
同形溞(Daphnia similis)对幽蚊幼虫反捕食策略的模拟研究*
2013-05-28 07:35王文侠舒婷婷陈非洲
湖泊科学 2013年2期
王文侠,舒婷婷,李 静,陈非洲
(1:中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,南京 210008)
(2:中国科学院大学,北京 100049)
淡水生态系统中,幽蚊幼虫等无脊椎动物不仅可以直接捕食浮游动物枝角类,而且可以通过间接方式如释放信息素对其产生影响[1-3].信息素自Karlson 等[4]研究以来得到广泛关注,捕食者能通过信息素对被捕食者的形态、行为、生活史及生理等各方面产生影响[5].淡水枝角类溞(Daphnia)由于分布广泛,易于培养,且条件适宜时进行孤雌生殖已成为研究捕食者信息素的模式动物[6-7].研究发现溞在胚胎期第3 次脱膜后便有了感知信息素的能力并出现形态上的可塑性反应[7].
溞在幽蚊幼虫等无脊椎捕食者释放的信息素刺激下产生不同的生活史变化[8-9].一方面捕食者信息素导致的可塑性反应存在种的特异性,如相同实验条件下蚤状溞(Daphnia pulex)体长无显著变化,玫瑰溞(Daphnia rosea)体长显著变长[10].另一方面对同一物种的研究也出现不同的结果[11-13].表型可塑性引起广泛关注,不仅是因为它们在生态系统食物网中发挥重要的生态效应,而且它们会导致进化的产生[14],为更好地了解物种进化和群落演替提供理论支持.
幽蚊幼虫是湖泊中重要的无脊椎动物,在淡水湖泊上层食物网的能量转换中起着关键性作用[15],它可以控制猎物的种类、丰度及空间分布[16-17].当其密度达到0.4 只/L 时,每天会消耗50%的浮游动物生产量或13%的总浮游动物生物量[18].由于幽蚊幼虫口裂宽度有限,通常优先捕食较小个体,大于其口裂宽度的猎物有更大的逃生机会[19].
同形溞(Daphnia similis)是淡水湖泊中常见的枝角类,是太湖等湖泊的春季优势种[20].本实验的主要目的是研究其在幽蚊幼虫环境中生活史参数的变化,分析其对无脊椎动物的反捕食策略及可能的适应机制.
1 材料与方法
实验用同形溞和幽蚊幼虫均取自太湖梅梁湾旁水泥池.幽蚊幼虫选择2 ~3 龄个体.同形溞分离出怀卵个体若干,置于烧杯中用培养液单克隆培养,加入浓度为2×106cells/L 用BG-11 培养液培养的栅藻作为食物,单克隆培养至第3 代,挑选出生12 ~24 h 的个体90 只(平均体长0.73 mm).
本实验设3 个处理,每个处理3 个重复,首先将90 只同形溞幼体随机放入9 个500 ml 的烧杯中,每个烧杯放10 只.处理1(加幽蚊):烧杯中加入3 只幽蚊幼虫,将其固定于直径5 cm 的特制圆柱形网(孔径约38 μm)内部使其与同形溞分开,并用特制培养液培养.处理2(加幽蚊培养液):烧杯中不加入幽蚊幼虫,同形溞的培养液为实验前一天培养过幽蚊幼虫的水,此水事先用孔径约38 μm 的生物网过滤.处理3(对照):烧杯中不加幽蚊幼虫,同形溞的培养液为特制培养液.为避免圆柱形网可能造成的干扰,每只烧杯均在相同的位置加入同样的圆柱形网.为避免自然水体中可能存在的信息物质,实验所用特制培养液为稀释的 BG-11 培养液,主要离子浓度为:Na+34 μmol/L,Mg2+30 μmol/L,Ca2+26 μmol/L,NO3-34 μmol/L,30 μmol/L,Cl-52 μmol/L.为减少代谢废物的干扰,每24 h 更换培养液.实验在室温下进行,于2011年8月12日开始,持续10 d.每24 h 测量同形溞体长并记录怀卵和产仔状况.
同形溞体长在显微镜下测量,文中图形绘制使用Excel 2007.实验数据采用方差分析和最小显著差法(LSD)多重比较分析并由SAS 软件实现.
