
珠江口磨刀门轮虫群落结构特征与水质生态学评价
2014-02-28 08:47王庆侯磊陈实吴志辉杨宇峰
生态环境学报 2014年5期
王庆,侯磊,陈实,吴志辉,杨宇峰*
1. 暨南大学水生生物研究所,广东 广州 510632;2. 水体富营养化与赤潮防治广东省教育厅重点实验室,广东 广州 510632;3. 北京师范大学-香港浸会大学联合国际学院,广东 珠海 519000
珠江口磨刀门轮虫群落结构特征与水质生态学评价
王庆1,2,侯磊1,2,陈实1,2,吴志辉3,杨宇峰1,2*
1. 暨南大学水生生物研究所,广东 广州 510632;2. 水体富营养化与赤潮防治广东省教育厅重点实验室,广东 广州 510632;3. 北京师范大学-香港浸会大学联合国际学院,广东 珠海 519000
轮虫是浮游动物群落中的重要组成部分,对水质变化反应敏感,调查轮虫群落的种类组成和丰度变化,可以获知水质变化和水体的营养状况,并可作为水质评价依据。于2009年7月至2010年1月,对珠江口磨刀门3个站点(S1滞水区、S2河道和S3水流交汇区)轮虫群落结构特征进行每月两次的调查,同时测定了水温、盐度、透明度、叶绿素、总氮、总磷等理化指标,利用非生物环境因子对水体的营养状况进行了TLIc加权综合营养状态指数评价,并通过轮虫种类组成、E/O指数、QB/T指数和生物多样性指数对水质进行了生态学评价。调查期间,水温12~29 ℃,盐度0.11~7.9 PSU,叶绿素a浓度0.89-9.56 μg·L-1,叶绿素a与水温呈显著正相关(p<0.0001),与盐度呈显著负相关(p<0.01)。磨刀门水域3个采样站点TLIc处于50~60之间,表明其水质均达到轻度富营养水平, 富营养程度按S1、S2、S3依次递减。调查期间,共记录轮虫56种,S1站点轮虫种类最多,其中污染指示轮虫42种。污染指示种占轮虫总数比例表明,珠江口磨刀门达到了富营养化水平,与利用非生物环境因子作出综合营养状态指数TLIc的评价结果一致。磨刀门的优势种为广布多肢轮虫(Polyarthra vulgaris)、螺形龟甲轮虫(Keratella cochlearis)、热带龟甲轮虫(Keratella tropica)和暗小异尾轮虫(Trichocerca pusilla)。调查期间,轮虫丰度在S1、S2、S3变化范围分别为0~199 ind·L-1、1~54 ind·L-1和0~48 ind·L-1,平均丰度分别为31 ± 54 ind·L-1、13 ± 15 ind·L-1和11 ± 13 ind·L-1。丰水期的8月下旬轮虫丰度达到最大值;枯水期丰度均较低。轮虫丰度与水温、总氮、叶绿素显著正相关(p<0.05),与盐度显著负相关(p<0.05)。磨刀门河口理化因子变化剧烈,轮虫多样性指数波动范围较大,反映出河口水域轮虫群落结构的不稳定性。冗余分析(RDA)表明水温、盐度和叶绿素是影响轮虫群落结构季节变化的主要因子。根据水质生态学评价指标,磨刀门水体已达轻度富营养水平,与非生物环境因子营养状态综合评价结果一致。本研究表明,评价一个水体的污染程度,应将生态学指标与理化指标结合起来综合评价,可获得更为真实的评价结果。
珠江口;轮虫;水质评价;生态学评价;富营养化
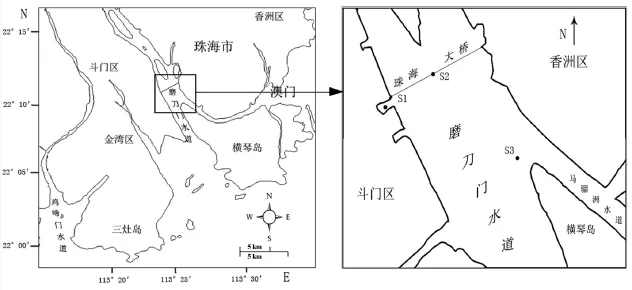
图1 珠江口磨刀门水道及采样站位图Fig.1 Map shown three sampling sites (S1, S2 and S3) in Modaomen of the Pearl River Estuary
1 材料与方法
1.1 采样时间与采样点布置
2009年7月至2010年1月,在珠江口磨刀门水道进行每月两次的调查和采样。本研究根据地形、水质状况和流速,于磨刀门水道设置3个站点,分别在珠海大桥西滞水区(S1站点)、珠海大桥中部(S2站点),磨刀门水道下游与马骝洲水道交汇区(S3站点),站点设置见图1。
1.2 样品采集与分析
水温、盐度、电导率、pH用YSI-556水质分析仪(美国)现场测定,将水质仪探头放入水面下约0.5 m处,缓慢旋转探头,待测定值在30s内未有显著变化即读取;用萨氏盘在背光处测定透明度(SD)。于每个站点采集表层0.5 m处的水样, 室内分平行样按国家水质标准方法GB3838-2002测定TN、TP,叶绿素a用丙酮分光光度法进行测定(Lorenzen, 1967),轮虫定量样品用20 μm浮游生物网过滤浓缩10 L水样,收集至50 ml聚乙烯瓶中,甲醛溶液固定(最终体积分数为4%)(章宗涉和黄祥飞, 1991)。定性样品分固定样品和活体样品,用38 μm的浮游生物网垂直和水平拖取,固定样品用甲醛溶液固定,活体样品带回实验室用于软体轮虫的镜检(章宗涉和黄祥飞, 1991)。种类鉴定依据Koste 分类系统(Koste, 1978)和《中国淡水轮虫志》(王家楫,1961),定量样品全部计数。
