
两用核不育系P88S愈伤组织的诱导及其遗传转化研究
2013-09-04 03:40袁定阳夏石头
湖南农业科学 2013年13期
崔 看 ,成 平 ,陈 锦 ,袁定阳 ,夏石头
(1.湖南农业大学生物科学技术学院,植物激素与生长发育湖南省重点实验室,湖南 长沙 410128;2.湖南省农业信息与工程研究所,湖南 长沙 410125;3.杂交水稻国家重点实验室,湖南 长沙 410125)
水稻(Oryza sativa L.)是人类最重要的粮食作物之一,传统遗传育种技术在提高水稻产量和改善米质等方面做出了巨大贡献[1]。但由于生态环境恶化和水稻种质资源匮乏的限制,传统的遗传改良方法已经不能够满足现代农业对新型种质资源的需求。P88S是由湖南杂交水稻研究中心罗孝和研究员在2003年利用双向系选(在长沙长日低温的条件下选不育,在海南短日低温下选相对可育)和温光加再生强化选择而选育出的异交结实率高、配合力强的超级杂交水稻两用核不育系,在超级稻生产上得到了广泛应用,但其对稻瘟病抗性较弱[2-3]。利用基因工程技术将植物抗病性状相关基因导入水稻基因组中,可以作为获得新种质资源的一种重要手段,在实现超级稻高产、优质、多抗、稳产目标中发挥重要作用[4]。文章以两用核不育系P88S为材料,研究了其愈伤组织的诱导及植物抗病基因snc1在其愈伤组织中的遗传转化,以期通过诱导、分化和继代培养获得高抗病超级稻新种质。
1 材料与方法
1.1 材料
超级杂交水稻两用核不育系P88S,由湖南杂交水稻研究中心罗孝和研究员提供;遗传转化载体为夏石头教授课题组改造的pG229质粒,含有Kan+与Basta双重抗性标记;供体菌为农杆菌EHA105,功能获得型突变基因Atsnc1[5],由加拿大英属哥伦比亚大学Michael Smith Laboratories李昕教授实验室惠赠。
1.2 方法
1.2.1 P88S愈伤组织的诱导 采取完全随机化设计,参照文献[6-7]选用4种诱导培养基MB、N6、NB和NMB进行愈伤组织诱导与继代培养,将P88S成熟胚分别接种于 MB、N6、NB、NMB 4种培养基上进行愈伤组织的诱导,2,4-D浓度分别设为2.0、2.5、3.0 mg/L,6-BA 的浓度分别设为 0.1、0.2、0.3 mg/L,并添加0.5 g/L酸水解酪蛋白(CH)和脯氨酸(Pro)。诱导7 d或继代培养12 d后,分别统计芽长、愈伤组织重量和诱导率等性状,以筛选出最适合P88S愈伤组织诱导和继代培养的最适培养基与最适生长物质浓度配比。
1.2.2 遗传转化植株的获得与检测 农杆菌的浸染转化、抗性愈伤组织的筛选、转化植株的检测按照成平等[8]方法,转化植株的检测采用试剂盒(北京康为世纪生物科技有限公司),提取转化植株的DNA,进行PCR检测。正向引物序列为:GTGGAGTTCCCATCTGAACATC,反向引物序列为:CCCATTTTGATTGCTGGAAAG。
2 结果与分析