2 结果与分析
实验阶段同形溞个体无死亡,实验结束时加幽蚊和加幽蚊培养液处理同形溞的平均体长分别达到2.26 ±0.08 mm 和2.24 ±0.10 mm(图1a).方差分析表明这两个处理同形溞的平均体长无显著差异,但都显著大于对照(1.99 ±0.09 mm)(P <0.05).
第5 d 起各处理同形溞均开始产幼仔,第7 d 加幽蚊处理同形溞的平均产仔数达到最大值(13 只/雌体).加幽蚊处理平均产仔数除第5、第6 d 与对照无显著差异外,其余4 d 显著多于对照(P <0.05).加幽蚊培养液处理平均产仔数在第8、第9 d 显著大于对照(P <0.05),其余时间与对照无显著差异.加幽蚊处理平均产仔数第7、第9 d 显著大于加幽蚊培养液处理(P <0.05),其余时间两处理无显著差异(图1b).
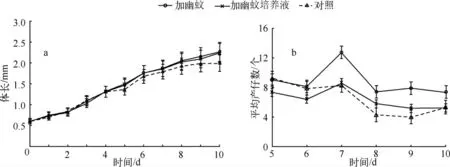
图1 不同处理同形溞平均体长(a)和平均产仔数(b)的变化Fig.1 Average body-length(a)and average number of offspring(b)of Daphnia similis in different treatments
实验期间加幽蚊处理同形溞的累积产仔数最多,达262 只;其次为加幽蚊培养液处理,达233 只;对照组的累积产仔数只有169 只.方差分析表明,加幽蚊与加幽蚊培养液处理同形溞累积产仔数无显著差异,但都显著大于对照组(P <0.05).
3 讨论
3.1 幽蚊幼虫对同形溞生活史参数的影响
加幽蚊及加幽蚊培养液处理的同形溞平均体长显著大于对照,说明幽蚊幼虫释放的信息素刺激了同形溞的生长,使其体长变长,从而减小了被幽蚊幼虫捕食的风险[21].Dodson 等[22]研究表明,第四龄期幽蚊幼虫捕食溞体长一般不超过1.30 mm,并且优先捕食较小个体.Krylov[23]测量了幽蚊幼虫对小个体溞(0.77 mm)和大个体溞(1.82 mm)的攻击效率,发现幽蚊幼虫对小个体溞的攻击效率更高.在将大、小溞1∶1 混合时,幽蚊幼虫捕食小个体溞显著多于大个体溞.虽然大个体溞与幽蚊幼虫的偶遇率更大,但随着溞体长的增大,幽蚊幼虫对其攻击效率变小[24-25].
平均产仔数和累积产仔数是同形溞繁殖能力大小的标志.实验结果表明在幽蚊幼虫信息素作用下同形溞的繁殖能力显著增强.这说明同形溞感知捕食风险后可以通过自身繁殖机制的调节来增加后代数量,从而保持种群平衡,体现了同形溞的社会性[26].
由于较多的能量能保证体长增长和繁殖能力增强,所以同形溞在感知捕食风险后将摄取更多食物.而在自然条件下觅食活动的增加意味着同形溞将面对更大的被捕食风险[27].事实上,由于随着行为活动的增加,摄入的能量和被捕食的风险同时增加,所以被捕食者经常面对摄食获能与存活间的行为导向型权衡(behavioral mediated trade-offs)[28].环境中的激素水平可增强同形溞的食欲和生理机能,使其有更高的代谢率从而促进其生长和繁殖[29],但这种促进作用是有一定限度的[30].而溞类由行为活动带来的风险可以通过垂直迁移而减弱[31].所以同形溞感知捕食风险后将用于生长、繁殖及行为活动等的能量进行权衡和再分配[32]形成独特的反捕食策略.从个体角度可以降低其被捕食的风险,维持其生长繁殖;从种群角度来讲可以增加后代数量,维持种群平衡;从群落与生态系统角度,可以保证各营养级间的平衡,维持生态系统结构和功能[33].