1.3 数据处理
数据统计分析采用SPSS 16.0 和Canoco 4.5软件,对水体的富营养化评价采用加权综合Carlson指数TLIc(金相灿等,1991),采用Shannon-Wiener和Margalef多样性指数和Pielou物种均匀度指数评价轮虫群落多样性,不同水体出现的轮虫种类采用Czekanowsili(Sc)相似性系数,轮虫物种优势度采用优势度(Y)(王云龙等,2005)。运用轮虫物种E/O 指数(尚玉昌,2002)和公式QB/T指数(Sládeček, 1983)评价水体营养状态。
1)TLIc加权综合营养状态指数公式:
TLIc = ∑WjTLI(j)
式中:TLI(∑)—综合营养状态指数;Wj—第j种参数的营养状态指数的相关权重;TLI(j)—代表第j种参数的营养状态指数。
2)Shannon-Wiener多样性指数(H') :
H' = -∑(Pi)(log2Pi)
3)Margalef多样性指数(d):
d = (S-1)/lnN
4)Pielou物种均匀度指数(J):
J = H'/lnS
式中S为种类数,Pi为某种个体数占总个体数的比例,N为样品中所有种类总个体数。
5)Czekanowsili相似性系数(Sc):
Sc = 100%×2C/(A+B)
式中,C为两水体共有种类数,A、B分别为水体A和B的种类数。
6)优势度(Y):
Y = (ni/N)×fi
式中,ni为第i种的个体数,fi为第i种出现的频率,N为所有种类出现的总个体数. Y > 0.02 为优势种[16]。
7)E/O指数: 按E/O = E (富营养型指示种数) /O(贫营养型指示种数)计算。E/O指数 < 0.5为贫营养型; 0.5~1.5为中营养型; 1.5~5.0为富营养型;>5.0为超富营养型(尚玉昌,2002)。
8)QB/T指数:按QB/T= B(臂尾轮虫属的种数)/T(异尾轮虫属的种数)计算。QB/T指数< 1为贫营养型; 1~2为中营养型; > 2为富营养型(Sládeček,
1983)。
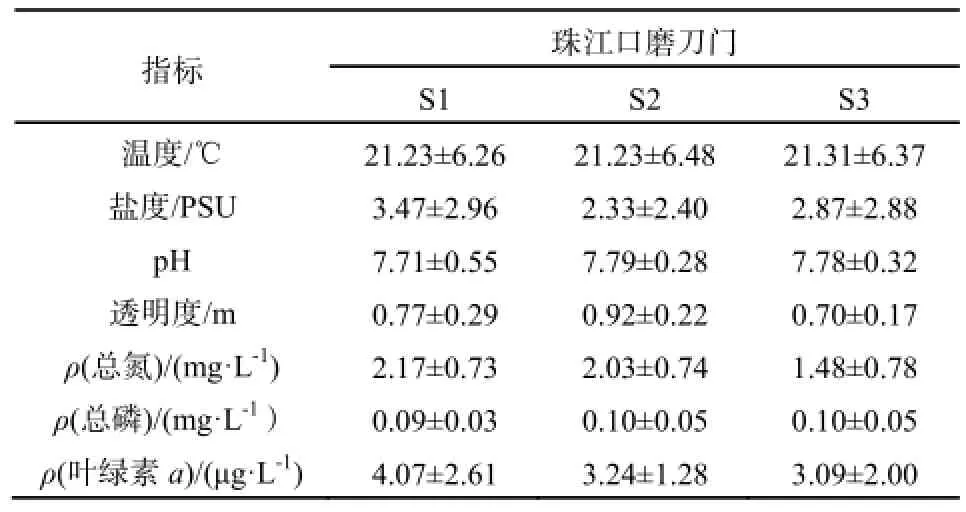
表1 珠江口磨刀门水质指标Table 1 Water quality indicators in Modaomen of the Pearl River Estuary
2 结果
2.1 理化因子的季节变化
珠江口磨刀门水道,是典型的河口区域,温度、盐度等理化因子季节变化较为剧烈(表1)。调查期间磨刀门河口最高水温29 ℃,最低水温12 ℃,平均温度21.2 ℃。磨刀门河口接近外海,盐度变化显著,最高盐度7.9记录于11月上旬S1站点,最低值为S2和S3的0.11(8月)。叶绿素a浓度1月下旬最低(S2,0.89 μg·L-1),9月上旬最高(S1,9.56 μg·L-1)。S1站点位于滞水区,较适于浮游植物生长,叶绿素a平均值为4.07 ± 2.61 μg·L-1,S2和S3站点水流较急,不利于浮游植物生长,叶绿素a平均值分别为3.24 ± 1.28 μg·L-1和3.09 ± 2.00 μg·L-1。叶绿素a与温度呈显著正相关(r=0.605,p<0.0001),与盐度呈显著负相关(r= -0.449, p=0.004)(图2)。
2.2 水体富营养化评价

图2 磨刀门水域水温、盐度与叶绿素a的季节变化Fig.2 Seasonal variations of water temperature, salinity and chlorophyll a in Modaomen Estuary
本文采用修正的Carlson营养状态指数(TLI),包括SD、TN、TP和Chl a在内的相关加权综合营养状态指数评价河口营养状态。珠江口磨刀门水域3个采样站点的TLIc都处于50-60之间(表2),表明其水质均达到轻度富营养水平(金相灿等,1991)。富营养程度按S1、S2、S3依次递减,磨刀门水域 S3采样点,由于海水稀释作用,其TLIc值为50.42,为研究调查站点的最低值。
