
杜仲黄酮生物合成途径相关基因表达差异研究
2014-01-19 06:54李铁柱杜红岩
经济林研究 2014年1期
李铁柱,杜红岩,王 璐
(1.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;2.国家林业局杜仲工程技术研究中心,河南 郑州 450003)
杜仲Eucommia ulmoidesOliv.是我国特有名贵药用树种[1-3],果实、叶片、树皮、雄蕊等组织和器官中含有丰富的苯丙素类、不饱和脂肪酸、黄酮类、环烯醚萜类、木质素类等活性成分[4]。其中杜仲黄酮具有抗氧化、降血脂、降血脂和提高免疫力等生物活性,其含量是判断杜仲药材品质的主要指标[5-9]。目前,至少有6个杜仲黄酮类化合物被分离纯化,它们是斛皮素、异斛皮素、芦丁、紫云英苷、斛皮素-3-O-α-L-吡喃阿拉伯糖基-(1→2)-8-D-葡萄糖苷和山奈酚[10-11]。杜仲的老叶中黄酮类含量最高,其它部位的含量相差不大,并且杜仲黄酮具有良好的热稳定性[12]。研究表明,10~200μg/mL浓度的杜仲黄酮有促进大鼠成骨细胞表达骨钙素(BGP)mRNA的表达水平[13],表明杜仲黄酮在基因水平促进了BGP蛋白的合成,但对骨细胞合成Ⅰ型胶原蛋白促进作用不显著[14]。杜仲黄酮还对动物油具有强的抗氧化能力[15],产生了与氨基胍类药物相似的抑制糖基化产物产生的功效[16]。杜仲黄酮中的斛皮素是主要的降血压成分,它是通过促进生物体内皮细胞释放NO舒张血压,还对动脉平滑肌细胞血管紧张素转换酶、抑制平滑肌细胞的增生及血管紧张素的生成产生稳定持久的抑制作用,来舒张血管降低血压[17-18]。
有研究证明,杜仲中的黄酮类物质主要是槲皮素和山奈酚。槲皮素(quercetin),别名栎精、槲皮黄素。分子式:C15H10O7;山奈酚,别名山奈酚,又叫山奈黄酮醇、四羟基黄酮、百蕊草素,英文名:kaempferol,分子式:C15H10O6。槲皮素和山奈酚在植物体内的合成代途径非常相似,首先,苯丙氨酸在苯丙氨酸裂解酶(PAL)和香豆酸-CoA酶(4CL)作用下,形成对香豆酰辅酶A。1分子的对香豆酰-CoA与3分子的丙二酰-CoA在查尔酮合成酶CHS的催化下,缩合生成查尔酮,再在查尔酮异构酶(CHI)的作用下,形成柚皮素,再经黄酮醇3-羟化酶作用形成二氢山奈酚。二氢山奈酚在类黄酮3’-羟化酶作用下,形成二氢槲皮素,最后在黄酮醇合成酶作用下,形成槲皮素[19-20]。二氢山奈酚也可直接通过黄酮醇合成酶的催化作用,形成山奈酚(见图1)。
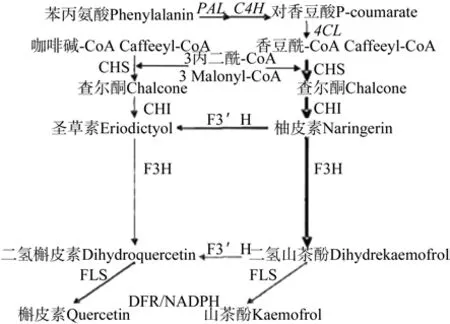
图1 槲皮素和山奈酚的合成途径Fig.1 Biosynthesis pathway of quercetin and kaempferol
1 材料与方法
分别于2010年5月28日和10月28日,在国家林业局泡桐研究开发中心采集杜仲优良品种“华仲6号”的叶片和果实,提取RNA,送至深圳华大基因公司进行转录组测序和基因功能注释[21-22]。
转录组测序得到的Unigene,利用NCBI网站上的BLAST工具,和其它物种已知的黄酮合成基因的CDS序列进行在线比较,用相似性得分比较高的已知基因确定并命名杜仲转录组中的黄酮合成相关基因。基因确定后,分析该基因(或基因家族)在杜仲叶片和果实中的表达量,推断该基因在杜仲黄酮生物合成中的地位和作用。
2 结果与分析
2.1 杜仲转录组中黄酮合成途径中被注释的Unigene基因
杜仲叶片和果实转录组数据中黄酮合成途径共涉及5个代谢位点(见图2)。代谢位点2.3.1.74注释的有Unigene10852等9条Unigene,叶片和果实中差异表达基因有4条,代谢位点5.5.1.6注释的Unigene有Unigene16591等3条,其中差异表达基因有1条;代谢位点1.14.11.9注释的Unigene有Unigene10234等24条Unigene,其中差异表达基因有13条。代谢位点1.14.13.21注释的Unigene有Unigene10190等42条,其中差异表达基因有20条;代谢位点1.14.11.23注释的Unigene有10234Unigene等49条Unigene,其中差异表达基因有30条。
2.2 杜仲黄酮合成途径相关酶基因的确定
经过进一步BLAST分析发现,Unigene19573、Unigene14862 、Unigene10852 等3条Unigene与葡萄、山茶等植物的查尔酮合酶相似性达到77%~94%,其中Unigene14862为查尔酮合酶全长cDNA;Unigene21146、Unigene7680等2条Unigene与棉花等植物种的查尔酮异构酶相似性达到73%~79%,其中Unigene21466为查尔酮异构酶全长cDNA;Unigene13947与蛇葡萄等植物种的黄烷酮3-羟化酶相似性达到89%;Unigene14810与葡萄等植物种的类黄酮3'-羟化酶相似性达到80%,为全长cDNA;Unigene3606、Unigene40118与山茶等植物种的黄酮醇合成酶相似性分别为84%、70%(见表1)。
2.3 杜仲黄酮合成相关基因的多样性及表达规律
2.3.1 EuCHS基因的多样性和表达调控规律
查尔酮合酶是黄酮合成的关键酶基因。转录组分析结果表明,注释为查尔酮合酶的Unigene14862、Unigene19573、Unigene10852 等3条Unigene在叶片和果实中有表达,分别命名为EuCHS1~EuCHS3(见表2)。