对同形溞体长和产仔数据分析可知,培养过幽蚊幼虫的过滤水与存在幽蚊幼虫的水引起的效果一致但程度减弱,这可能与幽蚊幼虫产生的信息素具有一定的挥发性有关[34].湖泊生态系统中脊椎和无脊椎捕食者同时存在,两者信息素对溞生活史参数影响不同[35].一般来说,脊椎捕食者信息素使溞的体长变小,初次产仔时间缩短[36-37].溞的生活史参数取决于起主导作用的捕食者[2,38].对俄罗斯Maly Okunenok 湖的野外研究发现,当脊椎捕食者数量减少而无脊椎捕食者数量增多时溞的平均体长显著变长[23].
3.2 枝角类溞在幽蚊幼虫信息素刺激下生活史的变化
作为研究捕食关系的模式动物,幽蚊幼虫-溞得到广泛研究,多数结果一致,溞可以通过增加颈齿数量[39-40]、改变身体弯曲度等形态变化[41],垂直迁移、躲避信息素浓度高的区域等行为变化[42-44]和增大体长、推迟初次繁殖时间等生活史变化[6,21]来降低被幽蚊幼虫捕食的危险.但其他研究出现不同的结果[37].溞的生活史变化是其在捕食者信息素作用下产生的最显著反应,对其分析将有助于了解溞反捕食的表型可塑性,进而更好地了解其对整个湖泊生态系统的影响.
首先,不同物种适应环境能力的差异可造成不同的表型可塑性,溞生活史的可塑性反应受其遗传基因的控制[45].不同溞本身的体长差异会对其表型可塑性造成很大影响,如蚤状溞体长通常比玫瑰溞体长大,条件相同时在幽蚊幼虫信息素刺激下玫瑰溞体长显著变长而蚤状溞体长并无显著变化[10].此外不同物种习性及昼夜垂直迁移模式的不同会造成其与捕食者偶遇率不同,从而产生不同的表型可塑性[40].其次,同一物种当自身体长或处理时间不同时表型可塑性不同,如用幽蚊幼虫信息素分别处理刚出生和出生72 h 的蚤状溞其初次繁殖时间分别比对照推迟1.2 d 和1.7 d[46].对前五龄期盔形溞(Daphnia galeata)用幽蚊幼虫信息素处理时其体长显著变长[21],而对第六龄期的盔形溞处理时其体长无显著变化[47-48].原因是溞在不同阶段对捕食者信息素的反应不同.最后,溞密度[49]及食物质量和浓度[50-51]对其本身生长和繁殖有影响,因此它们会使溞产生不同的表型可塑性反应.
虽然不同物种或不同生境下表型可塑性反应不同,但被捕食者所做出的决策并不是随机和盲目的,而是针对捕食者、环境及自身机能状况制定的最佳反捕食策略.同形溞在幽蚊幼虫信息素的作用下产生反捕食的表型可塑性,当此可塑性变化适应环境时便通过自然选择作用得以积累.这样一系列细微、连续的有利变异由于母体效应得以延续,逐渐提高后代的生存能力,所以说表型可塑性不仅会导致进化而且会决定进化的方向[14].
致谢:感谢于谨磊、姬娅婵、吴虹飞在实验过程中给予的帮助,感谢审稿人的宝贵意见.
[1]Brönmark C,Hansson LA.Chemical communication in aquatic systems:an introduction.Oikos,2000,88(1):103-109.
[2]Stibor H,Lampert W.Components of additive variance in life-history traits of Daphnia hyalina:seasonal differences in the response to predator signals.Oikos,2000,88(1):129-138.
[3]王松波,谢 平,朱龙喜.养殖水体中的微型裸腹溞对高浓度鱼类化学信息素的反应:以生命表研究为例.水生生物学报,2008,32(6):868-873.
[4]Karlson P,Lüscher M.“Pheromones”:a new term for a class of biologically active substances.Nature,1959,183:55-56.
[5]Kusch J.Behavioural and morphological changes in ciliates induced by the predator Amoeba proteus.Oecologia,1993,96:354-359.
[6]Weetman D,Atkinson D.Antipredator reaction norms for life history traits in Daphnia pulex:dependence on temperature and food.Oikos,2002,98(2):299-307.
[7]De Meester L.Neutral markers,ecologically relevant traits,and the structure of genetic variation in Daphnia.Aquatic Ecology,1997,31:79-87.