2.3 轮虫群落结构特征
2.3.1 种类组成
调查期间,磨刀门水域共发现轮虫56种,其中S1站点47种,S2站点35种,S3站点轮虫33种(表3)。3个站点共发现轮虫污染指示种42种,其中寡污性13种,占31.0%;o-β和β-o 9种,占21.4%;β-α中污性5种,占12.0%;β中污性15种,占35.7%。从不同等级轮虫种类占总数来看,珠江口磨刀门达到了富营养化水平,与利用非生物环境因子作出综合营养状态指数TLIc的评价结果一致。
利用Czekanowsili相似性系数评价,磨刀门水域S1与S2和S3相似度分别达到72.3%和73.2%;S2和S3相似度为72.5%。3个站点出现的种类均有很高的相似度,多数种类在3个站点同时出现。
2.3.2 丰度变化
磨刀门水域3个采样站点轮虫丰度较低,S1、S2、S3调查期间变化范围分别是0~199 ind·L-1、1~54 ind·L-1和0~48 ind·L-1,平均丰度分别为31 ± 54 ind·L-1、13 ± 15 ind·L-1和11 ± 13 ind·L-1。S1丰度最高,其次是S2站点,S3的轮虫丰度最低。丰水期的8月下旬轮虫丰度达到最大值;枯水期丰度均较低,
在枯水期的11月下旬,三个站点同时出现丰度的第二次高峰,轮虫丰度的最低值均出现在11月上旬(图3)。珠江口磨刀门轮虫丰度受多种理化因子影响,其中与TLIc相关性最大,其次是叶绿素a;轮虫丰度与温度呈现显著正相关(p<0.05) ,与盐度呈现显著负相关(p<0.05)(表4)。

表2 珠江口磨刀门营养状态综合评价结果Table 2 Results of the comprehensive evaluation of trophic state in Modaomen of the Pearl River Estuary
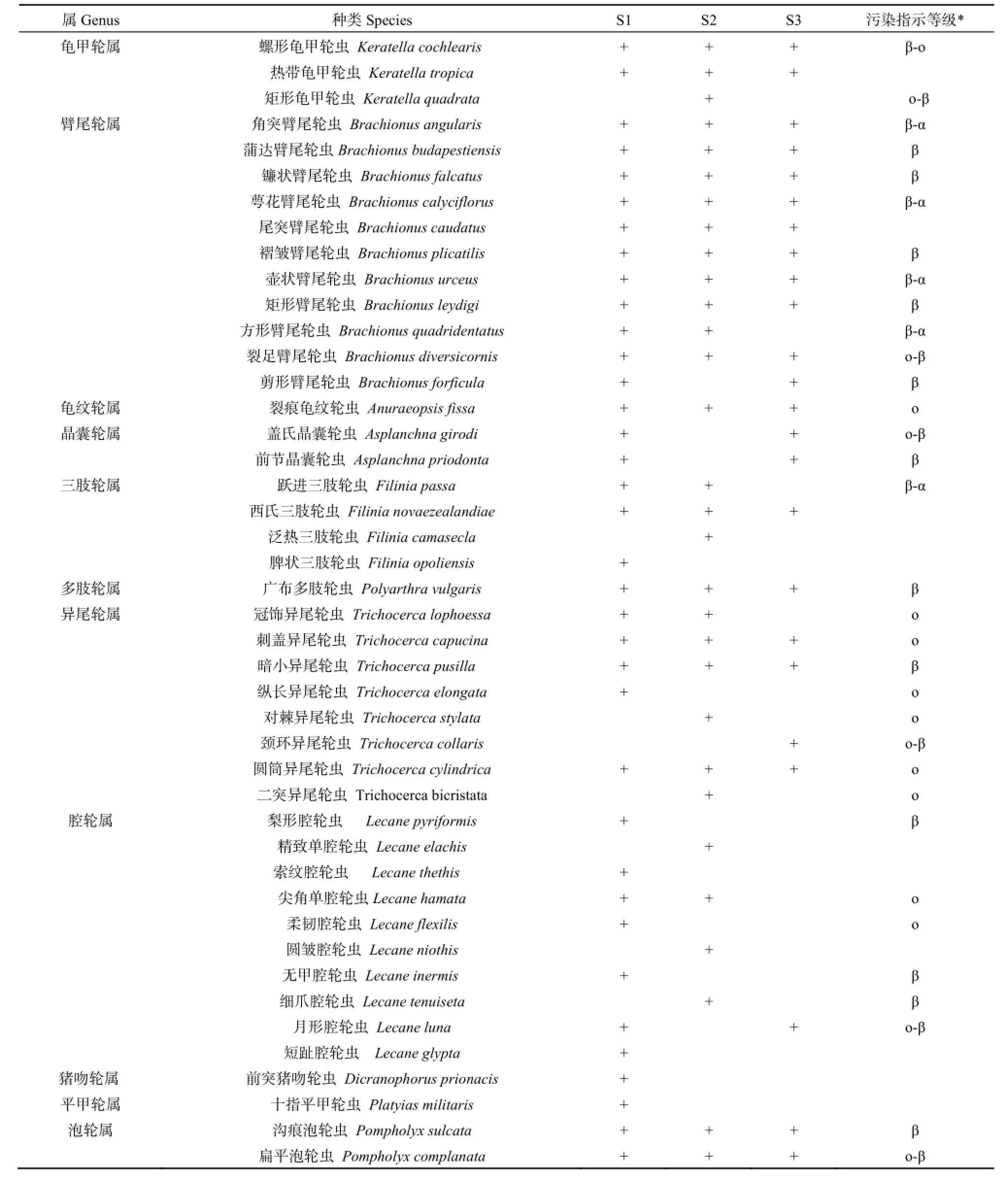
表3 磨刀门轮虫的种类组成Table 3 Species composition of rotifer in Modaomen Estuary

续表3

表4 轮虫丰度与各项理化指标相关性分析Table 4 Correlation analysis between rotifer abundances and physicochemical indicators
2.3.3 优势种
广布多肢轮虫(Polyarthra vulgaris)是磨刀门S1、S2和S3的第一优势种,其优势度分别是0.248、0.333、0.245。S1的第二优势种是螺形龟甲轮虫(Keratella cochlearis),优势度为0.090。热带龟甲轮虫(K. tropica)是S2的第二优势种,其优势度为0.078,S3的第二优势种为暗小异尾轮虫(Trichocerca pusilla),优势度为0.061。3个站点广布多肢轮虫的丰度最大值均出现在8月下旬,在9月下旬又出现了另一高值,而在秋冬枯水季节丰度较低(图3)。
3个采样点优势种和季节变化较为相似。异尾轮虫一般只出现在夏秋季节;轮虫的第一次高峰出现在2009年8月上旬,泡轮属、多肢轮属和臂尾轮属分别占S1、S2和S3三个站点轮虫总量的53.6%,46.7%和37.4%,分别是各站点该月上旬的优势属。轮虫的第二次高峰出现在2009年11月下旬,龟甲轮虫占S1轮虫总量的21.3%,多肢轮虫分别占S2和S3的28.5%和22.6%,是两个站点的优势属(图4、图5和图6)。
2.3.4 物种多样性指数
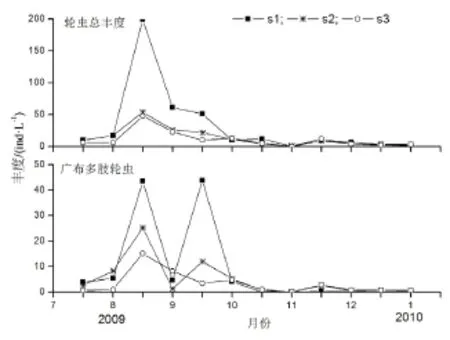
图3 磨刀门轮虫丰度的季节变化Fig.3 Seasonal variations of rotifer abundances in Modaomen Estuary
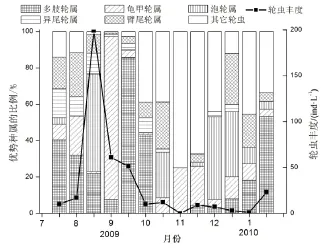
图4 磨刀门S1站点优势种属的季节变化Fig.4 Seasonal changes in the percentage of dominant rotifers at S1 in Modaomen Estuary
磨刀门水域轮虫种类数季节波动较大,3个站点的种类数在8月下旬同时达到最大值,在11月上旬种类数最少。轮虫物种多样性指数 H'值在
