
低温驯化对阿里山潜蝇茧蜂的影响
2014-04-04 03:19郭俊杰季清娥黄居昌陈家骅
环境昆虫学报 2014年1期
郭俊杰,季清娥,黄居昌,陈家骅
(福建农林大学益虫研究所,福建 福州 350002)
阿里山潜蝇茧蜂Fopius arisanus(Sonan)所有已知寄主均属实蝇科Tephritidae,且大多数属于果实蝇属 Bactrocera Macquart(Rousse et al.,2005),已有研究表明,阿里山潜蝇茧蜂是实蝇害虫寄生蜂中的优势种 (Haramoto et al.,1970;Vargas et al.,1993;Purcell et al.,1988;Wong et al.,1997;Harris et al.,1994)。福建农林大学益虫研究自引进阿里山潜蝇茧蜂以来,以桔小实蝇作寄主对其进行了人工大量繁育,并对其寄生效能、发育起点温度及有效积温、防控效果等进行了研究 (郭庆亮等,2006,2007;郑思宁等,2013)。但随寄主桔小实蝇在我国不断向北扩张 (汪恩国等,2013;屈海学等,2013),阿里山潜蝇茧蜂能否随寄主迁移,而在全国更大范围内定殖控制桔小实蝇尚有待研究。为此,本文对阿里山潜蝇茧蜂经低温驯化后以及越冬成蜂进行了过冷却点和结冰点的测定,并对越冬成蜂的寄生率和子代羽化率进行了研究,以期为该蜂在低温地区的定殖提供理论依据。
1 材料与方法
1.1 供试虫源
阿里山潜蝇茧蜂来源于室内以桔小实蝇为寄主人工饲养的种群,约第60代。饲养条件:幼虫在光周期为 L∶D=12∶12,相对湿度为75% ±5%,温度为25℃ ±1℃环境中饲养;成蜂在光周期为L∶D=12∶12,相对湿度为60% ±5%,温度为24℃ ±1℃的环境中饲养。
1.2 阿里山潜蝇茧蜂低温驯化后的过冷却点(SCP)和结冰点 (FP)
将羽化后24 h内的阿里山潜蝇茧蜂雌雄成蜂分别放入6℃的冰箱中驯化12 h、1 d、2 d、3 d,驯化完成后立即对其过冷却点和结冰点进行测定。每个处理雌雄虫各25头,以未经过低温驯化的成蜂作为对照。
过冷却点 (SCP)和结冰点 (FP)的测定:实验前先将温度计的热敏探头固定在一块厚薄均匀、平整的泡沫板上,然后将待测阿里山潜蝇茧蜂虫体对准热敏探头,用透明胶带将其固定在泡沫板上,在虫体上方盖一层脱脂棉,调整棉花厚度,使温度计的降温速率控制在1℃/min(李毅平等,1998)。将处理好的虫体连同泡沫板一起放进温度为-30℃的低温冰箱中。热敏电阻温度计显示屏上的温度开始下降,当降至虫体结冰温度时,虫体会迅速释放大量潜热,使虫体温度骤然上升,此时温度开始上升的转折点即为虫体的过冷却点,温度上升到一定程度后又突然开始下降,温度上升的最高点即为虫体结冰点。
1.3 阿里山潜蝇茧蜂越冬成蜂过冷却点 (SCP)及结冰点 (FP)
在头年12月将阿里山潜蝇茧蜂的蛹放在室外自然环境中,待其羽化后收集7日龄的雌雄成蜂,分别测定其的过冷却点和结冰点,测定方法同1.2。待阿里山潜蝇茧蜂在室外自然羽化12 d后提供寄主供其寄生,寄生过的寄主按寄生蜂饲养流程饲养,待幼虫化蛹后仍放置室外自然环境中羽化后,再次测定子代7日龄成蜂的过冷却点和结冰点。持续饲养至次年3月。