图2 KEGG数据库中的黄酮类物质生物合成途径Fig.2 The KEGG pathway of fl avonoids biosynthesis

表1 杜仲黄酮合成途径中相关酶基因和对应Unigene相似度信息Table 1 Information of similarity between gene and Unigene in flavonoids synthesis pathway

表2 杜仲查尔酮合成酶基因信息Table 2 Information of EuCHS genes
EuCHS1(Unigene14862) 和EuCHS2(Unigene19573)在叶片和果实中的表达量存在显著差异,EuCHS1在叶片中的表达量比果实高出7.88倍,而EuCHS2果实中的表达量比叶片高出6.56倍(见表3和图3)。推测杜仲叶片中的查尔酮合酶以EuCHS1为主,而果实中以EuCHS2为主。叶片EuCHS基因按照表达量由大到小的顺序排列依次为EuCHS1、EuCHS3、EuCHS2,果实中依次为EuCHS2、EuCHS1、EuCHS3。
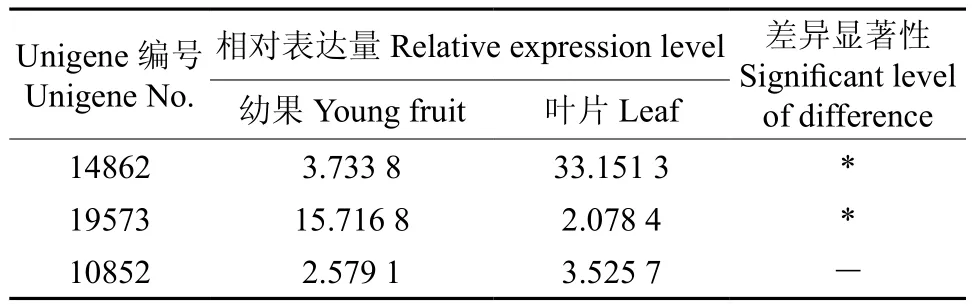
表3 查尔酮合酶相关Unigene在杜仲叶片和果实中的表达量Table 3 Expression of Unigene related with EuCHS in leaf and fruit
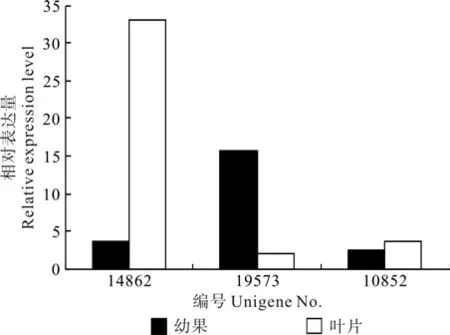
图3 查尔酮合酶相关Unigene在杜仲叶片和果实中的表达差异Fig.3 Expression difference of Unigene related with CHS in leaf and fruit
2.3.2 EuCHI基因的多样性和表达调控规律
在杜仲果实和叶片转录组数据中,注释为查尔酮异构酶的是Unigene21146,命名为EuCHI,Unigene7680为查尔酮异构酶依赖蛋白(见表4)。