1.05~3.63之间波动,物种丰富度指数d值在0.85~4.40之间波动, 均匀度指数J值在0.27~0.95之间波动,3个站点之间差异显著,S1各指数均值最小,S3站点各指数均值最大(表5)。河口区域理化因子变化剧烈,轮虫物种多样性指数波动较大。种类数与轮虫丰度呈现显著正相关(r=0.506, p=0.001);Pielou物种均匀度指数与轮虫丰度呈现显著负相关(r=-0.482,p=0.002)。种类数、物种多样性指数与多种理化因子具有相关性(表6)。

图5 磨刀门S2站点优势种属的季节变化Fig.5 Seasonal changes in the percentage of dominant rotifers at S2 in Modaomen Estuary
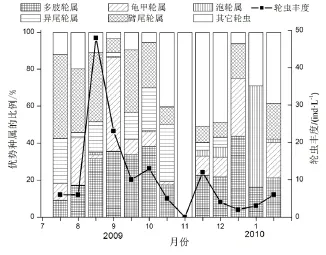
图6 磨刀门S3站点优势种属的季节变化Fig.6 Seasonal changes in the percentage of dominant rotifers at S3 in Modaomen Estuary
2.3.5 轮虫的 E/O指数和QB/T指数值
磨刀门水域轮虫的E/O值为3.19 ± 1.89,在0~8之间波动;轮虫的QB/T值为1.84 ± 1.11,在0~5之间波动。E/O指数和QB/T均响应了河口复杂的理化环境,波动较为剧烈,S2和S3两个站点之间显著相关(r=0.398,p=0.044)。通过E/O值可以判定磨刀门水体已经达到了轻度富营养状态,通过QB/T值可确定磨刀门水体水质在中营养与轻度富营养之间(表7)。综合两个指数,通过生态学的水质评价,磨刀门水质为轻度富营养型,与运用理化指标评价的水体综合营养状态指数TLIc结果一致。

表5 磨刀门轮虫物种多样性、丰富度及均匀度指数Table 5 Indices of species diversity, abundance and evenness of rotifer inModaomen Estuary

表6 种类数、多样性指数与理化因子相关性分析Table 6 Correlation analysis between species number, diversity index and physicochemical indicators

表7 磨刀门水域3个采样点的轮虫 E/O值和QB/T值Table 7 E/O index and QB/Tindex of rotifer at three sampling sites in Modaomen Estuary
2.4 样品的RDA排序
RDA是一种直接梯度分析方法, 能从统计学的角度评价一个或一组变量与另一组多变量之间的关系。对磨刀门轮虫种类进行的DCA分析显示排序轴最大梯度长度为2.313, 表明磨刀门水域轮虫群落对生态梯度的响应是线性的,因此利用基于线性模型的排序方法(RDA)来分析环境因子对轮虫群落的影响是比较适宜的。RDA冗余分析是在环境因子特征变量构成的空间上,对环境变量和轮虫种类排序作图,实现轮虫种类和环境因子的对应排序。对主要轮虫种类的丰度在主轴上排序,可以反映这些种类和环境因子之间的对应关系。对种类数据按出现频度和相对丰度进行了筛选,RDA排序分析的统
计信息如表8、表9。
RDA排序的全部特征值解释了轮虫群落变异程度的43.4%,前两轴共解释了轮虫变异程度的37.5%。水温、盐度、叶绿素a是影响轮虫时空分布的最主要因素,对轮虫分布影响最大。水体透明度、总氮、溶解氧对其分布也有一定影响。从轮虫种类分布来看(图7),角突臂尾轮虫(Brachionus angularis)、矩形臂尾轮虫(B. leydigi)和镰状臂尾轮虫(B. falcatus)受温度影响较大;螺形龟甲轮虫和热带龟甲轮虫受叶绿素a浓度的影响较大;磨刀门水域的绝大多数轮虫分布与盐度呈现明显的负相关关系。
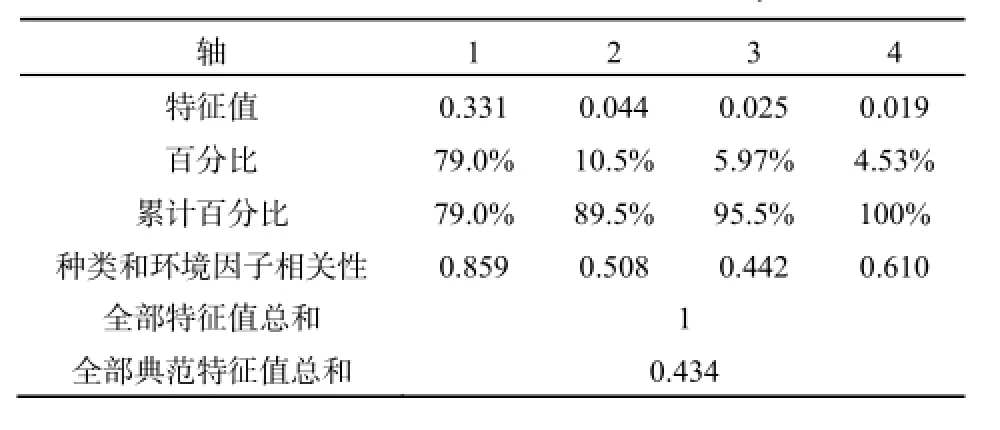
表8 磨刀门水域轮虫种类和环境因子之间RDA分析的统计信息Table 8 Statistical Information of RDA between rotifer species and environmental factors in Modaomen Estuary
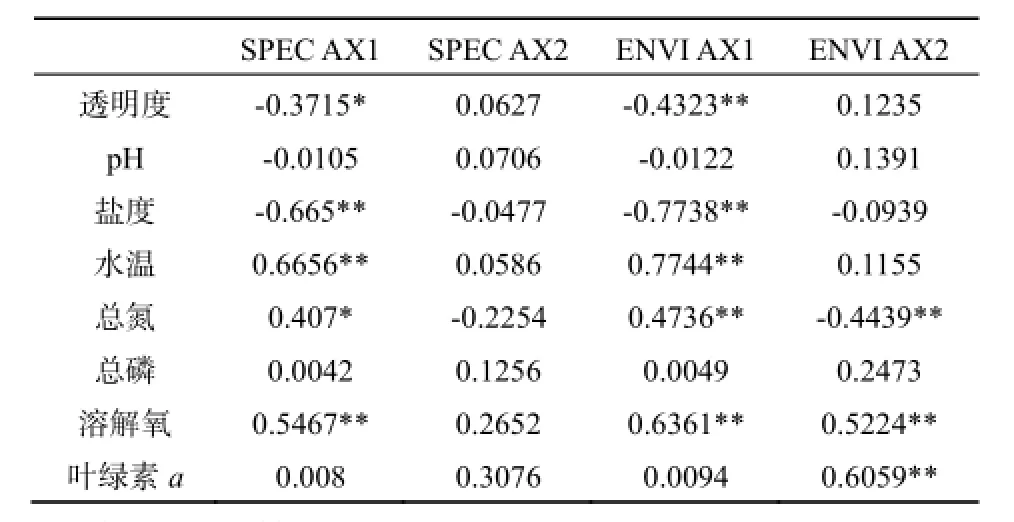