2.1 P88S愈伤组织诱导培养母液的筛选
将 P88S的成熟胚分别接种于 MB、N6、NB、NMB培养基上(含3.0 mg/L 2,4-D+0.2 mg/L 6-BA+0.5 g/L酸水解酪蛋白+0.5 g/L脯氨酸)诱导7 d后,观察和统计各培养基上愈伤组织的颜色、重量和芽长等。试验结果如表1所示,P88S在MB、NMB、N6培养基上诱导时,愈伤组织颜色淡黄,颗粒圆润饱满,而在NB培养基上诱导时,愈伤组织颗粒有褐化现象且颗粒较小。如图1所示,P88S在MB培养基上P88S诱导的芽最长,为2.797 cm,比在 N6、NB、NMB 培养基上分别高出 0.749、0.821、0.887 cm,差异达到极显著水平;在NB培养基上诱导的P88S愈伤组织重量最轻,为0.004 6 g,极显著低于其余3种培养基上诱导的愈伤组织;NMB培养基的诱导率最高,达95.66%,极显著高于MB培养基的愈伤组织诱导率。综合考虑各因素,NMB培养基是最适合P88S愈伤组织诱导的培养基。
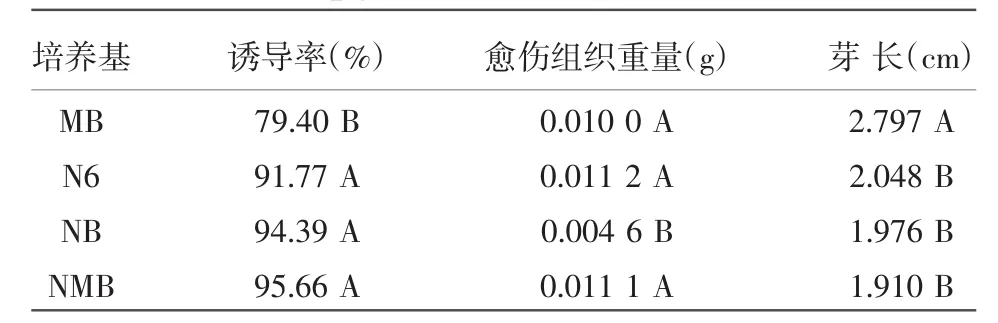
表1 P88S愈伤组织在不同培养基上诱导的芽长、愈伤组织重量和诱导率
2.2 P88S愈伤组织诱导培养基中植物生长物质浓度的选择
不同浓度2,4-D与6-BA对P88S愈伤组织诱导的影响如表2所示,不同浓度的2,4-D对于P88S芽长的影响不显著,而对于愈伤组织重量的影响差异达到极显著水平,当2,4-D质量浓度为3.0 mg/L时,P88S愈伤组织重量最大,平均达0.011 g,极显著高于其他2个浓度处理的愈伤组织重量。当添加3.0 mg/L的2,4-D时,诱导率达到94.70%,极显著高于添加2.5 mg/L 2,4-D时的诱导率,表明2,4-D质量浓度对P88S愈伤组织诱导率的有重要影响。而6-BA对芽长的影响明显,浓度为0.3 mg/L 6-BA处理的芽长极显著高于其他2个浓度处理的芽长,其愈伤组织诱导率与0.2 mg/L 6-BA处理的愈伤组织诱导率相近,但显著高于0.1 mg/L 6-BA处理的愈伤组织诱导率。综合考虑各因素,确定2,4-D最适质量浓度为3.0 mg/L,6-BA最适质量浓度为0.2 mg/L。
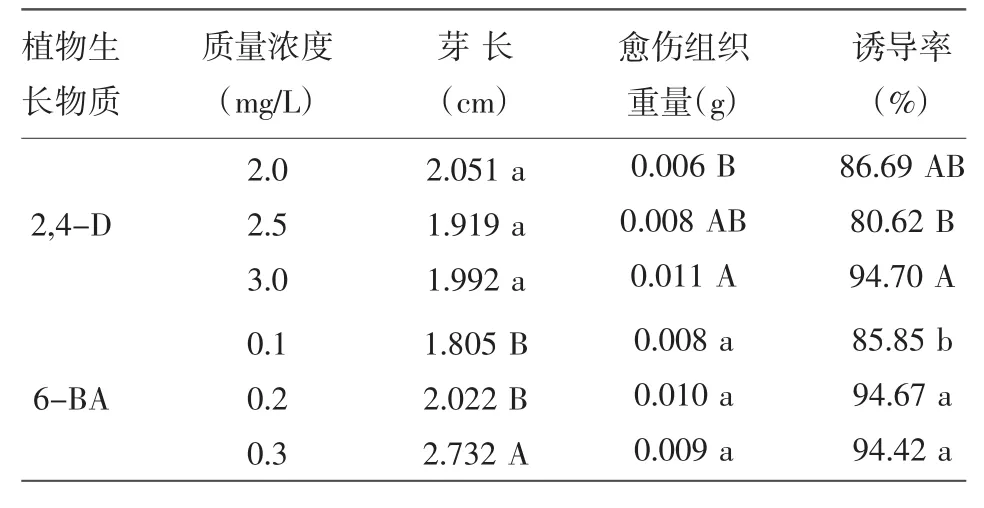
表2 2,4-D和6-BA对P88S愈伤组织诱导的影响
2.3 酸水解酪蛋白和脯氨酸对水稻成熟胚愈伤组织诱导的影响
根据愈伤组织对渗透压的要求,在试验中添加了 0.5 g/L酸水解酪蛋白(CH)和脯氨酸(Pro),统计结果表明,添加CH和Pro的培养基,芽长为2.103 cm,愈伤组织重量为0.011 2 g,诱导率为94.75%;不添加CH和Pro的培养基,芽长为2.690 cm,愈伤组织重量为0.006 9 g,诱导率为91.83%。与不添加CH和Pro相比,添加CH和Pro的培养基芽长较短,其诱导率稍高,并能显著提高愈伤组织的质量。且添加CH和Pro后,愈伤组织生长旺盛,颗粒圆润,颜色嫩黄鲜艳;而不添加CH和Pro,愈伤组织生长状态较差,颗粒干瘪较小,且有褐化现象。