表4 杜仲查尔酮异构酶基因信息Table 4 Information of EuCHI genes
查尔酮异构酶在杜仲叶片和果实中均有表达,但表达量未达到显著水平(见表5和图4),查尔酮异构酶依赖蛋白(Unigene7680)的表达量达到了显著水平,在果实中的表达量比叶片高出2.67倍。

表5 查尔酮异构酶相关Unigene在杜仲叶片和果实中的表达量Table 5 Expression of Unigene related with EuCHI in leaf and fruit

图4 查尔酮异构酶相关Unigene在杜仲叶片和果实中的表达差异Fig.4 Expression difference of Unigene related with CHI in leaf and fruit
2.3.3 EuF3H基因的多样性和表达调控规律
注释为黄烷酮3-羟化酶(F3H)的Unigene13947在叶片和果实中有表达,且表达量存在显著差异(见表6和图5),Unigene13947在果实中的表达量比叶片高出6.30倍。

表6 黄烷酮3-羟化酶相关Unigene在杜仲叶片和果实中的表达量Table 6 Expression of Unigene related with F3H in leaf and fruit

图5 黄烷酮3-羟化酶相关Unigene在杜仲叶片和果实中的表达差异Fig.5 Expression difference of Unigene related with F3H in leaf and fruit
2.3.4 EuF3'H基因的多样性和表达调控规律
注释为类黄酮3'-羟化酶的Unigene14810在叶片和果实中均有表达,但表达量无显著差异(见表7和图6)。

表7 类黄酮3'-羟化酶相关Unigene在杜仲叶片和果实中的表达量Table 7 Expression of Unigene related with F3’H in leaf and fruit
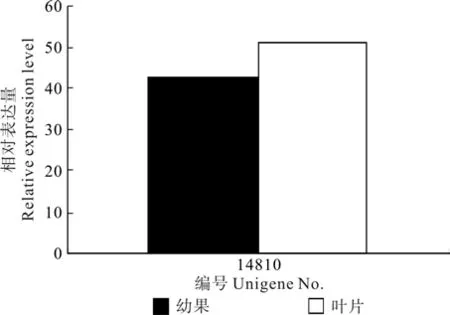
图6 类黄酮3’-羟化酶相关Unigene在杜仲叶片和果实中的表达差异Fig.6 Expression difference of Unigene related with F3’H in leaf and fruit
2.3.5 EuFLS基因的多样性和表达调控规律
黄酮醇合成酶是黄酮合成途径的最后1个酶,也是限速酶。转录组分析表明,注释为黄酮 醇 合 成 酶的 Unigene35503、Unigene3603、Unigene40118、Unigene43370等4条Unigene,分别命名为EuFLS1~EuFLS4(见表8)。