表9 前两个排序轴与环境因子间的相关系数Table 9 Correlation coefficient between the first two ordination axes and environmental factors
3 讨论
3.1 轮虫的种类组成
轮虫是水生态系统中的重要指示生物(Sládeček, 1983; Gutkowska等, 2013)。磨刀门河口既有寡营养型水体的常见种,也包括富营养型水体的指示种。有关轮虫指示种,虽然不同学者观点不同,但对多数指示种类的看法是一致的(Sládeček, 1983),富营养指示种中的臂尾轮属、裂痕龟纹轮虫、沟痕泡轮虫、扁平泡轮虫、圆筒异尾轮虫、长三肢轮虫、暗小异尾轮虫、螺形龟甲轮虫、矩形龟甲轮虫在磨刀门水体中都有出现。其中臂尾轮属有9种,种类数上占绝对优势,且螺形龟甲轮虫、暗小异尾轮虫都是优势种,说明磨刀门水体已受一定程度污染,与非生物环境因子评价一致。
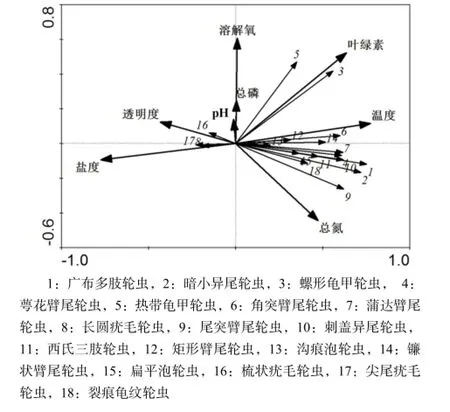
图7 磨刀门轮虫种类与环境因子之间的RDA排序图Fig.7 Ordination biplots between rotifer species and environmental factors in Modaomen Estuary
轮虫种类主要是广生性的,例如广布多肢轮虫、螺形龟甲轮虫等。广布多肢轮虫是磨刀门水域的第一优势种,它能在所有的淡水环境中存活,通常在夏天丰度达到最高值,冬天其个体和附肢生长到最大状态(Sládeček, 1983)。广布多肢轮虫在水体中移动速度要快于其它轮虫,这样不仅有利于捕食,而且还能逃避敌害(陈立斌等,2010),相对于其它种类,其更容易适应河口这种流水环境,且广布多肢轮虫和褶皱臂尾轮虫一样,也能够耐受一定的盐度(Onwudinjo等, 1994)。龟甲轮虫是浮游轮虫分布最广泛的种类之一,它们的食性很广,有机碎屑、鞭毛虫和藻类都是螺形龟甲轮虫的食物来源(Gilbert 和 Bogdan, 1981)。Onwudinjo等(1994)研究了尼日利亚的河口,发现热带龟甲轮虫和螺形龟甲轮虫在盐度16~18时也能生存,属于能耐受较高盐度的轮虫种类。
磨刀门臂尾轮虫种类最多,主要以藻类和有机颗粒为食物来源;耐受污水,属于污水指示种类,适应能力较强,能够应对时空上剧烈变化的严酷环境(Sousa等, 2008)。在极端环境下,捕食和竞争的压力都会减小,耐受力强的物种更易于存活(Herbst, 2001)。褶皱臂尾轮虫对盐度有一定耐受性,通常认为是海水种类,但在磨刀门水域其优势度很低,只在枯水期的下游站点检出。在马来西亚靠近南中国海的河口区域研究发现,褶皱臂尾轮虫在盐湖能耐受高盐度,但在河口这样的环境里,盐度波动剧烈,其丰度却很低 (Green, 1995)。
3.2 环境因子对轮虫时空分布的影响
已有的研究表明,温度是影响轮虫密度变化的主要环境因子(Galkovskaja, 1987)。盐度的变化是影响河口轮虫丰度的重要因素(Onwudinjo等, 1994;Park 和 Marshall, 2000a;Silva等, 2009;Medeiros等, 2010)。RDA分析显示,盐度是影响磨刀门水域轮虫群落结构时空变化的最主要因素,轮虫的丰度、种类数与盐度呈显著的负相关,这与Park和Marshall (2000a) 的研究结果一致。磨刀门3个采样站点的轮虫丰度最高值出现在8月下旬,且丰水期的丰度明显高于枯水期。种类数与丰度变化一致,轮虫丰度与种类数呈显著正相关(r=0.504,p=0.001),丰水期种类数最高。Rougier等(2005) 认为,在丰水期,由于强大的径流量,阻碍了海水回流,河口区域主要受径流的影响,很多淡水种类被冲刷到河口,种类丰富。同时,由于上游河道大量淡水的注入,使河口区域盐度降低,更适合轮虫的生长和繁殖,其丰度达到高峰(Silva 等, 2009)。
RDA分析还表明,温度和叶绿素a显著影响轮虫的季节变化,水温是影响轮虫季节分布的重要因素。黄祥飞等(1985) 研究了温度对轮虫卵发育的影响,随温度上升, 轮虫卵的发育时间缩短, 在丰富的食物供给条件下,种群周转加快, 导致种群密度迅速增加。磨刀门水域的轮虫数量峰值一般出现在20℃以上,广布多肢轮虫是磨刀门的第一优势种,研究表明,其卵在25℃的发育时间不足10℃时的三分之一(黄祥飞等,1985)。