[8]Zadereev YS.Maternal effects,conspecific chemical cues,and switching from parthenogenesis to gametogenesis in the cladoceran Moina macrocopa.Aquatic Ecology,2003,37:251-255.
[9]柯志新,黄良民.溞类(Daphnia)反捕食的表型可塑性及其研究进展.湖泊科学,2009,21(6):758-767.
[10]Sell AF.Morphological defenses induced in situ by the invertebrate predator Chaoborus:comparison of responses between Daphnia pulex and D.rosea.Oecologia,2000,125:150-160.
[11]Tollrian R.Chaoborus crystallinus predation on Daphnia pulex:can induced morphological changes balance effects of body size on vulnerability?Oecologia,1995,101:151-155.
[12]Vuorinen L,Ketola M,Walls M.Defensive spine formation in Daphnia pulex Leydig and induction by Chaoborus crystallinus De Geer.Limnology and Oceanography,1989,34(1):245-248.
[13]Walls M,Caswell H,Ketola M.Demographic costs of Chaoborus-induced defenses in Daphnia pulex:a sensitivity analysis.Oecologia,1991,87:43-50.
[14]Price TD,Qvarnström A,Irwin DE.The role of phenotypic plasticity in driving genetic evolution.Proceedings of the Royal Society B,2003,270:1433-1440.
[15]Moore MV,Yan ND,Pawson T.Omnivory of the larval phantom midge(Chaoborus spp.)and its potential significance for freshwater planktonic food webs.Canadian Journal of Zoology,1994,72(11):2055-2065.
[16]Yan ND,Keller W,Maclsaac HJ et al.Regulation of zooplankton community structure of an acidified lake by Chaoborus.Ecological Applications,1991,1(1):52-65.
[17]Riessen HP,Sommerville JW,Chiappari C et al.Chaoborus predation,prey vulnerability,and their effect in zooplankton communities.Canadian Journal of Fisheries and Aquatic Sciences,1988,45(11):1912-1920.
[18]Kajak Z,Ranke-Rybickova B.Feeding and production efficiency of Chaoborus flavicans (Meigen)(Diptera,Culicidae)larvae in eutrophic and dystrophic lakes.Hydrobiologia,1970,17:225-232.
[19]Gliwicz ZM,Umana G.Cladoceran body size and vulnerability to copepod predation.Limnology and Oceanography,1994,39:419-424.
[20]李 静,陈非洲.太湖夏秋季大型枝角类(Daphnia)种群消失的初步分析.湖泊科学,2010,22(4):552-556.
[21]Weber A,Declerck S.Phenotypic plasticity of Daphnia life history traits in response to kairomones:genetic variability and evolutionary potential.Hydrobiologia,1997,360:89-99.
[22]Dodson SI.Zooplankton competition and predation:an experimental test of the size-efficiency hypothesis.Ecology,1974,55:605-613.
[23]Krylov PI.Density-dependent predation of Chaoborus flavicans on Daphnia longispina in a small lake:the effect of prey size.Hydrobiologia,1992,239:131-140.
[24]Swift MC,Fedorenko AY.Some aspects of prey capture by Chaoborus larvae.Limnology and Oceanography,1975,18:795-798.
[25]Pastorok RA.Selection of prey by Chaoborus larvae:a review and new evidence for behavioral flexibility.Evolution and E-cology of Zooplankton Communities,1980,3:538-554.
[26]Thomson RL,Forsman JT,Mönkkönen M et al.Risk taking in natural predation risk gradients:support for risk allocation from breeding pied flycatchers.Animal Behaviour,2011,82:1443-1447.
[27]Hanazato T.Anthropogenic chemicals(insecticides)disturb natural organic chemical communication in the plankton community.Environmental Pollution,1999,105:137-142.
[28]Biro PA,Abrahams MV,Post JR et al.Behavioural trade-offs between growth and mortality explain evolution of submaximal growth rates.Journal of Animal Ecology,2006,75:1165-1171.
[29]Pestana JLT,Loureiro S,Baird DJ et al.Fear and loathing in the benthos:Responses of aquatic insect larvae to the pesticide imidacloprid in the presence of chemical signals of predation risk.Aquatic Toxicology,2009,93:138-149.