表8 黄酮醇合成酶相关Unigene在杜仲叶片和果实中的表达量Table 8 Expression of Unigene related with FLS in leaf and fruit
EuFLS1~EuFLS4在叶片和果实中均有表达,且表达量均达到显著水平(见表9),其中EuFLS3和EuFLS4在叶片中的表达量比果实分别高出12.98倍和44.24倍,而EuFLS1和EuFLS2在果实中的表达量比叶片分别高出15.38倍和17.65倍(见图7)。推测杜仲叶片中的黄酮醇合成酶以EuFLS3和EuFLS4为主,而在杜仲果实中以EuFLS1和EuFLS2为主。
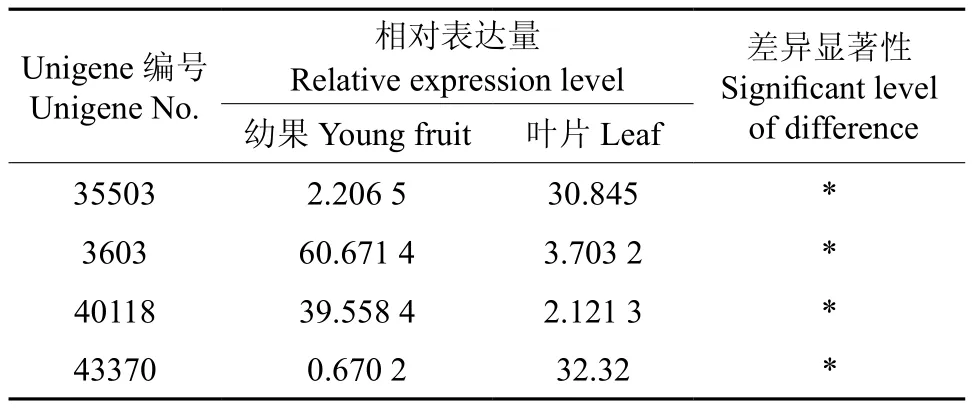
表9 黄酮醇合成酶相关Unigene在杜仲叶片和果实中的表达量Table 9 Expression of Unigene related with FLS in leaf and fruit
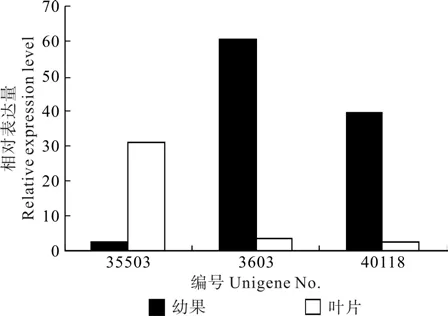
图7 黄酮醇合成酶相关Unigene在杜仲叶片和果实中的表达差异Fig.7 Expression difference of Unigene related with FLS in leaf and young fruit
3 结论与讨论
该研究揭示了杜仲黄酮(槲皮素和山奈酚)合成途径重要基因遗传多样性。在幼果和成熟叶片中获得黄酮合成相关酶11条,其中差异表达基因8条;查尔酮合酶(CHS)获得3个家族成员(EuCHS1~EuCHS3),EuCHS1和EuCHS2在叶片和幼果中表达量存在显著差异,EuCHS1在叶片中的表达量比果实高7.88倍,而EuCHS2果实表达量比叶片高6.56倍,杜仲叶片以EuCHS1为主,而果实以EuCHS2为主;杜仲查尔酮异构酶EuCHI在叶片和幼果均有表达,但表达量未达到显著水平,获得了查尔酮异构酶-like protein1条,在叶片和幼果中表达量达到了显著水平,且果实比叶片高2.67倍;杜仲黄烷酮3-羟化酶EuF3H在叶片和果实均有表达,且表达量存在显著差异,果实比叶片高6.30倍;杜仲类黄酮3'-羟化酶EuF3′H在叶片和果实均有表达,但表达量无显著差异,杜仲黄酮醇合成酶(FLS)基因获得了4(EuFLS1~EuFLS4)条家族成员,EuFLS1~EuFLS4在叶片和幼果均有表达,且均达到显著水平,其中EuFLS3和EuFLS4在叶片中的表达量比幼果分别高出12.98倍和44.24倍,而EuFLS1和EuFLS2在果实中的表达量比叶片分别高出15.38倍和17.65倍,故叶片以EuFLS3和EuFLS4为主,而果实以EuFLS1和EuFLS2为主。
槲皮素和山奈酚在植物体内的合成代途径非常相似,首先,苯丙氨酸在苯丙氨酸裂解酶(PAL)和香豆酸-CoA酶(4CL)作用下,形成对香豆酰辅酶A。1分子对香豆酰-CoA的与3分子的丙二酰-CoA在查尔酮合成酶CHS的催化下,缩合生成查尔酮,再在查尔酮异构酶(CHI)的作用下,形成柚皮素,再经黄酮醇3-羟化酶作用形成二氢山奈酚。二氢山奈酚在类黄酮3’-羟化酶作用下,形成二氢槲皮素,最后在黄酮醇合成酶作用下,形成槲皮素。
槲皮素和山奈酚合成途径中的基因中关键酶的遗传多样性有差异,如查尔酮合酶(CHS)和杜仲黄酮醇合成酶(FLS)的遗传多样性较高,在杜仲中分别获得了3条和4条家族成员,而杜仲查尔酮异构酶(CHI)和黄烷酮3-羟化酶F3H的基因遗传多样性较低,具有很高的保守性,在杜仲果实和叶片中均获得了1条基因。在果实和叶片中查尔酮合酶(CHS)和杜仲黄酮醇合成酶(FLS)的基因表达也较复杂,杜仲叶片以EuCHS1为主,而果实以EuCHS2为主;杜仲黄酮醇合成酶(FLS)基因家族中,EuFLS3和EuFLS4在叶片中的表达量比幼果分别高出12.98倍和44.24倍,而EuFLS1和EuFLS2在果实中的表达量比叶片分别高出15.38倍和17.65倍,故叶片以EuFLS3和EuFLS4为主,而果实以EuFLS1和EuFLS2为主。故需要进一步探究杜仲查尔酮合酶和黄酮醇合成酶的遗传多样性与槲皮素和山奈酚以及其它活性成分的积累的相关性和控制机理。
[1] 李时珍.本草纲目[M].北京:北京出版社,1995:1590-1596.
[2] 杜红岩.杜仲活性成分与药理研究的新进展[J].经济林研究,2003,21(2): 58-61,82.