叶绿素a是浮游植物丰度的主要指标,浮游藻类是轮虫的主要食物来源,磨刀门水域叶绿素a与轮虫丰度呈显著正相关(r=0.412, p=0.009)。在亚马逊地区Mossoró河口,叶绿素含量与轮虫丰度变化显著相关,在丰水期达到高峰(Silva等, 2009),与磨刀门河口的研究结果一致。丰水期,河口区域水流扰动剧烈,有机碎屑的浓度升高(Rougier等, 2005),浮游植物、细菌和原生动物能够大量繁殖,同时它们又是轮虫的主要食物来源(Arndt, 1993),为轮虫的增殖创造了条件。
河口是河流与海洋交汇的区域,易受水动力学影响,不同区域具有明显的生态梯度,因此轮虫的分布具有一定程度的空间异质性,S1、S2和S3站点同处于河口流域,轮虫丰度季节波动一致,但轮虫密度却有差异,S1水流相对和缓,很多渔民在这里生活,水体富营养程度较高,轮虫食物充足,丰度也最高。Park和 Marshall (2000b) 认为,水体富营养化程度与轮虫丰度有关,通常富营养状态的水体轮虫食物充足,更适于生长繁殖。磨刀门S3站点离外海最近,受水流和潮汐影响较大,盐度波动剧烈,轮虫丰度处于较低水平。
磨刀门轮虫在枯水期也出现了一个峰值,10月底至11月初,珠海市沿海水域暴发了一次大规模旋沟藻赤潮,11月下旬,磨刀门轮虫的丰度出现高值,可能因为浮游植物的大量繁殖,为轮虫提供了丰富的食物。在异养鞭毛藻赤潮中,轮虫是其主要的摄食者,赤潮的发生与轮虫的丰度紧密联系在一起(Heinbokel等, 1988)。
王庆和杨宇峰(2007)对珠江广州河段轮虫群落结构进行了较系统的调查研究,发现轮虫年平均丰度达到1272 ind·L-1。珠江口磨刀门轮虫的平均丰度远低于珠江广州河段,且珠江广州河段轮虫丰度高峰出现在枯水期前期,与磨刀门水域的研究结果相反。珠江广州河段理化因子比较稳定,主要受径流量影响,在丰水期,珠江径流量大,轮虫丰度较低;在枯水期前期,径流量减少,流速较慢,水温和盐度均适宜,轮虫丰度达到高峰(王庆和杨宇峰,2007)。磨刀门水域受潮汐的影响大,决定轮虫丰度波动的首要因子是盐度,丰水期抑制了涨潮的影响,水体处于淡水环境,适宜轮虫生长和繁殖;在枯水期,受海水潮汐影响显著,水体盐度升高,轮虫丰度迅速下降。在Chesapeake河口,也发现类似的研究结果(Park 和 Marshall, 2000a)。
3.3 轮虫与水质营养状况的关系
黄祥飞等(1985)认为多样性指数能够反映水体的污染程度,多用来评价水环境质量。多样性指数与水质关系复杂,与水体类型、计数方法及鉴定种类的水平等诸多因素都有关系(雷安平等,2003)。因此,在评价水体营养状态时,可以将多样性指数作为参考目标。在河口区域,理化因子变化剧烈,群落结构极不稳定,轮虫的多样性指数很好的响应了这种复杂环境,数值波动范围较大。3个站点之间,相对水流缓慢的滞水区S1所在水域,常有多艘船只停泊,污染程度最高,多样性指数最低,与非生物因子营养评价结果一致。
用轮虫的E/O和QB/T指数评价水体营养状况,在湖泊水质评价中应用较为广泛,通过对磨刀门河口进行水质生态学评价发现,E/O指数能准确的判定河口水体营养状态,且与理化指标判定一致;QB/T指数评价河口水体富营养程度较理化指标值偏低。QB/T指数通常适用于静水或者流速较慢的水体(Sládeček, 1983),但本研究中,QB/T值与理化评价差别并不大。
由于河口环境变化剧烈,轮虫与环境的关系复杂(Gutkowska等, 2013; Janakiraman等, 2012),仅从生态学评价磨刀门水质还不够全面,且有学者认为轮虫的污染等级也不能单纯以其种类和数量为准
(李共国等,2003),因而,评价一个水体的污染程度,应将生态学指标与理化指标结合起来综合评价,可获得更为真实的评价结果。
4 结论
1)调查期间共记录轮虫56种,其中污染指示轮虫42种,优势种为广布多肢轮虫、螺形龟甲轮虫、热带龟甲轮虫和暗小异尾轮虫。磨刀门水域轮虫多样性指数波动范围较大,轮虫群落结构不稳定。
2)轮虫丰度范围为0~199 ind·L-1,滞水区采样点丰度最高,丰度高峰出现在丰水期(8月下旬)。轮虫丰度与水温、总氮、叶绿素显著正相关(p<0.05),与盐度显著负相关(p<0.05)。
3)水质生态学评价指标显示磨刀门水体已达轻度富营养水平,与非生物环境因子营养状态综合评价结果一致。
ARNDT H. 1993. Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates) — a review [J]. Hydrobiologia, 255-256(1): 231-246.
