 17531-17536.
[5] Robatzek S, Somssich I E.A new member of theArabidopsisWRKY transcription factor family, AtWRKY6, is associated with both senescence- and defence-related processes [J].Plant J,2001,28(2):123-133.
[6] Pandey S P, Somssich I E.The role of WRKY transcription factors in plant immunity[J].Plant Physiol, 2009, 150(4):1648-1655.
[7] Wu X, Shiroto Y, Kishitani S,et al.Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter [J].Plant Cell Rep,2009,28(1):21-30.
[8] Wang Z, Zhu Y, Wang L,et al.A WRKY transcription factor participates in dehydration tolerance inBoea hygrometricaby binding to the W-box elements of the galactinol synthase(BhGolS1) promoter [J].Planta, 2009,230(6):1155-1166.
[9] Jiang Y, Deyholos M K.Functional characterization of Arabidopsis NaCl-inducible WRKY25 and WRKY33 transcription factors in abiotic stresses [J].Plant Mol Biol, 2009,69(1-2): 91-105.
[10] Li S, Fu Q, Huang W,et al.Functional analysis of anArabidopsistranscription factor WRKY25 in heat stress [J].Plant Cell Rep,2009, 28(4): 683-693.
[11] 彭丽丽,姜卫兵,健 韩.源库关系变化对果树产量及果实品质的影响[J].经济林研究,2012,30(3):134-140.
[12] Jaillon O, Aury J M, Noel B,et al.The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla [J].Nature, 2007, 449(7161): 463-467.
[13] Marchive C, Leon C, Kappel C,et al.Over-expression of VvWRKY1 in grapevines induces expression of jasmonic acid pathway-related genes and confers higher tolerance to the downy mildew [J].PLoS One, 2013, 8(1): 54185.
[14] Tamura K, Peterson D, Peterson N,et al.MEGA5: molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance, and maximum parsimony methods [J].Mol Biol Evol, 2011, 28(10): 2731-2739.
[15] Cai B, Yang X, Tuskan G A,et al.MicroSyn: a user friendly tool for detection of microsynteny in a gene family [J].BMC Bioinformatics, 2011, (12):79.
猜你喜欢
华北电力大学学报(社会科学版)(2021年2期)2021-07-21
科学与财富(2021年3期)2021-03-08
——以多重共线性内容为例
长沙航空职业技术学院学报(2019年2期)2019-07-13
温州大学学报(自然科学版)(2019年2期)2019-06-04
广州大学学报(自然科学版)(2019年1期)2019-05-07
图书馆理论与实践(2018年2期)2018-01-28
中国奶牛(2017年2期)2017-03-22
求学·理科版(2017年1期)2017-03-02
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