每个处理雌雄各25头,以室内人工饲养的7日龄成蜂作为对照。
1.4 阿里山潜蝇茧蜂越冬成蜂寄生率和子代羽化率的变化
分别给头年12月-次年3月在室外自然环境中羽化12 d后的阿里山潜蝇茧蜂提供寄主供其寄生。具体操作为在每个自制微型笼中引入20对寄生蜂,在每个接蜂盘上将500粒寄主卵均匀铺开,放置室温下寄生,24 h后取出接蜂盘,按寄生蜂饲养流程饲养。分别检测其寄生率、羽化率。每个处理设置5个重复,以室内人工饲养的阿里山潜蝇茧蜂作为对照。
1.5 数据处理
采用SPSS 17.0和Excel 2007软件处理。方法采用单因素方差分析和Duncan's新复极差法进行多重比较阿里山潜蝇茧蜂过冷却点和结冰点、成蜂寄生率及子代羽化率的差异。
2 结果与分析
2.1 低温驯化后阿里山潜蝇茧蜂成蜂的过冷却点及结冰点
阿里山潜蝇茧蜂雌雄成蜂SCP、FP均随驯化时间的延长呈先升高后下降的趋势;当处理时间为24 h时,雌雄成蜂SCP、FP均达到最高值,与对照差异显著;处理时间为72 h时,雌雄成蜂SCP、FP达最低值,但与对照无显著性差异。相同处理时间,雄虫SCP值高于雌虫。
雌雄蜂12 h与24 h组SCP、FP间无显著性差异,但与对照组间差异显著;48 h与72 h组SCP、FP间无显著性差异,且与对照组间也无显著性差异;但12 h、24 h组与48 h、72 h组SCP、FP间差异显著。SCP变化幅度最大是处理48 h,为5.1℃,最小的是处理24 h,为4.7℃;FP 24 h,为6.1℃,最小的是对照组,为4.7℃;见表1、表2。

表1 6℃低温驯化不同时间后阿里山潜蝇茧蜂成蜂过冷却点 (SCP)Table 1 The super cooling points of female and male adults of F.arisanus after exposed to 6℃for different hours

表2 6℃低温驯化不同时间后阿里山潜蝇茧蜂成蜂结冰点 (FP)Table 2 The freezing points of female and male adults of F.arisanus after exposed to 6℃for different hours
2.2 阿里山潜蝇茧蜂越冬成蜂过冷却点及结冰点
从头年的12月份到次年的3月份,随着时间的推移阿里山潜蝇茧蜂雌雄蜂的SCP、FP值均低于室内饲养的对照组,到次年3月份雌雄虫SCP、FP均降至最低值,与对照组差异显著,且同月份雌蜂的SCP、FP值均低于雄蜂。见表3、表4。
雌蜂头年12月份的SCP与对照组和次年1月份间无显著性差异,但1月份与对照组有显著性差异,其他各组间差异显著。次年3月份SCP变化幅度最大达5.4℃,变化幅度最小的是次年2月份,为4.5℃。
雄蜂的SCP12月份、次年1月份以及对照间无显著性差异,但均与次年2月份、3月份间有显著性差异,且次年2月份与3月份间也有显著性差异。SCP变化幅度最大的也是次年3月份为5.4℃,最小的是次年1月份为4.7℃。
从头年12月份-次年3月份,阿里山潜蝇茧蜂越冬雌雄成蜂的FP均是对照组、头年12月份、次年1月份三者间无显著性差异,而均与次年2月份、3月份间存在显著性差异,而后二者间差异显著。