DUGGAN I C, GREEN J D, SHIEL R J. 2001. Distribution of rotifers in North Island, New Zealand, and their potential use as bioindicators of lake trophic state [J]. Hydrobiologia, 446-447(1): 155-164.
GALKOVSKAJA G A. 1987. Planktonic rotifers and temperature [J]. Hydrobiologia, 147: 307-317.
GILBERT J J, BOGDAN K G. 1981. Selectivity of Polyarthra and Keratella for flagellate and flagellate cells [J]. Verhandlungen des Internationalen Verein Limnologie, 21: 1515-1521.
GREEN J. 1995. Associations of planktonic and periphytic rotifers in a Malaysian estuary and two nearby ponds [J]. Hydrobiologia, 313-314(1): 47-56.
GUTKOWSKA A, PATUREJ E, KOWALSKA E. 2013. Rotifer trophic state indices as ecosystem indicators in brackish coastal waters [J]. Oceanologia, 55 (4): 997-899.
HEINBOKEL J F, COATS D W, HENDERSON K W, et al. 1988. Reproduction rates and secondary production of three species of the Rotifer Genus Synchaeta in estuarine Potomac River [J]. Journal of Plankton Research, 10: 659-674.
HERBST D B. 2001. Gradients of salinity stress, environmental stability and water chemistry as a templet for defining habitat types and physiological strategies in inland salt waters [J]. Hydrobiologia, 466: 209-219.
JANAKIRAMAN A, NAVEED M S, ALTAFF K. 2012. Impact of domestic sewage pollution on rotifer abundance in Adyar estuary [J]. International Journal of Environmental Sciences, 3(1):689-696.
KOSTE W. 1979. Rotatoria:Die R dertiere Mitteleuropas [M]. Berlin: Gebr der Borntraeger.
LORENZEN C J. 1967. Determination of chlorophyll and pheo-pigments: spectrophotometric equations [J]. Limnology and Oceanography, 12(2): 343-346.
MEDEIROS A M A, BARBOSA J E L, MEDEIROS P R, et al. 2010. Salinity and freshwater discharge determine rotifer distribution at the Mossoró River Estuary (Semiarid Region of Brazil) [J]. Brazilian Journal of Biology, 70(3):551-557.
ONWUDINJO C C, EGBORGE A B M. 1994. Rotifers of Benin River, Nigeria [J]. Hydrobiologia, 272(1): 87-94.
PARK G S, MARSHALL H G. 2000a. The Trophic Contributions of Rotifers in Tidal Freshwater and Estuarine Habitats [J]. Estuarine, Coastal and Shelf Science, 51(6): 729-742.
PARK G S, MARSHALL H G. 2000b. Estuarine relationships between zooplankton community structure and trophic gradients [J]. Journal of Plankton Research, 22(1): 121-135.
ROUGIER C, POURRIOT R, LAM-HOAI T, et al. 2005. Ecological patterns of the rotifer communities in the Kaw River estuary (French Guiana) [J]. Estuarine, Coastal and Shelf Science, 63(1-2): 83-91.
SILVA A M A, BARBOSA J E L, MEDEIROS P R, et al. 2009. Zooplankton (Cladocera and Rotifera) variations along a horizontal salinity gradient and during two seasons (dry and rainy) in a tropical inverse estuary (Northeast Brazil) [J]. Pan-American Journal of Aquatic Sciences, 2(4): 226-238.
SLÁDEČEK V. 1983. Rotifers as indicators of water quality [J]. Hydrobiologia, 100(1): 169-201.
SOUSA W, ATTAYDE J L, ROCHA E S, et al. 2008. The response of zooplankton assemblages to variations in the water quality of four man-made lakes in semi-arid northeastern Brazil [J]. Journal of Plankton Research, 30: 699-708.
陈立斌,郭凯,赵文. 2010. 北京官厅水库轮虫群落结构与水体富营养化状况[J]. 湖泊科学, 22(2): 256-264.
陈立婧,顾静,彭自然,等. 2008. 滆湖轮虫群落结构与水质生态学评价[J]. 动物学杂志, 43(3): 7-16.
高原,赖子尼,王超,等. 2008. 珠江口浮游动物分布特征研究[J]. 中国水产科学, 15(2): 260-268.
胡德礼,刘秋海,吴超羽,等. 2009. 基于DEM的西江磨刀门水道近40年来河床演变特征研究[J]. 地理与地理信息科学, 25(2): 55-58, 62.黄祥飞,胡春英,伍焯田. 1985. 武汉东湖的轮虫[J]. 水生生物学报, 9(02): 129-142.
金相灿,刘鸿亮,屠清瑛. 1991. 湖泊富营养化调查规范[M]. 北京: 中国环境科学出版社: 286-302.
雷安平,施之新,魏印心. 2003. 武汉东湖浮游藻类物种多样性的研究[J].水生生物学报, 27(2): 179-184.
李共国,虞左明. 2003. 千岛湖轮虫群落结构及水质生态学评价[J]. 湖泊科学, 15(2): 169-176.
尚玉昌. 2002. 普通生态学[M]. 北京: 北京大学出版社: 284-295.
王家楫. 1961. 中国淡水轮虫志[M]. 北京: 科学出版社: 21-283.
王庆,杨宇峰. 2007. 珠江广州河段轮虫群落结构的初步研究[J]. 水生生物学报, 31(2): 233-239.
王云龙,袁骥,沈新强. 2005. 长江口及邻近水域春季浮游植物的生态特征[J]. 中国水产科学, 12(3): 300-306.
章宗涉, 黄祥飞. 1991. 淡水浮游生物研究方法[M]. 北京:科学出版社: 358-387.