2.4 P88S愈伤组织最适继代培养基的确定
愈伤组织一般都要继代1次,即培养约12d后才进行转化,且转化前会将愈伤组织一分为二,造成一定创伤以有利于农杆菌的浸染。在对同一批愈伤组织进行MB、N6、NB、NMB培养基继代培养,并观测继代培养生长和增重情况后,发现NMB培养基是P88S愈伤组织继代培养的最适培养基,其增重最大,为0.233 g,增重达185%,极显著高于N6和NB培养基上愈伤组织的增重(图1)。
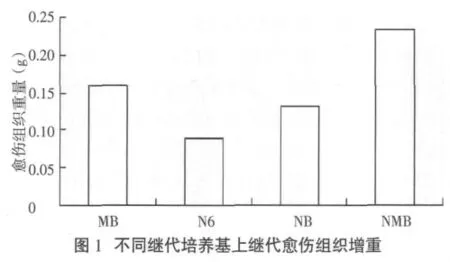
2.5 P88S愈伤组织最适分化培养基的确定
分化培养的目的是让愈伤组织重新分化成小苗,比较研究了DL和NMB两种母液对P88S愈伤组织分化的影响,试验结果表明,不同碳源对愈伤组织分化有一定影响,以麦芽糖作为碳源时,分化启动较快,愈伤组织生长较迅速,7d左右就可见新的愈伤组织小颗粒,且愈伤组织紧致,绿豆大小的颗粒,出绿点时间为10~15d;以蔗糖作为碳源时,愈伤组织生长缓慢,一般2个星期左右才会有新愈伤组织,且愈伤组织松散,一夹即碎,出绿点时间为15~20d。P88S在DL分化培养基中培养25d后分化出芽,而在NMB分化培养基中培养25d后,愈伤组织有褐化死亡的迹象,且仅有部分愈伤组织出现绿点。因此,选用麦芽糖为碳源的DL分化培养基作为P88S的最适分化培养基。
2.6 遗传转化植株的获得与检测
当分化培养基中绿芽长到长于2 cm后,将其从愈伤组织上剥离,转入生根培养基中。大约2周幼苗就长出完整的根系,加水炼苗2~3d后将其移出,洗净掉根上的培养基,剪掉变黄的叶子,移栽到稻田土中(其转化流程见图2)。遗传转化植株验证时,取移栽到稻田土中T0代转化植株叶片,用试剂盒法提取小量DNA,利用snc1基因的特异引物进行PCR扩增检测5株P88S转化植株,结果显示P88ST0-5中检测到了目的基因snc1的存在(图2中h)。
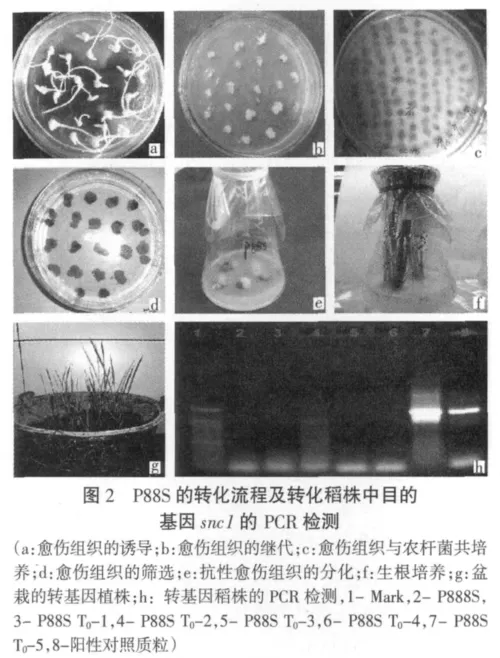
3 结论与讨论
以超级杂交水稻两用核不育系P88S为材料,研究发现P88S成熟胚愈伤组织的最适诱导和继代培养基是NMB培养基,其中2,4-D和6-BA的最适浓度分别为3.0 mg/L和0.2 mg/L,而添加0.5 g/L的酸水解酪蛋白和脯氨酸有利于愈伤组织的诱导。P88S成熟胚愈伤组织的最适分化培养基是DL分化培养基,麦芽糖为抗性愈伤组织分化的最适碳源。并通过农杆菌介导遗传转化法共获得5株转化稻株,经PCR检测其中1株含目的基因s nc1。