2.3 阿里山潜蝇茧蜂越冬成蜂寄生率及子代羽化率的变化
阿里山潜蝇茧蜂越冬代成蜂寄生率及其子代的羽化率均随时间的推移先下降后上升,但均低于对照组;寄生率及其子代的羽化率均在次年2月份时降至最低,且与对照组相比差异显著,随后均回升,但仍低于对照组。
阿里山潜蝇茧蜂越冬代雌蜂次年2月份的寄生率与其他各组差异显著,而头年12月份、次年1月份及3月份间无显著性差异,但均与对照有显著性差异。
阿里山潜蝇茧蜂越冬代的子代羽化率头年12月份、次年3月份间无显著性差异,但二者与其他各组以及其他各组之间差异显著 (表5)。
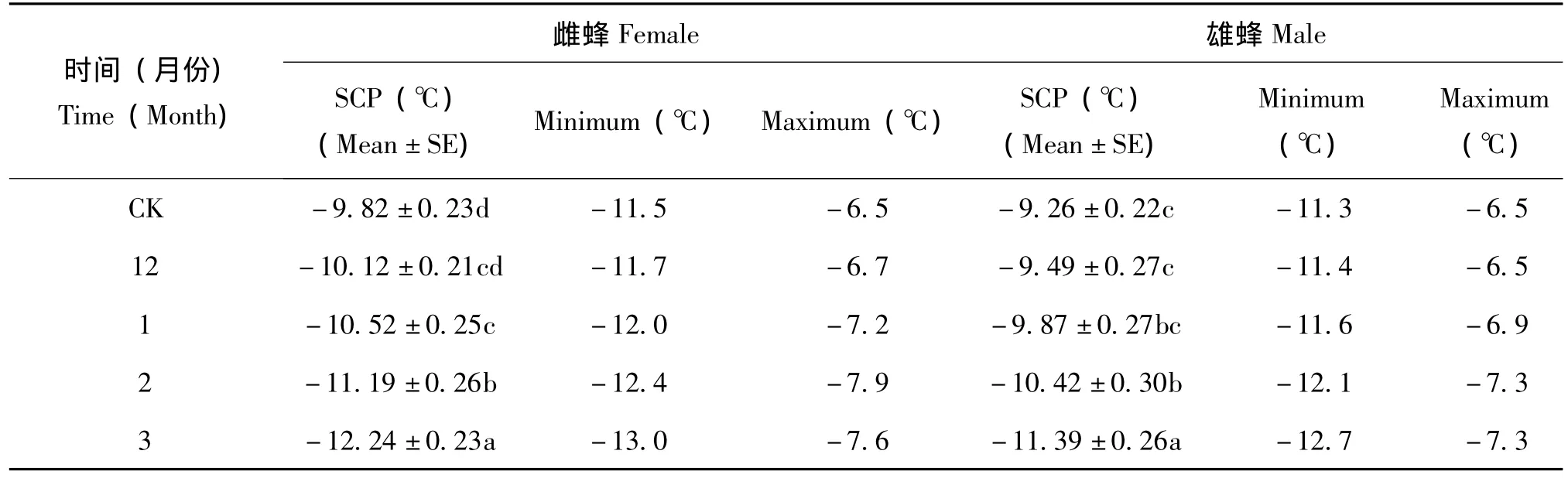
表3 阿里山潜蝇茧蜂越冬成蜂过冷却点 (SCP)Table 3 The super cooling points of overwintering adults of F.arisanus

表4 阿里山潜蝇茧蜂越冬成蜂结冰点 (FP)Table 4 The freezing points of overwintering adults of F.arisanus
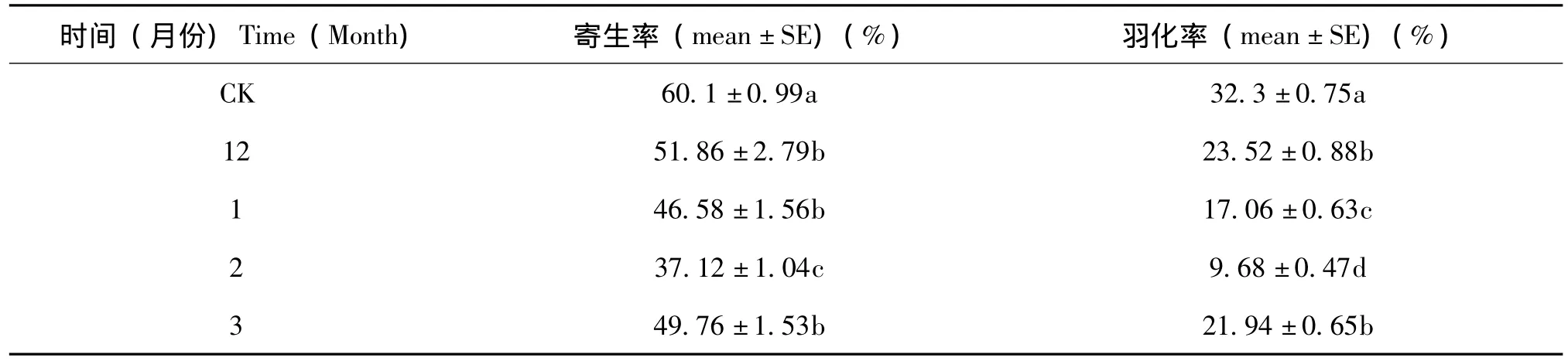
表5 阿里山潜蝇茧蜂越冬成蜂平均寄生率和子代平均羽化率Table 5 The parasitism rate of overwintering female adults and offspring emergence rate of F.arisanus
3 结论与讨论
经过低温驯化阿里山潜蝇茧蜂成蜂的过冷却点和结冰点均是先升高后下降,但随着驯化时间的延长,过冷却点和结冰点均与对照组相差异不显著,由此可推断,低温驯化并不能显著降低成蜂的过冷却点和结冰点,从而提高其耐寒性。
阿里山潜蝇茧蜂从头年12月份-次年3月连续三代在室外自然环境中,7日龄雌雄成蜂过冷却点和结冰点均是次年3月降至最低,可能原因是阿里山潜蝇茧蜂在室外越冬,长时间进行低温锻炼,连续几代后其子代过冷却点才会显著下降。已有研究标明,某个特定种或种群,只有经过一定量冷驯化后才足够刺激其抗寒机制其启动 (陈兵等,2005)。此外Lee et al.(1987),景晓红等(2002)同样认为,昆虫在经受低温胁迫前,通过一定时间的低温锻炼后,其耐寒性才可以显著提高。
阿里山潜蝇茧蜂越冬代成蜂的寄生率和其子代羽化率均随时间推移显著下降,在次年2月份时成蜂寄生率和子代羽化率均降至最低,与对照组差异显著,可能原因是2月份是福州最寒冷的天气,造成了阿里山潜蝇茧蜂成蜂性成熟时间延迟,同时低温导致成蜂行为不活跃,交配几率减少,但在3月份气温逐步回升,其寄生率和子代羽化率均有所提高。但低温环境对阿里山潜蝇茧蜂的影响需结合气象资料进行更加深入的研究。
Reference)
Rousse P,Harris EJ,Quilici S.Fopius arisanus,an egg - pupal parasitoid of Tephritidae,Overview [J].Biocontrol News and Information,2005,26(2):59-69.