Community structure of rotifers and ecological assessment of water quality in Modaomen of the Pearl River Estuary
WANG Qing1,2, HOU Lei1,2, CHEN Shi1,2, WU Zhihui3, YANG Yufeng1,2*
1. Institute of Hydrobiology, Jinan University, Guangzhou, 510632, China; 2. Key Laboratory of Eutrophication and Red Tide Control, Education Department of Guangdong Province, Guangzhou, 510632, China; 3. Beijing Normal University–Hong Kong Baptist University Untied International College, Zhuhai, 519000, China
Rotifers are a key part of the zooplankton community and react to changes in water quality. Rotifer abundance and species composition reflect the conditions and nutritional status of bodies of water, and, therefore, form a basis for the evaluation of water quality. Rotifer community structure was investigated at three different sites in the Pearl River Estuary: S1, a stagnant water district; S2, the center of the river; and S3, the confluence of rivers at the Modaomen outlet. Samples were obtained twice monthly from July 2009 to January 2010. Water temperature, salinity, transparency, chlorophyll a (chl a), total nitrogen (TN), total phosphorus (TP) and other physicochemical factors were measured simultaneously. The comprehensive trophic state index, TLIc, was evaluated using abiotic environmental factors. Also ecological assessment of water quality was carried out through not only rotifer species composition but also by the E/O, QB/Tand biodiversity indices. During the survey period, water temperature ranged from 12~29 ℃and salinity ranged from 0.11~7.9 PSU. The concentration of Chl a, which ranged from 0.89~9.56 μg·L-1, was positively correlated with water temperature (p<0.0001) and negatively correlated with salinity (p<0.01). The TLIc at the three sites ranged from 50-60, which suggested that water quality reached mild eutrophication. The degree of eutrophication descended in order from S1 to S2 and S3. During the survey period, 56 species of rotifer were recorded, of which 42 were pollution indicators. Rotifer species were highest at S1. The proportion of pollution indicator rotifer species of the total showed that the water in the Modaomen Estuary reached the eutrophication level, consistent with TLIc. Polyarthra vulgaris, Keratella cochlearis, K. tropica and Trichocerca pusilla were the dominant species. The rotifer abundances at S1, S2 and S3 ranged from 0~199 ind·L-1, 1~54 ind·L-1and 0~48 ind·L-1, respectively, with mean abundances of 31 ± 54 ind·L-1, 13 ± 15 ind·L-1and 11 ± 13 ind·L-1. The highest abundance was recorded at S1 in the late August; the abundances in the dry season were lower. Rotifer abundances were positively correlated with water temperature, TN, and Chl a.(p<0.05). A significant negative correlation was detected between salinity and rotifer abundance (p<0.05). The physicochemical factors in the Modaomen outlet of the estuary changed dramatically and the diversity index of rotifers fluctuated widely, which indicated that the community structure of rotifers was unstable. Redundancy analysis showed that water temperature, salinity and Chl a. were the most important factors influencing the seasonal variations of rotifer community structure. The ecological assessment of water quality indicated that the Modaomen outlet of the estuary had reached a level of mild eutrophication, in accordance with the comprehensive evaluation of trophic state based on abiotic environmental factors. The results show that a comprehensive evaluation combining ecological indicators and physical-chemical indicators is the most realistic way to assess water pollution.
Pearl River Estuary; rotifer; evaluation of water quality ; ecological assessment; eutrophication
Q784
A
1674-5906(2014)05-0824-10
国家自然科学基金项目(41173079);广东省自然科学基金(S2012040006824);高等学校博士学科点专项科研基金(20124401120013);广东高校优秀青年创新人才培养计划项目(2012LYM_0025)
王庆(1981年生),男,助理研究员,博士,主要从事河口浮游动物生态学研究。E-mail: wq2010@jnu.edu.cn
*通讯作者:杨宇峰,男,教授,博士生导师。E-mail: tyyf@jnu.edu.cn
2014-04-01
王庆,侯磊,陈实,吴志辉,杨宇峰. 珠江口磨刀门轮虫群落结构特征与水质生态学评价[J]. 生态环境学报, 2014, 23(5): 824-833.
WANG Qing, HOU Lei, CHEN Shi, WU Zhihui, YANG Yufeng. Community structure of rotifers and ecological assessment of water quality in Modaomen of the Pearl River Estuary [J]. Ecology and Environmental Sciences, 2014, 23(5): 824-833.
珠江是我国三大江河之一, 径流量居全国第二位。珠江流域位于典型的亚热带季风气候区, 高温多雨, 每年4-9月是珠江汛期(王庆等,2007)。珠江口地处亚热带,是咸淡水交汇水域,受珠江径流、广东沿岸流和外海水的影响,生态环境独特,生物组成复杂(高原等,2008)。磨刀门水道是珠江口八大口门之一,其入海径流量和输沙量在珠江八大口门中均居首位(胡德礼等,2009)。
轮虫是浮游动物群落中的重要组成部分,是河流生态系统中微食物网的重要环节(Arndt,1993)。轮虫个体小、繁殖快,对水质变化反应敏感。调查轮虫群落的种类组成和丰度变化,可以获知水质变化和水体的营养状况(Duggan等,2001;Janakiraman等, 2012),并可作为水质评价依据(Sládeček,1983; Gutkowska等, 2013)。国内外学者对轮虫在生物监测中的指示作用进行了较多的研究(Duggan等, 2001;Sládeček,1983;李共国等,2003;陈立婧等,2008;陈立斌等,2010),但对低盐河口轮虫群落特征和生物评价的研究较少。本文首次对珠江口磨刀门水域的轮虫群落结构进行每月2次的高频度调查,并从种类数、多样性指数、E/O指数、QB/T指数等几个方面分析了珠江口磨刀门轮虫群落结构的特点及其对环境因子的响应,旨在为我国轮虫区系和珠江口水质监测及评价提供科学依据。
猜你喜欢
诗歌月刊(2022年6期)2022-06-23
好孩子画报(2021年3期)2021-04-02
科学养鱼(2020年10期)2020-11-23
课外语文·中(2020年2期)2020-03-24
课外语文(2020年5期)2020-03-14
知识窗(2019年4期)2019-04-26
青年文学家(2016年35期)2017-04-26
广东海洋大学学报(2015年3期)2015-12-22
华南农业大学学报(2015年5期)2015-12-04
中国洗涤用品工业(2015年5期)2015-02-28