拟南芥 SNC1(suppressor of npr1-1,constitutive1)基因[5]是一种重要的水杨酸诱导型广谱抗病基因,该基因功能获得型突变导致snc1突变体植株组成性表达病程相关蛋白,能够组成性激活植物的抗病机制,并对细菌病原体Pseudomonas syringae pv maculicola ES4326(P.s.m.ES4326)和卵菌病原体 Hyaloperonospora arabidopsidis Noco2(H.a.Noco2)等病原物产生广谱抗性。因为Atsnc1基因是一种获得功能型突变基因,可以组成性表达致病机理相关(PR)基因,从而激活抗病性反应,P88S转化稻株中snc1基因是否有同样的生理功能,还有待进一步研究。
[1]彭雪明,张小平,周文新,等.中国超级稻研究进展与栽培研究展望[J].作物研究,2009(1):1-6.
[2]张志刚,罗孝和,罗 珊.水稻光温敏核不育系P88S的选育与应用[J].新品种推广与应用,2008,27(11):123-125.
[3]罗孝和,夏石头,罗 珊,等.两用核不育系及其超级稻研究[J],杂交水稻,2010,(25):59-63.
[4]沈志忠.近代中国水稻品种改良探析[J].江海学刊,2010,(6):164-170.
[5]Zhu Z H,Xu F,Zhang YX,et al.Arabidopsis resistance protein SNC1 activates immune responses through association with a transcriptional corepressor[J].Proceedings of the National Academy of Sciences,2010,107(31):13960-13965.
[6]Lin Y J,Zhang Q F.Optimizing the tissue culture conditions for high efficiency transformation of indica rice[J].Plant Cell Reports,2005,(23):540-547.
[7]李明浩,陈 炜,邢莉萍,等.普通小麦品种Alondra’s遗传转化体系的建立[J].植物学报,2010,(4):466-471.
[8]成 平,陈 锦,袁定阳,等.植物抗病基因snc1在超级稻父本0293中的遗传转化[J].湖南农业大学学报(自然科学版),2012,38(3):267-270.
猜你喜欢
清华金融评论(2022年4期)2022-04-13
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
现代园艺(2020年7期)2020-04-22
湖北农业科学(2019年20期)2019-12-11
农业与技术(2018年5期)2018-04-25
农家之友(2018年12期)2018-03-12
农产品加工(2017年19期)2017-11-14
中国稻米(2017年2期)2017-04-28
发明与创新(2016年7期)2016-12-18