[30]Bettridge C,Lehmann J,Dunbar RIM.Trade-offs between time,predation risk and life history,and their implications for biogeography:A systems modelling approach with a primate case study.Ecological Modelling,2010,221(5):777-790.
[31]Peacor SD,Pangle KL,Vanderploeg HA.Behavioral response of Lake Michigan Daphnia mendotae to Mysis relicta.Journal of Great Lakes Research,2005,31:144-154.
[32]Taylor BE,Gabriel W.To grow or not to grow:optimal resource allocation for Daphnia.The American Naturalist,1992,139:258-266.
[33]Garay-Narváez L,Ramos-Jiliberto R.Induced defenses within food webs:The role of community trade-offs,delayed responses,and defense specificity.Ecological Complexity,2009,6:383-391.
[34]Lass S,Spaak P.Chemically induced anti-predator defences in plankton:a review.Hydrobiologia,2003,491:221-239.
[35]Riessen HP.Predator-induced life history shifts in Daphnia:a synthesis of studies using meta-analysis.Canadian Journal of Fisheries and Aquatic Sciences,1999,56:2487-2494.
[36]Reede T.Effects of neonate size and food concentration on the life history responses of a clone of the hybrid Daphnia hyalina×galeata to fish kairomones.Freshwater Biology,1997,37:389-396.
[37]Spaak P,Vanoverbeke J,Boersma M.Predator-induced life-history changes and the coexistence of five taxa in a Daphnia species complex.Oikos,2000,89:164-174.
[38]Stibor H.Predator induced life-history shifts in a freshwater cladocera.Oecologia,1992,92:162-165.
[39]Repka S,Pihlajamaa K.Predator-induced phenotypic plasticity in Daphnia pulex:uncoupling morphological defenses and life history shifts.Hydrobiologia,1996,339:67-71.
[40]Mirza RS,Pyle GG.Waterborne metals impair inducible defences in Daphnia pulex:morphology,life-history traits and encounters with predators.Freshwater Biology,2008,54:1016-1027.
[41]Weber A,Vesela S.Optimising survival under predation:chemical cues modify curvature in Daphnia galeata.Aquatic E-cology,2002,36:519-527.
[42]Szulkin M,Dawidowicz P,Dodson SI.Behavioural uniformity as a response to cues of predation risk.Animal Behaviour,2006,71:1013-1019.
[43]De Meester L,Cousyn C.The change in phototactic behaviour of a Daphnia magna clone in the presence of fish kairomones:the effect of exposure time.Hydrobiologia,1997,360:169-175.
[44]Michels E,De Meester L.Inter-clonal variation in phototactic behaviour and key life-history traits in a metapopulation of the cyclical parthenogen Daphnia ambigua:the effect of fish kairomones.Hydrobiologia,2004,522:221-233.
[45]Lass S,Vos M,Wolinska J et al.Hatching with the enemy:Daphnia diapausing eggs hatch in the presence of fish kairomones.Chemoecology,2005,15:7-12.
[46]Gorur G,Lomonaco C,Mackenzie A.Phenotypic plasticity in host-plant specialisation in Aphis fabae.Ecological Entomology,2005,30(6):657-664.
[47]Riessen HP,Sprules WG.Demographic costs of antipredator defenses in Daphnia pulex.Ecology,1990,71(4):1536-1546.
[48]Tollrian R.Predator-induced morphological defences:costs,life history shifts,and maternal effects in Daphnia pulex.E-cology,1995,76:1691-1705.
[49]Jeschke JM,Tollrian R.Density-dependent effects of prey defenses.Oecologia,2000,123:391-396.
[50]Gliwicz ZM,Maszczyk P.Daphnia growth is hindered by chemical information on predation risk at high but not at low food levels.Oecologia,2007,150:706-715.
[51]S'lusarczyk M.Food threshold for diapause in Daphnia under the threat of fish predation.Ecology,2001,82(4):1089-1096.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
延边大学学报(社会科学版)(2022年6期)2022-12-06
昆明医科大学学报(2022年1期)2022-02-28
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
现代畜牧科技(2021年6期)2021-07-16
中国畜牧杂志(2020年8期)2020-08-16
汉字汉语研究(2019年4期)2019-03-04
民族古籍研究(2018年1期)2018-05-21
西南医科大学学报(2015年1期)2015-08-22