Haramoto FH,Bess HA.Recent studies on the abundance of the Oriental and Mediterranean fruit flies and the status of their parasites [J].Proceedings of the Hawaiian Entomological Society,1970,20(3):551-556.
VargasRI, Stark JD, Uchida GK, etal. Opiine parasitoids(Hymenoptera:Braconidae)ofOrientalfruitfly(Diptera:Tephritidae)on KauaiIsland,Hawaii:Islandwide relative abundance and parasitism rates in wild and orchard guava habitats[J].Environmental Entomology,1993,22(1):246 -253.
Purcell M, HerrJC, MessingRH, etal. Interactionsbetween augmentatively released Diachasmimorpha longicaudata(Hymenoptera:Braconidae)and a complex of Opiine parasitoids in a commercialguava orchard [J]. BiocontrolScience and Technology,1988,8:139-151.
Wong TTY,Ramadan MM.Parasitization of the Mediterranean and Oriental fruit flies(Diptera:Tephritidae)in the Kula area of Maui,Hawaii[J].Journal of Economic Entomology,1987,80(1):77-80.
Harris EJ,Bautista RC.Fruit trap:A detection and collection tool for Opiine parasitoids(Hymenopera:Braconidae)of the oriental fruit fly,Bactrocera dorsalis(Diptera:Tephritidae)[J].Entomophaga,1994,39(3/4):341-349.
Guo QL,Huang JC,Ji QE,et al.Parasitical efficiency of Fopius arisanus on eggs of Bactrocera dorsalis[J].Entomological Journal of East China,2006,15(4):291-293.[郭庆亮,黄居昌,季清娥,等.阿里山潜蝇茧蜂对桔小实蝇卵的寄生效能[J].华东昆虫学报,2006,15(4):291-293]
Guo QL,Huang JC,Ji QE,et al.Studies on the developmental threshold temperature and effective accumulative temperature of Fopius arisanus[J].Entomological Journal of East China,2007,16(1):41-43.[郭庆亮,黄居昌,季清娥,等.阿里山潜蝇茧蜂Fopius arisanus的发育起点温度和有效积温[J].华东昆虫学报,2007,16(1):41-43]
Wang EG,Wang YC,Yu SH,et al.Population dynamics of the oriental fruit fly(Bactrocera dorsalis)in citrus orchard at Linhai,Zhejiang Province,China[J].Journal of Biosafety,2013,22(2):91-96.[汪恩国,王永才,余山红,等.临海柑桔园桔小实蝇种群数量消长规律[J].生物安全学报,2013,22(2):91-96]
Qu HX,Sun JS.Observation of the living habit of Bactrocera dorsalis[J].Chinese Horticulture Abstract,2013,2:51,62.[屈海学,孙静双.北京地区桔小实蝇的生活习性观[J].中国园艺文摘,2013,2:51,62]
Li YP,Gong H.Insect cryobiology:II.Ice nucleator agents(ice nucleator proteins)and insect antifreeze[J].Entomological Knowledge,2000,37(4):250-254.[李毅平,龚和.昆虫低温生物学:Ⅱ.冰核物质 (冰核蛋白)和昆虫的耐冻性[J].昆虫知识,2000,37(4):250-254]
Chen B,Kang L.The insect cold hardiness of geographic variation and seasonal fluctuation of the migratory locust and Liriomyza for example[M].Entomological Research Progress and Prospect.Beijing:Science Press,2005,170-202.[陈兵,康乐.昆虫耐寒性的地理变异和季节波动规律-以飞蝗和斑潜蝇为例[M].昆虫学研究进展与展望.北京:科学出版社,2005,170-202]
Lee REJr,Chen CP,Denlinger DL.A rapid cold-hardening process in insects[J].Science,1987,238:1415-1417.
Jing XH,Kang L.Research Progress in Insect Cold Hardines[J].Acta Ecologiac Sinica,2002,22(12):2202-2207.[景晓红,康乐.昆虫耐寒性研究.生态学报,2002,22(12):2202-2207]
猜你喜欢
作文周刊·小学一年级版(2022年24期)2022-06-18
海峡姐妹(2019年4期)2019-06-18
扬子江(2019年3期)2019-05-24
小天使·一年级语数英综合(2019年2期)2019-01-10
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
小学生导刊(低年级)(2017年1期)2017-06-12
广西蚕业(2015年3期)2015-11-30
语文知识(2015年9期)2015-02-28
广东第二课堂·初中(2009年8期)2009-08-26