[3] 薛 萍.杜仲研究现状与发展前景[J].经济林研究,1995,13(3): 56-58.
[4] 薛程远,曲范仙,刘 辉,等.杜仲叶乙醇提取物对小鼠免疫功能的影响[J].甘肃中医学院学报,1998,15(3):50-52.
[5] 李家实,阎玉凝.杜仲皮与叶化学成分初步研究[J].中药通报,1986,(8):41-42.
[6] 张康健,王 蓝,张凤云,等.杜仲叶与皮有效成分含量的比较研究[J].西北林学院学报,1996,(2):44-48.
[7] 黎继烈,刘宗敏,钟海雁,等.金柑总黄酮对小鼠胃肠吸收功能的影响[J].中南林业科技大学学报,2007,27(2):79-82.
[8] 杜红岩,李 钦,杜兰英,等.杜仲雄花茶营养成分的测定分析[J].中南林业科技大学学报,2007,27(6):88-91.
[9] 黎继烈,王 卫,曾超珍,等.金橘黄酮提取物对大鼠降血脂功能的影响[J].中南林业科技大学学报,2008,28(5):89-92.
[10] Hye Y K,Byung H M,Hak J L,et al.Flavonol glycosides from the leaves ofEucommia ulmoidesOlive with glycation inhibitory activity[J].J Ethnopharm-Acol,200,93(2):227-230.
[11] 成 军,赵玉英,崔育新,等.杜仲叶黄酮类化合物的研究[J].中国中药杂志,2000,25(5):284-287.
[12] 张兰萍,马 龙,张 斌,等.杜仲黄酮类化合物的研究进展[J].食品工业科技,2009,30(3):359-363.
[13] 白喜婷,朱文学,罗 磊,等.杜仲雄花及花茶中总黄酮含量的测定[J].食品科技,2008,(4):186-188.
[14] 李三华,刘坤祥,莫宁萍,等.RT-PCR法检测杜仲总黄酮对大鼠成骨细胞骨钙素表达的影响[J].遵义医学院学报,2011,34(3): 223-225.
[15] 李三华,何志全,陈全利,等.杜仲总黄酮对成骨细胞增殖及Ⅰ型胶原蛋白表达的影响[J].西北药学杂志,2011,26(4):272-274.
[16] 吴 春,胡小妹,陈琳琳,等.杜仲黄酮的提取及抗氧化活性研究[J].哈尔滨商业大学学报:自然科学版,2004,20(5):509-511.
[17] Kim H Y,Moon B H,Lee H J,et al.Flavonol glycosides from the leaves ofEucommia ulmoidsOlive with glycation inhibitory activity [J].J Ethnopharm-Acol,2004,93(2):227-230.
[18] Chan E C,Pannangpetch P,Woodman O L.Relaxation to fl avones and fl avonols in rat isolated thoracicaorta: mechanism of action and structure-activity relationships [J].J Cardiovasc Pharmacol,2000,35(2): 326-333.
[19] 孙 涓,余世春.槲皮素的研究进展[J].现代中药研究与实践,2011,(3):85-88.
[20] 李云峰,郭长江.槲皮素代谢的研究进展[J].生理科学进展,2002,(1): 53-55.
[21] 李铁柱,杜红岩,刘慧敏,等.杜仲果实和叶片转录组数据组装及基因功能注释[J].中南林业科技大学学报,2012,32(11):122-130.
[22] 李铁柱,杜红岩,刘慧敏,等.杜仲幼果和成熟果实转录组数据组装及基因功能注释[J].中南林业科技大学学报,2012,32(10): 9-17.
猜你喜欢
中华实用诊断与治疗杂志(2022年2期)2022-09-02
河北果树(2021年4期)2021-12-02
热带作物学报(2020年9期)2020-10-29
中国现代医生(2020年12期)2020-07-04
祝您健康(2019年7期)2019-07-12
天然产物研究与开发(2018年9期)2018-10-08
天然产物研究与开发(2018年8期)2018-09-10
中成药(2017年9期)2017-12-19
江苏农业科学(2015年10期)2015-12-23
医学研究杂志(2015年8期)2015-06-22
